
The Effect of Parental Education and Parental Involvement on ...

PIETRO BRANDMAYR
RICERCHE ETOLOGICHE E MORFOFUNZIONALI SULLE CURE
PARENTALI IN CARABIDI PTEROSTICHINI (COLEOPTERA:
CARABIDAE, PTEROSTICHINAE).
Rerum natura. nusquam magis quam in minimis tota.
PLINIO
Estratto da REDIA, voI. LX, 1977, pp. 275-316, figg. 17, tabb. 2 Pubblicato dall' Istituto Sperimentale per la Zoologia Agraria di Firenze - 6 Ottobre 1977


PIETRO BRANDMAYR (*)
RICERCHE ETOLOGICHE E MORFOFUNZIONALI SULLE CURE
PARENTALI IN CARABIDI PTEROSTICHINI (COLEOPTERA:
CARABIDAE, PTEROSTICHINAE).
Dopo la pubblicazione di un primo tentativo di· sintesi dei comportamenti parentali noti nei Carabi di, che riguardava soprattutto la scoperta dei nidi pedotrofici di Carterus calydonius, (BRANDMAYR, 1974b; BRANDMAYR e BRANDMAYR-ZETTO, 1974), abbiamo concentrato le indagini sulla vasta tribù dei Pterostichini ('). All'interno di questo gruppo sono infatti note ben tre diverse modalità di ovodeposizione, e precisamente:
1) Interramento diretto delle uova nel substrato, in una piccola cavità scavata mediante l'apparato ovopositore, in particolare con le appendici biarticolate del IX urite (stili). È una modalità primitiva, certamente la più diffusa, e non implica ulteriore contatto tra madre e prole. È nota dei generi più vari, Abax, Pterostichus, Poecilus, Bothriopterus, Steropus, Calathus, Platynus, ecc. (risultati di allevamento di AA. vari).
2) Costruzione mediante l'apparato ovopositore di «capsule ovigere» di terriccio, che vengono poi attaccate al substrato o ad un annesso del suolo (pietra, muschio, ramoscelli). È probabile che in tal modo le uova siano meglio protette da disseccamento e soprattutto dagli attacchi fungini (LasER, 1970, 1972). È un comportamento piuttosto complesso rispetto alla media dei Carabidi, ma che, come il precedente, non si prolunga oltre l'ovodeposizione. Scoperto da BARGAGLI (1874) su Percus passerinii e), successivamente fu osservato da RILEY (in CLAASSEN, 1919 e KING, 1919) nei generi Chlaenius, Brachinus CS) e Galerita, dimostrando la vasta diffusione di tale
(*) Istituto di Zoologia ed Anatomia Comparata, Università di Trieste. C) Intesi qui come sinonimo degli «Pterostichidae» di }EANNEL (1941-42), autore sul
quale ci siamo basati in buona parte anche per l'ulteriore suddivisione (sottotribù e generi). [l) Dell'uovo l'autore scrive: «Un guscio di terra poco tenace lo rioopre completa
mente ». (') Recentemente ERVIN (1967) estende le osservazioni ad altre due spp. di Brachinus
(fidelis e pallidus).

- 276-
comportamento nella famiglia. Più recentemente la costruzione di capsule ovigere è stata seguita in laboratorio su Abax ater (LasER, op. cit.) ed A. springeri (BRANDMAYR e BRANDMAYR-ZETTO, op. cit.), da LUMARET (1971) su Percus (Pseudopercus) navaricus ed in natura da DICKER (1951) su Anchomenus dorsalis. Sembra dunque che questo tipo di cure prenatali, nelle quali l'ovopositore viene impiegato in maniera specializzata, sia molto diffuso nei Pterostichini e che l'estensione delle osservazioni a più generi e specie possa portare un contributo non trascurabile alla biologia del gruppo.
3) Ovodeposizione e veglia sulle uova in celle sotterranee, del tipo normalmente scavato per ibernazione o estivazione, sino alla schiusa ed alla pigmentazione delle larve. Si tratta di cure di tipo molto evoluto, oltrepassanti l'ovodeposizione e che pongono la madre a contatto con la prole, una vera« Brutpf1ege » secondo v. LENGERKEN (1954). Scoperte da BOLDORI (1933) sono oggi note per varie specie delle subtribus Pterostichina e Molopina. Alla lista pubblicata in BRANDMAYR e BRANDMAYR-ZETTO, comprendente 1 sp. di Abax, 4 di Molops, 2 di Tanythrix, 4 di Pterostichus (s.l.) sono da aggiungere Platysma am/;racinum (LINDROTH, 1946) ed Abax ovalis (LAMPE, 1975). In contrapposizione a quelle postnatali di Carterus calydonius (tipo II), queste cure sono state chiamate di tipo I, considerate meno complesse e ritenute una risposta evolutiva di Carabidi carnivori ad un raffreddamento del clima, forse alle glaciazioni pleistoceniche. Come nella modalità precedente, la veglia delle madri sulle uova avrebbe lo scopo di proteggere queste dagli attacchi fungini, frequenti soprattutto nelle spesse coltri umose dei suoli forestali. LAMPE (op. cit.) infatti ha osservato una maggiore mortalità in uova di A. ovalis sottratte alle celle materne. LaSER (1972) pone in relazione le cure parentali con le preferenze ambientali di singole specie: Abax ater, ad esempio, quale costruttore di capsule ovigere, non presenterebbe esigenze edafiche particolari (a patto che il terreno presenti una certa plasticità), mentre A. paralleus, esercitando cure parentali postnatali, sarebbe legato a suoli freschi, porosi e ben aerati, dotati di un certo grado di umidità.
Nel corso di questo lavoro sono state allevate od osservate in natura, sino all'ovodeposizione, 1 sp. di Molops, 5 di Abax, 1 di Pterostichus, 1 di Platysma. Alle osservazioni sul comportamento abbiamo voluto affiancare uno studio mONofunzionale dell'apparato ovopositore, poichè le tre diverse modalità di ovodeposizione descritte implicano un uso diverso delle strutture genitali da parte delle !j1 !j1. Si è tentato in genere di rispondere ai seguenti interrogativi:
Esiste un nesso tra cure parentali, esigenze ecologiche e fenologia delle singole specie?

- 277-
Quali sono le relazioni tra etologia e filogenesi nei Pterostichini? Le cure parentali postnatali sono da considerarsi una conquista re
cente del gruppo? Sono collegate a determinate caratteristiche morfofunzionali delle spe
cie che le presentano? In altre parole, è possibile riconoscere a priori, senza allevamento, ma basandosi sul solo esame morfologico, il tipo di cure esercitato dalle ~ ~ di una specie?
I risultati sono esposti e discussi separatamente per ogni genere.
MATERIALE E METODI
Trattandosi per lo più di specie montane o alpine, per gli allevamenti si è resa necessaria la stabulazione in armadi o celle climatiche, a temperatura regolabile e fotoperiodo controllato. Se non altrimenti indicato, le condizioni standard di allevamento corrispondono a 15 + 1 °e ed al giorno lungo delle latitudini dell'Italia settentrionale (lat. di rif. = 45°), con LD 15/9, intensità luminosa variabile tra 200 e 300 lux, da tubi fluorescenti Philips « dayligt » tonalità 55 ( = 6.500 °K). Si è curato che i substrati dei terrari corrispondessero il più possibile a quelli dei biotopi e delle località di provenienza. Gli esemplari venivano nutriti due volte per settimana con carne fresca, a pezzi o macinata. Le specie amanti sostanze zuccherine (gen. Molops e Pterostichus s.l.) hanno ricevuto anche frutta a pezzetti, soprattutto mela. Mediante annaffiamento si è mantenuta costante l'umidità del terreno.
Per lo studio mONologico abbiamo utilizzato preparati di ovopositori trattati con KOH (soluzione in acqua deionizzata). Lo studio biometrico su Abax è stato svolto con microscopio stereoscopico Galileo munito di oculare micrometrico. Per altri particolari vedansi risultati.
RISULTATI
Gen. Abax
Abax carinatus ssp. sulcatus Fiori.
L'allevamento è stato realizzato con Il esemplari raccolti in un bosco ripario a salici e pioppi nei pressi di Peteano (Gorizia), lungo il basso corso del fiume Isonzo, il 14/IV /1975.
Nove di essi sono stati mantenuti in condizioni naturali, all'aperto, men- .

- 278-
tre una sola coppia è stata posta a 15°C ed LD 15/9 (luce artificiale). Questi ultimi due esemplari si sono dimostrati molto più attivi ed è stato nel loro terrario che abbiamo rinvenuto, il lO/IV, 5 larve al I stadio, vaganti nel terriccio. Dopo 15 giorni venivano raccolte altre 6 larve, probabilmente nate dalla medesima ovodeposizione. Non si sono osservati comportamenti parentali di alcun tipo, nè capsule ovigere, nè tantomeno la veglia sulle uova in celle sotterranee, ed è quindi presumibile che la deposizione delle uova avvenga secondo la modalità più primitiva, il semplice interramento.
Interessante il fatto che solo individui esposti a temperature fresche siano in grado di riprodursi. È probabile infatti che gli esemplari del terrario all'aperto, con temperature spesso ben superiori ai 20"C, siano stati inibiti nella funzionalità delle gonadi. In questo secondo terrario le copule sono cessate ben presto, a meno di un mese dalla data di cattura. Le pochissime uova deposte erano abbandonate sul terreno, senza protezione, e non sembravano essere state fecondate (mancanza di sviluppo dell'embrione).
Abax exaratus Dejean; A. banningeri Scha'uberger; A. oblongus Dejean.
Queste specie filogeneticamente affini tra loro vengono trattate insieme: tutte e tre infatti depongono le uova in capsule ovigere, modellate con gli apici addominali.
A. exaratus è stato allevato con 8 ~ ~ ed 8 ~ ~ provenienti dalla Val Sessera, Biellese (Prov. di Vercelli, Piemonte) e dai dintorni del Santuario di Oropa, lungo il sentiero che porta al M. Mucrone (m 800-1900 s.l.m.), raccolti dal 17 al 19/VI/'75.
La superficie del terrario era ricoperta per metà da humus dolce, per metà da terriccio argilloso fme derivante dalla degradazione di rocce cristalline caratteristiche della prima località di raccolta (sieniti). Le ovodeposizioni hanno avuto luogo dal terzo giorno di stabulazione in poi, sino al mese di agosto, le copule si sono verificate sino ad anche un anno dopo. Il periodo di sviluppo embrionale è di 27-28 giorni a 15°C.
A. oblongus: 12 ~ ~ e Il ~ ~ provenienti da vari biotopi montani, sub-alpini ed alpini della Grigna settentrionale, dal p.sso del Cainallo sino al Rif. Bietti (m 1719) lungo la Cresta di Piancaformia, e sul versante meridionale della Cima Palone a N del Rif. Bogani (ex Monza, m 1816). Dopo poco più di una settimana dalla cattura, avvenuta dal 21 al 23/VI/'75 si osservano le prime capsule ovigere. Anche in questo caso le copule perdurano molto più a lungo delle ovodeposizioni. Sviluppo embrionale di 25 giorni a 15°C.
A. biinningeri: 13 ~ ~ ed 8 ~ ~ provenienti dalla medesima località della sp. precedente, raccolti soprattutto nelle faggete montane vicino al p.sso

- 279-
del Cainallo (m 1290), nei pressi di Esino Lario. Numerose le copule osservate, a partire dal giorno di cattura (20jVlj'75) sino almeno ,a novembre. Scarse invece le ovodeposizioni, avvenute dal mese di agosto in poi, cioè dopo due mesi di stabulazione alle condizioni standard. Una capsula ovigera è stata rinvenuta sul pezzo di carne dato in pasto agli adulti, ma si è trattato di un singolo caso.
L'insieme dei comportamenti osservati, integrati con i dati di LosER (1970, 1972) su A. ater, è suddivisibile in tre fasi principali:
a) La!j? in procinto di deporre raggiunge la parte più umida del terrario, estrae gli uriti addominali invaginati e con moto circolare degli stili inizia a raccogliere particelle molto fini di terriccio. Questo si ammassa per adesione in quantità crescente attorno all'apparato ovopositore, sino a che tutta l'estremità dell'addome si trova ricoperta da un sottile strato di argilla molle, giungente sin sotto alle elitre.
b) segue una fase di deambulazione della madre con tutta la masserella terrosa attaccata all'addome. :È la più lunga di tutta la sequenza comportamentale e può durare anche 15 o 20 mihuti. (4)
c) La !j? si avvicina al substrato prescelto (parte inferiore o superiore di una pietra, di una foglia secca, una zolla di muschio, un qualsiasi « annesso del suolo ») e vi appoggia l'addome, esercitando una certa pressione. L'uovo viene deposto in una cavità cilindrica trasversale modellata nella crosta terrosa dagli stili, dal lobo posteriore dei valviferi e dall'area membranosa del 9° sternite. Nel frattempo si notano numerose inarcature, verso l'esterno, dei lembi liberi della capsula, ed è probabilmente il bordo delle elitre ad incidere un solco nella massa argillosa, coincidente esattamente con il bordo superiore della cavità dell'uovo. Questa viene richiusa, a deposizione avvenuta, mediante il lembo superiore che si accolla a quello inferiore. Movimenti trasversali dell'apice addominale suggellano definitivamente la capsula, con la cementazione completa della parete rivolta alla madre. La durata di questa terza fase è breve, non superando i due minuti.
Non è escluso che la fase a) sia preceduta, almeno in certe specie, da una serie di piccoli morsi ad una zolletta di argilla, sino a renderla soffice e pastosa, in modo da essere poi facilmente raccolta con l'estremità addominale. Simile comportamento è stato osservato da LUMARET (1971) su Percus navaricus. Abbiamo talora osservato !j? !j? di A. exaratus vagare (fase b) con la
(4) Succede talora, almeno in laboratorio, che la femmina vagante con l'addome ricoperto d'argilla sia soipresaà divorare un pezzo di carne. Tentativi di copula da parte dei 8 8 in tale fase hanno sempre conseguenze negative suIl'ovodeposizione, causando la dispersione dell'argilla raccolta dalla madre. . .

- 280-
masserella terrosa sul pigidio e le mandibole sporche di argilla. In A. oblongus si sono notate spesso capsule incomplete abbandonate sul terreno, rivolte con la concavità verso l'alto, fatto pure notato da LUMARET (op. cit.). Si tratta a mio avviso di involucri di terriccio abbandonati all'inizio della fase b), forse perchè mal riusciti o inglobanti corpi estranei troppo voluminosi.
Non a caso quello di fig. I in LUMARET e quello qui raffigurato (fig. I D) inglobano entrambi un rametto o un pezzo di radice.
La forma delle capsule non varia specificamente, a differenza delle dimensioni che sono proporzionali a quelle degli apici addominali. Per A. oblongus ed exaratus il diametro ma&simo non è superiore ai 4,5 -5,0 mm, quello minimo è di 3 mm circa. Le capsule di A. biinningeri sono un po' più piccole.
La struttura degli involucri è stata esaminata con particolare attenzione in A. exaratus, che aveva a disposizione terriccio molto fine (vedi note specifiche, pago 278) e che conservava le impronte degli organi plasmatori. La fig. II rappresenta in a e b la capsula dopo la schiusa della larva, con la superficie di rottura ed il coperchio. Questo presenta al suo interno l'impronta dei segmenti (terghi) addominali della madre. Una linea di rottura principale sembra rappresentata dal « solco delle elitre» (linea nera ingrossata sopra la cavità dell'uovo), il quale ha probabilmente la funzione di facilitare la rottura del coperchio da parte della larvetta neonata. In c è invece raffigurata una sezione trasversale della celletta ovigera, con la cavità modellata da stili e val-viferi. La parete a diretto contatto con il sasso è sempre la più sottile: le <i1 <i1
infatti ottengono la perfetta aderenza dell'argilla al substrato dirigendo lo sforzo dell'apparato ovopositore su quella parete. È soprattutto a spese di questa che viene costruita la cavità contenente l'uovo, e non è raro il caso che essa sia delimitata, in quel punto, dal sasso stesso.
Significativi sono ancora, a nostro avviso, i seguenti fatti: A. exaratus, le cui <i1 <i1 avevano la scelta tra due diversi tipi di terreno,
ha costruito gli involucri ovigeri solamente con il terriccio argilloso proveniente dalla località di raccolta, ma non con l'humus, materiale meno plastico e poco adatto a simile comportamento parentale. Ciò starebbe ad indicare una scelta attiva tra tipi di terreno più o meno plastici.
Specie silvi cole come A. exaratus e biinningeri fissano le uova incapsulate tanto sulla superficie che sul lato inferiore delle pietre. Infatti nei boschi esse sono comunque protette dalla radiazione solare diretta, che viene intercettata dallo strato delle chiome, e quindi da un disseccamento troppo rapido ed intenso. Specie esclusivamente praticole come A. oblongus, affine sì ad exaratus, ma legato a formazioni vegetali erbacee di altitudine, depongono le uova soprattutto sul lato inferiore delle pietre. Nell'ambito dunque del medesimo

Fig. I
A, B. Capsule ovigere di A. exaratus dopo la fuoriuscita della larva. In A si nota anche il «coperchio ». Visibile la cavità trasversale dell'uovo con i residui del corion ed il solco formato sopra questa dall'apice delle elitre. Ingrandimento: x 6,5 risp. x 5. - C. Gruppo di capsule ovigere attaccate sul lato inferiore di una pietra da A. oblongus; x 3. -D. Involucro di terl1iccio a forma di coppetta, probabilmente .abbandonaw all'inizio della fase b, perchè inglobante un rametto; x 6,5.

- 282-
grado di complessità del comportamento materno, notiamo delle varianti di indubbio valore adattativo. La tendenza di A. oblongus ad ovodeporre solamente sulla superficie inferiore delle pietre rappresenta un'ulteriore garanzia di sopravvivenza delle uova in biotopi prativi sassosi alto-montani ed alpini, soggetti a forti escursioni termiche ed igriche.
Quanto alle funzioni della capsula argillosa, l'ipotesi di una difesa dagli attacchi fungini sembra forse la più plObante. Non ci sembra infatti possibile che un piccolo grumo terroso protegga l'uovo dal disseccamento in maniera più completa di una cella scavata direttamente nel suolo (a meno che l'uovo stesso non possieda esigenze particolari quanto a respirazione e traspira-
a b
Fig. II
Capsula ovigera dopo la fuoriuscita della larva (schematico). La supetficie di rottura è punteggiata ed è evidenziato il solco delle elitre. - b. Il coperchio, recante l'impronta dei somiti addominali. c -sezione longitudinale. La freccia indica il solco delle elitre.
zione). La capsula terrosa invece, soprattutto se impastata precedentemente con succhi gastrici o spruzzata eventualmente con il secreto di ghiandole pigidiali potrebbe essere una buona barriera contro la penetrazione delle spore e delle ife fungine.
La spiegazione del sorgere di queste cure nei Pterostichini o tribù affini piuttosto che, ad esempio, nei Carabini può essere ricercata anche a livello di preadattamento morfofunzionale. L'ovopositore di Abax può essere infatti definito anche un complesso di organi « plasmatori », specialmente se confrontato con quello più robusto, ma puramente « scavatore » di Carabus.
In Abax (fig. III) ii valvifero C) possiede un lobo posteriore, laterale
(5) La terminologia adottata è quella di LINDROTH (1957), modificata da NOONAN (1973) sulla base di note e disegni inediti di SNODGRASS su Pterostichus sp., conservati alla Smithsonian Institution, Washington, e con aggiunte di DANE LUTTO, 1975/76.

- 283-
pl.sg. VIII
2
VI V
Fig. III
tg.IX
vulva
-A'~-- art. disto si.
art. bs.sl.
emist.vlII
L'apparato ovopositore di Abax. 1, Visione ventrale. In questa figura come in molte seguenti le parti sclerificate esterne sono contrassegnate da punteggiatura lassa, quelle che nel preparato sono coperte da membrane articolari da punteggiatura fitta. Gli sterni addominali visibili sono rappresentati dal solo contorno. Scala in mm. -2, Ovopositore di Abax, visione laterale. Apd. = apodema; bs. = basale; a. = area; art. = articolo; artico = articolare; disto = distale; emist. = emisternite; est. esterno; ins. = inserzione; inv. = invaginato; lb. = lobo; mb. = membranoso; p. = parte; pl. = pleurite; posto = posteriore; scI. = sclerite o sclerificazione; sg. = segmento; sI. = stilo; st. = sterno; tg. = tergo; vf. = valvifel'O.
rispetto alla base dello stilo, che permette alla femmina di plasmare la cavità trasversale della capsula ovigera. Ciò può avvenire soprattutto se durante la preparazione di tale cavità cilindrica orizzontale e perpendicolare all'asse del corpo gli stili vengono accollati ai lobi posteriori, formando così con l'area membranosa rigonfia del go sternite una superficie plasmatrice emicilindrica. In Carabus invece (fig. IV) il valvifero non si espande poste-

- 284-
riormente a formare un lobo posteriore; manca inoltre l'area membranosa sternale, il che conferisce all'ovopositore dei Carabini un carattere più strettamente fossorio ed un'attitudine a forare substrati anche molto resistenti. Anche la presenza dei due scleriti vulvari (residuo probabile di una sclerificazione sternale), corrispondenti ai« Gelenkfortsatze » di FRANZ (1929) è in accordo con la funzione scavatrice dell'apparato, proteggendo l'apertura vaginale e impedendo la penetrazione di corpi estranei durante la ovodeposizione.
ins.mb.artic. VIII-IX
2 Pig. IV
1, 2. Ovopositore di Carahus creu/zeri P., visione ventrale risp. dorsale: VI: = vulvare; end. = condilo; proc. = processo. Per le altre abbreviazioni vedasi fig. III.
Abax ovalis Duftschmid.
IX
Allevato allo scopo di verificare sulle popolazioni delle Alpi orientali il comportamento parentale osservato da LAMPE, (1975) per quelle dello Schiefergebirge (dintorni di Koln).
I due & & e le tre ~ ~ provenienti dalla Selva di Tarnova (Trnovski Gozd, Smrekova Draga, m 100 circa s.l.m.), altopiano carsico boscoso situato ad E di Nova Gorica (Slovenia occ.), si sono dimostrati molto attivi e sessualmente maturi sin dai primi giorni di stabulazione (22/V /,75). Anche le popolazioni della località citata vegliano sulle uova in nidi sotterranei. Lo sviluppo embrionale a 15°C dura circa un mese. Come di norma nessuna delle

- 285-
due femmine-madri ha mostrato, anche durante l'esecuzione di fotografie, reazioni di fuga, nè si è potuta osservare opera di pulitura delle uova, anzi queste sono spesso pressate contro il fondo della cella e semicoperte di limo (6). Il risultato positivo è un argomento a favore della costanza intraspecifica dei fenomeni etologici osservati, del fatto cioè che le modalità di ovodeposizione non variano a seconda della popolazione o della regione geografica presa in considerazione.
Fig. V
Abax beckenhaupti (esemplare ~), specie di un gruppo praticolo alto-montano ed alpino, caratterizzato dalla mancanza del rilievo careniforme alla base della VII interstria elitrale; x 2,2.
Nel corso di questa ricerca si è tentato di allevare anche altre specie di Abax, e precisamente beckenhaupti Duftschmid, eccheli Bertolini, continuus Baudi. I risultati negativi sono dovuti in parte all'inadeguatezza del substrato, in parte allo stato di senescenza degli individui raccolti.
Le ovodeposizioni condotte a buon fine e riguardanti 5 specie, unite ai dati già noti dalla letteratura, permettono di fornire un quadro sintetico abbastanza soddisfacente sul comportamento parentale nel genere Abax (Tab. I).
Oggi sono dunque conosciute le cure di 9 delle 17 specie di Abax note
(6) L'inibizione del meccanismo scatenante predisposto alla reazione di fuga è cosÌ forte, che è possibile, mediante una pinzetta, togliere i piccoli grumi di terra dalle elitre della !i? senza che questa dia segni manifesti di inquietudine.

- 286-
Fig. VI
Cella-nido di A. ovalis, con larve appena schiuse o già pigmentate. Durante i controlli spesso la !j? -madre s'addossa obliquamente ad una parete laterale dell'escavazione; x 3.
TABELLA I
Interramento Costruzione
Veglia sulle Specie diretto d. uova
di capsule uova in celle Autore
con gli stili ovigere con
sotterranee gli stili
ovalis + LAMPE (1975) e hoc opus
parallelus + LaSER (1970, 1972) ater + LaSER (1970, 1972) exaratus + hoc opus biinningeri + hoc opus oblongus + hoc opus springeri + BRANDMAYR e BRAND-
MAYR-ZETTO (1974) carinatus + hoc opus pyraeneus + XAMBEU (1900)

- 287-
(secondo BRANDMAYR, 1976, in c. di stampa; DANELuTTo, 1975/76). La distribuzione dei tre diversi tipi di comportamento non sembra a prima vista legata a particolarità morfologiche esterne della specie. È però interessante notare come le cure evolute, cioè la veglia sulla covata in celle sotterranee sino alla schiusa ed alla pigmentazione delle larve, sia comparsa esclusivaDJ.t!nte nel cosiddetto « secondo gruppo» di FIORI (1896) C) cioè negli Abax aventi l'articolo ungueale privo di spine, mentre la costruzione di capsule ovigcre sembra invece comparsa nel primo gruppo, ad articolo ungueale inferiormente spinoso.
A. pyraeneus però, pur appartenendo al secondo, sembra esercitare ,cure ancora più primitive, vale a dire il normale interramento delle uova.
Ci si è quindi posto il quesito: esistono caratteri morfologici in base ai quali sia possibile dedurre a priori la modalità di ovodeposizione delle diycrse specie?
Ci siamo naturalmente rivolti all'apparato che più direttamente è coinvolto in questi comportamenti, l'ovopositore, cercando di stabilire una c:orrelazione tra di esso e l'etologia delle ~ ~. Avendo notato in precedenza una certa differenza nelle dimensioni relative degli stili tra A. ovalis ed altre specie, abbiamo approfondito il problema da un punto di vista biometrico. Infatti le dimensioni degli stili corrispondono proporzionalmente alle dimensioni di determinati muscoli che entrano in azione durante il modellamento delle capsule ovigere o nell'interramento dell'uovo. Si può quindi presumere che ,specie utilizzanti l'ovopositore possiedano stili più voluminosi nei confronti del resto dell'addome, (8) rispetto a quelle che lo mantengono inattivo.
Abbiamo perciò misurato la lunghezza e la larghezza massima degli stili di tutte le spp. di Abax a nostra disposizione, confrontandole con lunghezza e la'rghezza massime del VII sterno addominale, moderatamente schiacciato sotto un coprioggetto (preparati in KOH). Immaginando che le inserzioni muscolari abbiano una superficie proporzionale a quella degli organi, abbiamo ricavato il rapporto: lunghezza X larghezza stili/lunghezza X larghezza VII urosterno. Tale rapporto dovrebbe dunque essere tanto maggiore quanto maggiori sono le dimensioni e quindi l'efficacia degli stili in rapporto alle dimensioni addominali.
(') In base a DANELUTTO, 1975/76, A. schuppeli e carinatus non rientrano in nessuno dei due gruppi di FIORI; vedi albero filogenetico, fig. 11.
(') Il volume dell'addome, struttura fortemente canalizzata, non var,ia sensibilmente in relazione al comportamento materno della specie. In quelle a cure parentali postnatali c'è comunque la tendenza all'arrotondamento del contorno posteriore del VII utosterno, fatto che è ben visibile soprattutto in Molops.
- 288-
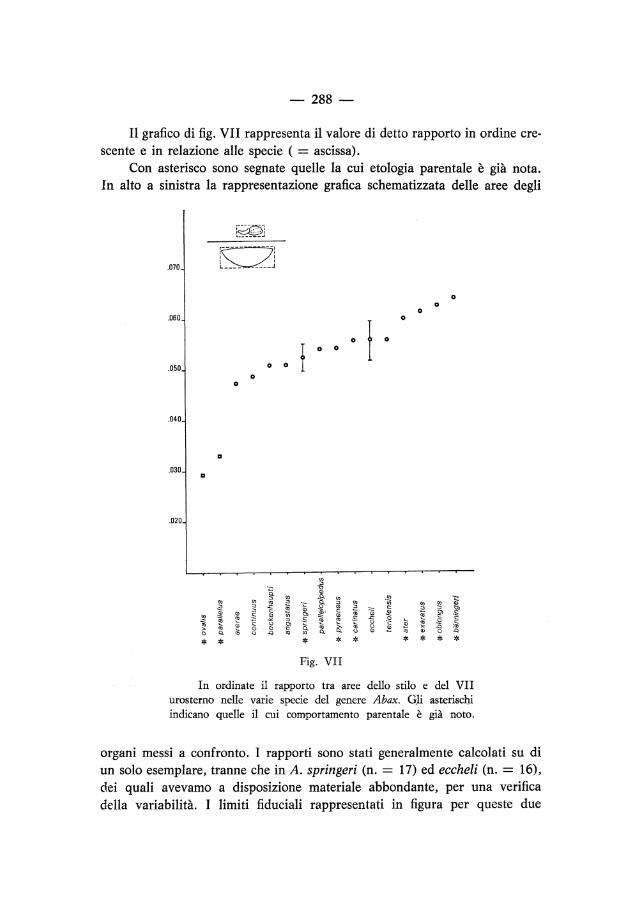
Il grafico di fig. VII rappresenta il valore di detto rapporto in ordine crescente e in relazione alle specie ( = ascissa).
Con asterisco sono segnate quelle la cui etologia parentale è già nota. In alto a sinistra la rappresentazione grafica schematizzata delle aree degli
.070
o o
o .060 o
.050 o
o
.040
" .030
"
.020
'" 15.
il "
'" " .g. .;: ~ ';: ~ '" '" " .2 "'" " " 13 " '" ~ " <: ". ;; '" .S
'" ~ '" ~ .~ "" <: <: ~ ~ ~
" .§ " àì ~ <:
~ ~ ~ <ll Q ~ " ;; .Q :~
o Q " .Q '" Q " " o .Q
* * * * * * * Fig. VII
In ordinate il rapporto tra aree dello stilo e del VII urosterno nelle varie specie del genere Abax. Gli asterischi indicano quelle il cui comportamento parentale è già noto.
organi messi a confronto. I rapporti sono stati generalmente calcolati su di un solo esemplare, tranne che in A. springeri (n. = 17) ed eccheli (n. = 16), dei quali avevamo a disposizione materiale abbondante, per una verifica della variabilità. I limiti fiduciali rappresentati in figura per queste due

- 289-
specie sono stati calcolati per un coefficiente di sicurezza del 99,9% e sono un buon indice della attendibilità degli altri rapporti, eseguiti con metodologia uniforme e dalla medesima persona.
Il grafico permette sostanzialmente di individuare 2 gruppi di Abax: quelli a stili ridotti e quelli a stili normali.
Il primo coincide con le cure parentali già note di A. ovalis e parallelus, i quali vegliano sulle uova in celle sotterranee e non usano più gli stili. Il secondo, molto più numeroso, assomma i costruttori di capsule ovigere e gli interratori di uova, quindi tutte le specie che normalmente usano l'ovopositore.
Possiamo quindi ragionevolmente concludere che A. ovalis e parallelus sono le due uniche specie del genere ad esercitare cure parentali evolute e che è possibile, mediante dati biometrici e considerazioni morfofunzionali, prevedere che anche le altre specie ad etologia parentale sconosciuta usano ancora l'ovopositore.
Le ricerche su questo apparato sono state estese ai generi sistematicamente più prossimi ad Abax: Percus e Molops.
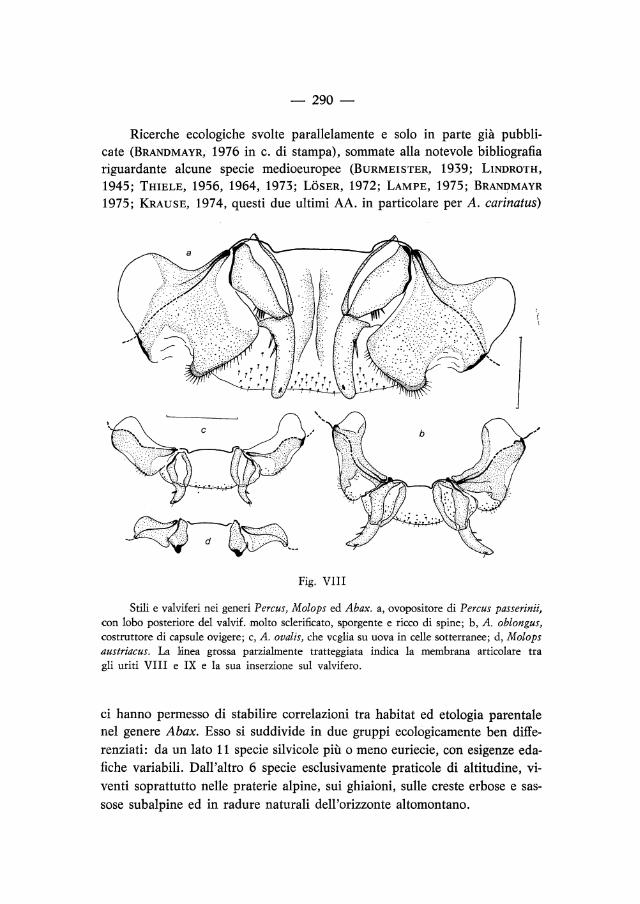
La fig. VIII rappresenta in visione ventrale stili e valviferi di alcuni rappresentanti dei tre generi. Molops è un genere che BRANDMAYR e BRANDMAYRZETTO (1974) hanno ipotizzato a «cure parentali primarie», vale a dire evolutosi da ceppi che già vegliavano sulle uova in celle sotterranee e che quindi da tempo ha smesso di usare l'ovopositore, vedi pago 297 e segg. Infatti in esso il valvifero possiede apodemi molto ridotti ed i due articoli dello stilo sono fusi (o comunque molto rimpiccioliti). In Abax le due uniche specie a cure uguali a quelle di Molops mostrano una chiara riduzione degli stili e delle espansioni apodematiche dei valviferi, ma il grado di regressione dell'intero apparato è molto minore. Specie con etologia non « molopoide » (es. nella figura: A. oblongus) hanno stili e valviferi normali e ben sclerificati. Percus, la cui attività di costruttore di capsule ovigere è nota (BARGAGLI, 1874; LUMARET, 1971) possiede valviferi e stili molto grandi e fortemente sclerificati.
Facciamo notare anche a questo proposito come nel genere Abax, il quale possiede quindi caratteristiche etologiche intermedie tra Percus e Molops, le cure parentali più evolute siano comparse in due specie del secondo gruppo, mancante delle spine sull'ultimo articolo tarsale.
Non è escluso che tale carattere sia correlato (in quale modo non sappiamo) ad una « predisposizione » verso un'evoluzione molopoide del comportamento.
- 290-
Ricerche ecologiche svolte parallelamente e solo in parte già pubblicate(BRANDMAYR, 1976 in c. di stampa), sommate alla notevole bibliografia rìguardante alcune specie medioeuropee (BURMEISTER, 1939; LINDROTH, 1945; THIELE, 1956, 1964, 1973; LasER, 1972; LAMPE, 1975; BRANDMAYR 1975; KRAUSE, 1974, questi due ultimi AA. in particolare per A. carinatus)
Fig. VIII
Stili e valviferi nei generi Percus, Molops ed Abax. a, ovopositore di Percus passerinii, con lobo posteriore del valvif. molto sclerificato, sporgente e ricco di spine; b, A. oblongus, costruttore di capsule ovigere; c, A. ovalis, che veglia su uova in celle sotterranee; d, Molops austriacus. La hlnea grossa parzialmente tratteggiata indica la membrana articolare tra gli uriti VIII e IX e la sua inserzione sul valvifero.
ci hanno permesso di stabilire correlazioni tra habitat ed etologia parentale nel genere Abax. Esso si suddivide in due gruppi ecologicamente ben differenziati: da un lato Il specie silvicole più o menoeuriecie, con esigenze edafiche variabili. Dall'altro 6 specie esclusivamente praticole di altitudine, viventi soprattutto nelle praterie alpine, sui ghiaioni, sulle creste erbose e sassose subalpine ed in radure naturali dell'orizzonte altomontano.

- 291-
Non si può parlare di legame stretto tra la formazione vegetale e cure parentali. Possiamo solo affermare che:
la veglia sulle uova in celle sotterranee ha luogo solo in due specie silvicole;
la costruzione di capsule ovigere di terriccio sembra essere compatibile tanto con l'ambiente praticolo che con quello silvicolo. Nel primo caso vi è una tendenza ad attaccare gli involucri sulla faccia inferiore delle pietre, nel secondo questi vengono fissati dove capita, su un qualsiasi annesso del suolo. Le capsule ovigere sarebbero quindi connesse non solo ad esigenze edafiche minori (LaSER, 1972), ma, più in generale, ad una maggiore possibilità di colonizzazione di ecosistemi ~Illche molto diversi tra loro (bosco/prato);
la deposizione diretta nel terreno ha luogo pure solo in specie silvicole;
l'ambiente delle formazioni boschive presenta quindi la maggior varietà di cure parentali.
Quest'ultimo fatto in particolare farebbe pensare ad un'origine silvicola del gen. Abax (soprattutto perchè tra le spp. silvicole troviamo anche la modalità più primitiva di ovodeposizione).
La conquista delle formazioni prative alpine sarebbe invece posteriore, molto probabilmente glaciale. Tutto ciò in accordo con il fatto che Abax è da considerarsi, nell'ambito dei Molopina, a differenziazione in gran parte recente (opinione sostenuta da HORION, in litt. a LasER, 1970; THIELE, 1971; come pure da MARCUZZI, 1952/53; 1962). Uno sguardo all'albero filogenetico del genere, ricostruito in DANELUTTO {1975/76), permette di evidenziare le relazioni tra evoluzione e cure parentali. Il ceppo originario, probabilmente tardo-pliocenico o di origine anche anteriore, avrebbe posseduto cure parentali semplici, cioè l'interramento delle uova, come certamente sappiamo di carinatus. La dicotomia più importante di tutta la storia del genere (segnata con asterisco), corrispondente alla sinapomorfia: articolo ungueale spinoso (plesiomorfo, ramo a sin.) - a.u. liscio (apomorfo, ramo di dest.) segna probabilmente anche una svolta etologica. Infatti gli Abax del primo gruppo sembrano caratterizzati in toto dalla costruzione di capsule ovigere di argilla, ed è tra essi che si sono segregati polifileticamente nel quaternario i due ceppi praticolialpini (gr. beckenhaupti egr. oblongus). Nell'altro ramo (<< secondo gruppo») sono invece comparse cure parentali postnatali, ma si tratta senza dubbio di un'acquisizione molto recente, com'è testimoniato non solo dalla scarsa regressione subita dall'apparato ovopositore, ma anche dal fatto che A. parallelus ed ovalis costituiscono un gruppo chiaramente fenetico

// schuppeli
- 292-
./-....-.-., parallelus \
Fig. IX
I i
",,---- ...... /" oblongus "'\
/ \ I \
/ \ I arerae , I ,
/ leriolensis l I I I eccheli J
I springeri /
/ " beckenhaupli //
....... _---""'/
Albero filetico del gen. Abax (compreso Abacopercus schiippeli Palld., il cui ovipositore è di dimensioni normali), com'è stato riscontrato in DANELUTTO (tesi di laurea ined., 1975/76) sulla base di dati morfofunzionali degli apparati genitali, ecologia, etologia e distribuzione geografica, e prendendo in considerazione anche caratteri morfologici larvali. La linea grossa continua racchiude le spp. del «primo gruppo », che probabilmente sono tutte plasmatrici di capsule ovigere; quella tratteggiata racchiude i praticoli di altitudine, ad insorgenza chiaramente polifiletica; il tratteggio punteggiato evidenzia l'insorgere distinto delle cure parentali evolute in parallelus ed ovalis. Sono sottolineati i nomi delle specie per le quali la modalità primitiva del semplice interramento delle uova è accertata.
dal punto di vista del comportamento, malgrado la fortissima affinità etologica. A. parallelus è in realtà ben più affine ad A. continuus, specie ad ovopositore non regredito, originata si da popolazioni padane di parallelus rimaste isolate nell'Italia settentrionale durante un periodo catatermico molto recente. Ciò è comprovato non solo dalla distribuzione geografica allopatrica e complementare delle due specie, ma anche dalla morfologia dell'apparato genitale, in particolare dell'endofallo (DANELUTTO, op. cit.).
Queste ultime considerazioni costituiscono una prova ulteriore in fa-

- 293-
vore di un'origine recente delle cure parentali postnatali dei Pterostichini, probabilmente sotto l'influsso del clima pluviale pleistocenico. Nel gen. Abax la loro origine è certamente secondaria, posteriore cioè alla formazione delle specie che le presentano, motivo per il quale il loro apparato ovopositore è ancora scarsamente regredito, specie se confrontato con quello del gen. Molops. La costruzione di capsule ovigere sembra caratterizzare un gruppo di Abax filogeneticamente omogeneo, complessivamente più euriecio, e sembra di origine più antica (tardo Pliocene?). Alcune specie infine (sicuramente pyraeneus e carinatus) hanno conservato nel tempo la modalità più primitiva di ovodeposizione.
Il genere studiato costituisce l'interessante esempio di un gruppo di insetti la cui notevole uniformità di shuttura scoraggiava a prima vista ogni interpretazione in senso adattativo del suo frazionamento in specie. In realtà la diversificazione dei ceppi si è avuta non solo a livello fisiologico-ecologico, come posto in evidenza dai lavori di LaSER (1972), THIELE (1973), LAMPE (1975), ma anche etologico e morfofunzionale.
Gen. Molops
Molops striolatus ssp. striolatus Fabricius.
Le osservazioni di KOMAREK (1954), JEANNEL (1948), sommate all'esperienza personale (BRANDMAYR e BRANDMAYR-ZETTO, 1974) ci hanno indotto a considerare questo genere come interamente caratterizzato da cure parentali evolute, protraentisi oltre l'ovodeposizione. L'allevamento di M. striolatus, riuscito dopo alcuni tentativi falliti, rappresenta un'ulteriore prova della validità dell'ipotesi. Visto il risultato negativo di esperienze precedenti, basate su un numero troppo alto di individui per terrario e su temperature di stabulazione tropo elevate (23°C), si è tentato con un capace cristallizzatore (13 = 25 cm) contenente solo 2 a a e 3 ~ ~ provenienti dagli abieti-faggeti della Selva di Tarnova (Trnovski Gozd, Slovenia, 22/V /,75). Riempito di humus dolce e dotato di alcune pietre per riparo, è stato mantenuto a temperature molto fresche (14-15 ~C), corrispondenti a quelle medie della coltre umosa forestale delle altitudini di provenienza (1000 m circa) nella tarda primavera (dati in. di DROVENIK, 1975). Avendo notato, dopo più di un mese di stabulazione e numerose copule, che ~ ~ tendenti ad isolarsi venivano molestate dagli altri esemplari, abbiamo aggiunto pezzi di coccio sino a coprire quasi interamente la superficie dell'humus. Dopo altri 10 giorni una ~ si rinchiude sotto una pietra, in una cella elissoidale (cm 3 X 5), molto appiat-

- 294-
tiia, deponendovi le prime due uova, seguite nel corso della settimana successiva da altre tre. Una seconda ~ inizia pure l'ovodeposizione, in un'altra cella, sul cui fondo nel corso di soli due giorni si ammucchiano ben 11 uova. Durante i controlli, eseguiti sollevando i ripari, le madri si dimostrano inquiete, spostandosi nel perimetro dell'escavazione, ma senza oltrepassarne i bordi, anche se mal definiti. Neanche in questa specie ho potuto osservare la pulizia delle uova mediante gli gnatiti, come KOMAREK (op. cit.) in M. piceus, nè la difesa attiva a colpi di mandibole di Tanythrix senilis (LEONARDI, 1969).
Fig. X
'i' di Molops striolatus durante la vq;lia sulle uova. Le antenne sono rigidamente protratte in avanti (segno di stress dovuto al controllo?).
In entrambe le ~ ~ si è osservata oofagia, ad una settimana dall'ultima ovodeposizione, segno probabile di condizioni di allevamento non ottimali. LAMPE (1975) ritiene che in A. ovalis l'oofagia possa essere determinata da un'umidità troppo bassa del substrato. Malgrado la notevolissima durata del periodo di sviluppo embrionale (2 uova, isolate a 14 ·C, si sono schiuse dopo quasi 50 giorni!) le madri di norma probabilmente non abbandonano la cella per nutrirsi. Femmine sessualmente mature, dissezionate nei mesi di giugno

- 295-
e luglio presentano di solito un corpo adiposo sviluppatissimo, a spese del quale si sostengono durante la vigilanza sulla « covata ».
Questo tipo di comportamento può essere posto in relazione con il ciclo fenologico, pure molto peculiare, di questo Molops. In base a rilievi eseguiti soprattutto sul M. Slavnik (M. Taiano) in Istria (Slovenia occ.), vetta carsica a 1000 m. s.l.m., tale ciclo si può considerare sostanzialmente bimodale, con una prima comparsa nel mese di aprile o di maggio, un massimo in maggio-giugno ed una successiva graduale scomparsa verso la fine di luglio.
Fig. Xl
Disturbata dal lampo fotografico la ~ -madre è retrocessa di 2 cm, senza però oltrepassare il bordo ideale della cella (che J1isulta meglio delimitata solo sulìa destra del fotogramma). Le antenne permangono rigidamente protese.
In settembre la specie ricompare nuovamente alla superficie del terreno, e la sua attività si protrae sino a stagione molto inoltrata (novembre). La fig. XII rappresenta il ciclo annuale di attività degli adulti, costruito su dati ottenuti con trappole di Barber, munite di esca zuccherina. Esso è correlato ai fattori temperatura, evaporazione e precipitazioni piovose, ma la scomparsa della popolazione durante il massimo calore estivo non è interpretabile solo in termini di estivazione. L'esercizio di cure parentali in celle sotterranee, a veglia

- 296-
di uova dal periodo embrionale molto lungo, chiarisce invece non solo l'interruzione della curva fenologica, ma anche l'andamento annuale del rapporto tra i sessi, con assoluta predominanza di ~ ~ nel mese di luglio, nel quale le ~ ~ sono già sotterra ad apprestare le celle di cova. L'intero ciclo biologico
di M. striolatus è in realtà di un tipo sinora riscontrato in ben poche specie di Carabidi (LAMPE, 1975). Lo sviluppo di una generazione completa richiede infatti 2 anni, ed è riassumibile nello schema di fig. XIII. Le larve alI stadio, nate verso la fine dell'estate, nel periodo autunno-primo inverno raggiungono lo st. II e poi il III, giungendo a impuparsi in primavera ed in estate.
La nuova generazione, riconoscibile alla dissezione per la scarsità di corpo adiposo, l'immaturità delle gonadi e la ridotta sclerificazione degli edeagi (gli esemplari escono in superficie a melanogenesi completa) segue la frazione sessualmente matura della popolazione nella sua estivazione. Ricompare poi brevemente in autunno (stagione nella quale il rapporto sessi si inverte a favore delle !j! !j!, perchè i maschi vecchi muoiono sottoterra), ma si riproduce solamente l'anno successivo, probabilmente grazie ad una dormanza geneticamente fissata, dopo aver completato le riserve adipose.
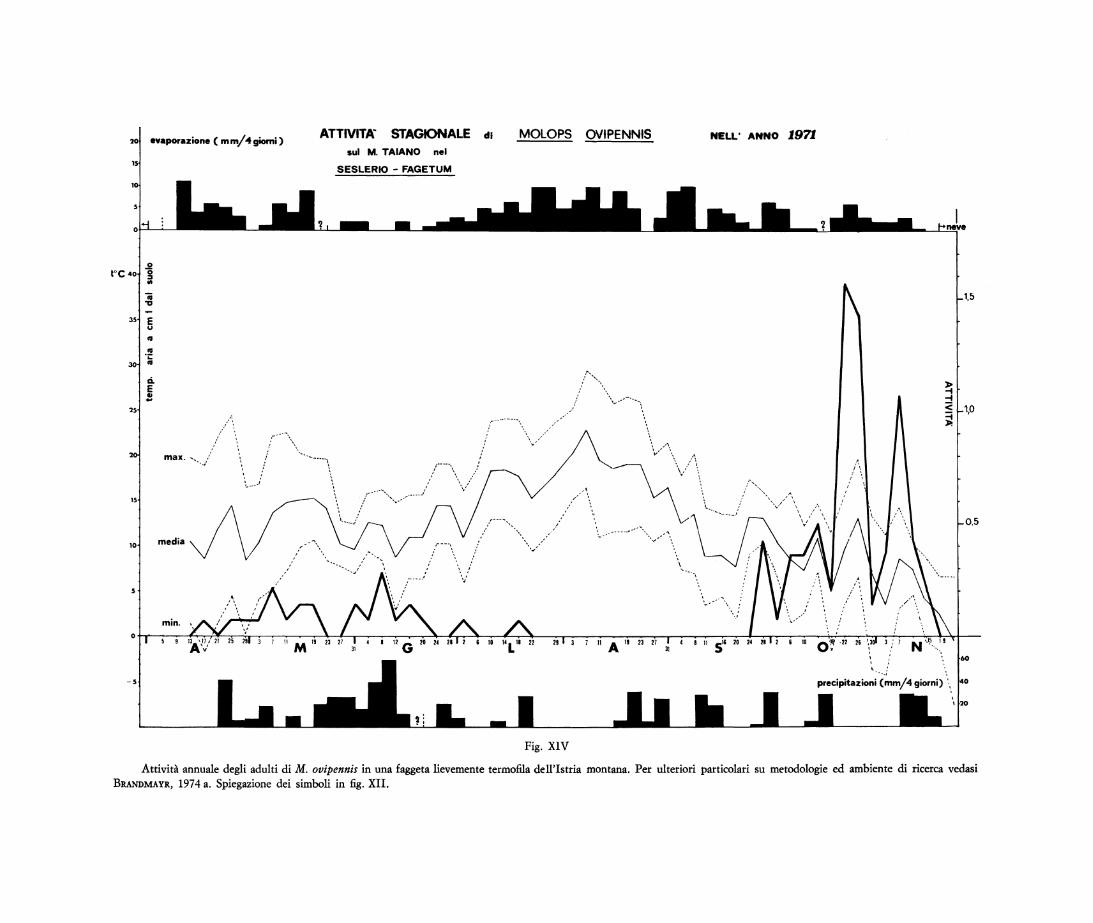
Molto simile il ciclo biologico di M. ovipennis (9), le cure parentali del quale abbiamo avuto occasione di osservare in natura (BRANDMAYR e BRANDMAYR-ZETTO, 1974). L'andamento dell'attività degli adulti sulla superficie del terreno presenta quei due picchi ancora più distanziati e, a differenza di Molops striolatus ,il massimo di comparsa si registra in autunno. In questa stagione infatti compare la gran massa degli immaturi, in questo caso riconoscibili per l'incompleta melanogenesi.
In conclusione, le cure parentali evolute di Molops sembrano connesse ad un ciclo biologico e ad un andamento fenologico fortemente modificato, comprendente un'estivazione, nel corso della quale si ha ovodeposizione in celle sotterranee. Che la scomparsa estiva non sia unicamente in funzione di questa modalità riproduttiva lo si evince dal fatto che, tutt'al più con un certo
ritardo, anche i maschi si infossano nel terreno, accompagnati per di più anche dalla frazione non sessualmente matura della popolazione. :È: poi interessante notare come alte temperature di stabulazione (20 T - 25 "C, esperimenti pre
cedenti a quanto riferito in questo lavoro) stimolino fortemente l'attività di scavo degli individui, i quali vengono poi ritrovati al fondo dei terrari in celle o gallerie, anche sotto dieci cm di spessore del substrato eO). Per
(') Nel Carso Triestino questa specie è rinvenibile sotto pietre anche in pieno inverno. eO) Durante la fase estiva i Molops si raccolgono sotto pietre profondamente infossate.
Questo ha probabilmente indotto JEANNEL (1949, in GRASSÉ, Traité de Zoologie) a definirli endogei, cosa che non è accettabile, poichè gli adulti sono attivi in superficie (catture con pit. traps non sarebbero altrimenti possibili).

o
15
lO
-5
evaporazione (mm/4 giorni)
o 'O " ..
E u
'" .! :ii DE Il ...
,
/',/ .... ,.. ..... ~".\,
max. /",---_.'" "\j
media
min.
ATTIVITA- STAGIONALE sul M. TAIANO nel
CARtel (HUMILlS)-SESLERETUM JUNCIFOLIAE
",'--1
r ..... /,' \ ,
, , .
'"" ... J
, I
;
di MOLOPS STRIOLATUS NELL' ANNO 1971
1,5
\.--- ..... '\ /'1 • ! \
\,,,,/ \.
"
,/ /\
, .', /',
I \.
'\,,/ "
\j 0,5
'. ".:
'. ".'------------\
.' . .....
60
40
20
Fig. XII
Attività degli adulti alla superficie del terreno del pascolo magro montano carsico. La scala dell'attività (linea grossa) si trova a destra ed è espressa in individui catturati! /trappola in quattro giorni. Le temperature hanno la scala a sinistra e riguardano esattamente lo strato d'aria nel quale gli individui sono attivi.

- 297-
questo genere di Carabidi è quindi forse quanto mai valida 1'ipotesi di una connessione tra cure parentali postnatali di tipo I ed un rallentamento dello sviluppo embrionale e di tutto il ciclo biologico, quale risposta a condizioni paleoclimatiche diverse dalle attuali, forse glaciali. L'estivazione del gen. Molops potrebbe essere la testimonianza di questo adattamento e, se non direttamente la causa del comportamento parentale, averne perlomeno favorito l'insorgere, in un'epoca meno recente che in altri Carabidi ad etologia consimile.
M. striolatus 100'/.
50 r-
r1- rapporto sessi ('4 o'd'adulti attivi
f-~~+-+-+-+-+-+-+-+---I--r- alla superficie)
~~ con uova mature
f----------- -f-------
G F M A M G L A S O N D
nuova generazione
pupe
III stadio
Il stadio
I stadio
ovideposizione
Fig. XIII
M. ovipennis 100 'I.
50
G F M A M G L A S O N D
Cicli biologici di M. striolatus ed ovipennis. Il rapporto sessi è basato sul conteggio di rispettivamente 81 e 125 esemplari (annata '71) abbondantemente confermato però da raccolte dirette effettuate per allevamenti e dissezioni. Le linee tratteggiate indicano fasi e periodi non suffragati da reperti o osservazioni.
La morfologia dell'apparato ovopositorecostituisce infatti un'ulteriore prova a favore del fatto che Molops è un genere a cure parentali primarie, nate con il genere stesso, o con i suoi primissimi rappresentanti.
La fig. XV rappresenta gli apparati ovopositori di alcuni Molopina (sensu JEANNEL, 1941-42 e DANELUTTO, 1975/76). In a è raffigurato quello di Molops striolatus, che è sostanzialmente uguale anche in M. ovipennis e pIitvicensis. Lo sterno VIII è profondamente diviso in due emisterniti, la membrana articolare VIII - IX è molto ridotta, al punto che per evidenziare il complesso stili-valviferi è stato necessario asportare parte d'un emisternite. Il valvifero è ridotto ad una sottile listerella sclerificata, priva di lobo posteriore.

- 298-
L'articolo basale dello stilo è molto sottile, quello distale è ridotto ad una semplice squametta priva di sensilli. Si tratta dunque di un ovopositore del tutto regredito, dove probabilmente l'unica parte a restare funzionale è lo sfintere vaginale, che aprendosi permette la caduta dell'uovo sul pavimento della cella-nido sotterranea. Praticamente dello stesso grado-la regressi~ne· in M. austriacus e piceus (f), dove il valvifero è meno ridotto, ma gli articoli dello stilo sono fusi. Il genere Tanythrix, a distribuzione geografica alpino-meridionale, presenta un apparato fortemente ridotto, con fusione del lobo posteriore alla parete membranosa del IX somite ed un articolo distale piccolissimo. T. edurus (e) e marginepunctatus hanno invece terminalia quasi normali, mancanti però dei sensillichetici. Stenochoromus montenegrinus (d), specie ad occhi ridotti e lapidicola del Montenegro e dell'Albania, ha un ovopositore perfettamente normale, con apodemi molto sviluppati e ricco di sensilli chetici. Speomolops sardous, Molopino cavernicolo ed anoftalmo di Sardegna, possiede un apparato normale, abbastanza sc1erificato in rapporto alla sua condizione ipogea, ricco di sensilli e dotato di due grosse spine sul lato esterno degli articoli distali degli stili. Typhlochoromus, genere endogeoed anoftalmo delle Prealpi Carniche presenta un ovopositore normale, almeno in base alle figure di PORRO e PORRO (1968).
L'interpretazione di questi dati modo-funzionali non può essere disgiunta, a nostro avviso, da un contesto storico-evolutivo. Mentre HORION (vedi pago 291) sostiene anche per il gen. Molops un'origine recente, JEANNEL (1924, 1950) ha sull'epoca di comparsa dei Molopina un'opinione ben diversa. Per l'autore francese infatti questi sarebbero un gruppo antico di faune terziarie originato si sull'Egei de, il quale avrebbe dato origine ad alcune linee proprie della Tirrenide, i cui relitti si trovano oggi nella catena Catalana ed alle Baleari (II) (Molopidius, Zariquieya, Henrotius) e ad alcune linee dinarico-alpine meridionali (Typhlochoromus, Henrotiochoromus, Tanythrix, Stenochoromus), ivi compresi i Molops attuali. A nostro avviso le vedute di JEANNEL si possono accettare se modificate con quelle di HORION, considerando cioè Molops e Abax i rami più recenti ed in fase di differenziazione di un gruppo, i Molopina, di origine cenozoica.
I generi più antichi, residui di faune mioceniche o anche premioceniche, a distribuzione relitta, fortemente discontinua, in gran parte anoftalmi,. endogei, cavernicoli oedafofili, sarebbero ad etologia più semplice,
( 1 ) Anche Speomo!ops sardòus PATRIZI, 1955, è da aggiungere alle «linee» della Tirrenide, probabile relitto di quei Molopini che secondo JEANNEL, 1950, hanno invaso la Mesogeide eocenica. Gli autori sono concordi nel considerare questo genere monobasico un relitto di gruppi a più ampia geonemia mediterranea oligocenica (FIORI, 1961) o comunque premiocenica (BACCETTI, 1964; CASALE, 1972).
l'C
-s
evaporazione ( m mi" giorni )
max. '. :' -. .;
:' "
./ '
I
l, "
~---- ./
ATTIVITA- STAGIONALE di
sul M. TAIANO nel
SESLERIO - FAGETUM
.-----,
___ o'
MOLOPS OVIPENNIS NELL' ANNO 1971
Fig. XIV
,
.'.
" /' \,
\. .. -., "
precipitazioni (mm/4 giorni) ....
1.5
1,0
0.5
Attività annuale degli adulti di M. ovipennis in una faggeta lievemente termofila dell'Istria montana. Per ulteriori particolari su metodologie ed ambiente di ricerca vedasi BRANDMAYR, 1974 a. Spiegazione dei simboli in fig. XII.

- 299-
vulva
Fig. XV
Ovopositori di vari Molopina: a, M. striolatus; b, Speomolops sardous; c, Tanythrix senilis; d, Stenochoromus montenegrinus; e, Tanythrix edurus; f, M. austriacus. Rappresentazione delle sclerificazioni ed abbreviazioni come nelle figg. III-IV.

- 300-
a cure parentali meno complesse. Lo si può desumere dalla morfologia degli ovopositori di Typhlochoromus, Stenochoromus, Speomolops. I ceppi più recenti - tardo Pliocene, Quaternario (12) - avrebbero modificato il proprio comportamento parentale, vegliando sulle uova in celle sotterranee. Questo cambiamento è stato sporadico e recente in Abax e Tanythrix {pro parte), con specie a cure parentali secondarie ed ovopositore poco ridotto, un po' meno recente in Molops (cure primarie, ovopositore fortemente regredito in tutte le specie) ed in Tanythrix senilis.
Altri argomenti a favore della differenziazione recente di Molops sono, oltre alle stesse cure parentali postnatali, il carattere sostanzialmente neoendemico degli areali di molte specie, la sua area di diffusione complessivamente molto ristretta, (europeo-medio e sud-orientale), il ciclo fenologico degli adulti comprendente un'estivazione e l'attività invernale (ricordo di epoche più fredde?).
Interessante ancora il caso di Tanythrix, nel quale coesistono specie ad ovopositore appena ridotto o molto regredito, a testimonianza di un'insorgenza polifiletica della veglia sulle uova. La comparazione delle figg. XV d, e, c, nell'ordine da l'idea della modificazione subita dall'ovopositore in relazione all'antichità della sua entrata \< in disuso» in un gruppo omogeneo. Stenochoromus e Tanythrix infatti appartengono a nostro avviso alla medesima linea evolutiva (si noti ad esempio la dentellatura sull'articolo distale dello stilo). In d (Stenoch.) un apparato presumibilmente funzionante, in e (T. edurus) uno caduto in disuso da poco, in c (T. senilis) uno fortemente regl'edito, e che si può quindi supporre in disuso da molto tempo.
Non è escluso che molte delle ipotesi sin qui formulate debbano essere modificate in futuro, alla luce di ulteriori ricerche. Quello che ci preme riaffermare in questo contesto è che le cure parentali sono senza dubbio una delle « chiavi» per la comprensione della radiazione adattativa dei Molopina. L'estensione degli studi morfofunzionali dell'ovopositore e dell'etologia parentale a tutti i generi di questo gruppo di Carabidi di origine senz'altro cenozoica, ma a forte espansione anche recente, porterà forse un po' di chiarezza in molti problemi storico-evolutivi sinora praticamente insoluti.
(U) Dove per quaternario s'intende quel periodo di 2-3 milioni d'anni nel quale giunge al suo acme il raffreddamento del clima, iniziatosi ancora nel Terziario antico. I periodi glaciali propriamente detti vennero preceduci, nel primo Quaternario, da altre oscillazioni climatiche, comprendenti anche fasi fredde che nella classificazione medio--europea son dette «Prategelen» (freddo), «Tegelen» (caldo), «Eburon» (freddo), quest'ultimo parzialmente corrispondente al Calabriano. Per maggiori dettagli vedasi SCHWARZBACH, 1974.

- 301-
Gen. Percus (osservazioni morfofunzionali).
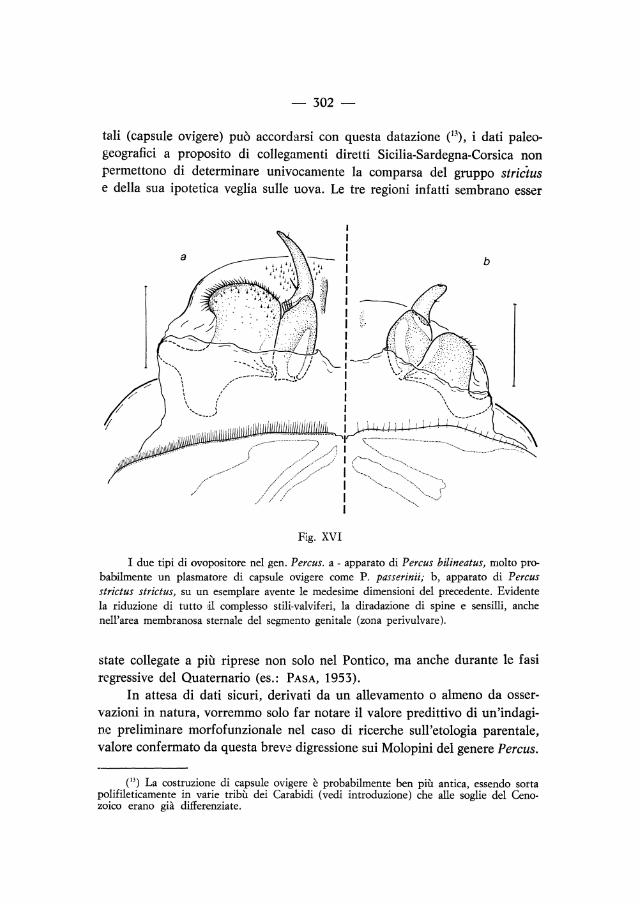
Forti dell'esperienza sui due generi precedenti, abbiamo voluto estendere l'indagine morfofunzionale dell'ovopositore al terzo dei grandi generi dei Molopina, limitandoci però in questa fase alle specie italiane. Percus occupa con le sue circa quindici specie l'area mediterranea occidentale, dalla penisola iberica agli Appennini, le grandi isole tirreniche e le Baleari, e, con il solo lineatus, anche Algeria e Tunisia. Le scarse conoscenze etologiche sono limitate, come già detto, a 2 specie, P. (Pseudopercus) navaricus dei Pirenei orientali e P. (s. str.) passerinii, dell'Appennino tosco-emiliano (LuMARET, 1971 risp. BARGAGLI, 1874). Contrariamente a quanto si sarebbe potuto pensare, anche in questo genere alcune specie possiedono un ovopositore ridotto. La maggior parte dei Percus ad VIII stria elitrale apicalmente infossata ha terminalia fortemente adattati alla costruzione di involucri terrosi a protezione delle uova, con forte sviluppo e sclerificazione del lobo posteriore del valvifero, che risulta sporgente e ricco di spine (fig. VIlla; fig. XVI a). Questo tipo morfologico è stato riscontrato in Percus dejeani, paykulli, linea
tus, villai, bilineatus, corsicus, passerinii. Diversa invece la struttura dell'ovopositore nel gruppo « strictus », a stria VIII anche apicalmente non infossata. Le tre entità tassonomiche controllate sono P. strictus strictus (Sardegna sett.), strictus lacertosus (Sicilia) e grandicollis(Sardegna settentrionale, Corsica). In questi Uìg. XVI b) stili e valviferi sono ridotti, le spine sono scarse, limitate alla parte più esterna del lobo posteriore, che è in pari tempo meno pronunciato, grossolanamente quadrato; sclerificazione e sensilli della zona membranosa perivulvare quasi assenti.
:t<: possibile quindi ipotizzare una «caduta in disuso» dell'ovopositore nel gruppo « siculo-sardo-corso » dello strictus, o perlomeno ad una diversa funzione di tale apparato.
Mancano sinora dati concernenti le cure parentali di queste specie. Oualora però in futuro si rivelasse che vegliano sulle uova in cripte sotterranee, sorgerebbero notevoli problemi circa l'epoca di comparsa di detto comportamento. Il genere Percus infatti è considerato « tipicamente paleotirrenico» (es.: FOCARILE, 1973; accenni anche in JEANNEL, 1942, p. 384) ed una
distribuzione siculo-sardo-corsa viene di solito ritenuta di origine molto an
tica, miocenica (LA GRECA, 1961).
Se da un lato la vasta distribuzione che hanno nel genere le cure paren-
- 302-
tali (capsule ovigere) può accordarsi con questa datazione e3), i dati paleogeografici a proposito di collegamenti diretti Sicilia-Sardegna-Corsica non permettono di determinare univocamente la comparsa del gruppo strictus e della sua ipotetica veglia sulle uova. Le tre regioni infatti sembrano esser
a b
-,
(:::::~'},'LLjj "
........ ___ ,1
.~ ....... ~ ......... -........ :
.....
Fig. XVI
I due tipi di ovopositore nel gen. Percus. a - apparato di Percus bilineatus, molto probabilmenre un plasmatore di capsule ovigere come P. passerinii; b, apparato di Percus strictus strictus, su un esemplare avente le medesime dimensioni del precedente. Evidente la riduzione di tutto dI complesso stili-valviferi, la diradazione di spine e sensilli, anche nell'area membranosa sternale del segmento genitale (zona perivulvare).
state collegate a più riprese non solo nel Pontico, ma anche durante le fasi regressive del Quatemario (es.: PASA, 1953).
In attesa di dati sicuri, derivati da un allevamento o almeno da osservazioni in natura, vorremmo solo far notare il valore predittivo di un'indagine preliminare morfofunzionale nel caso di ricerche sull'etologia parentale, valore confermato da questa breve digressione sui Molopini del genere Percus.
(") La costruzione di capsule ovigere è probabilmente ben più antica, essendo sorta polifileticamente in varie tribù dei Carabidi (vedi introduzione) che alle soglie del Cenozoico erano già differenziate.

- 303-
Gen. Platysma
Platysma (Melanius = Pseudomaseus) anthracinum ssp. hespericum
Buccia:rell,i e Sopracordevole.
Abbiamo potuto confermare anche per la ssp. italiana le osservazioni di LINDROTH (1946) sulla tipica, mediante il ritrovamento in natura di un esem
plare di sesso femminile, chiuso in una celletta subsferica scavata a 4 cm di profondità in terreno argilloso. Era circondata da 18 uova lunghe 1,5 -1,6 mm. Poste a 18°C, queste sono schiuse dopo 12 giorni. Data del ritrovamento: 4/VI/1974; biotopo: bosco planiziale della bassa pianura friulana (Querco-Carpinetum boreoitalicum, sec. P IGNATTI , 1953).
Dissezionata, la ~ dimostra di essere alla sua prima ovodeposizione: ovari color bianco, quasi diafani; corpi lutei appena accennati, molto chiari.
Delle due specie di Melanius sino ad oggi allevate, l'altra, cioè Pl. nigrita non presenta cure parentali evolute, ma il semplice infossamento delle uova mediante gli stili, come desumibile da THIELE, 1966, 1968. La comparazione delle morfologie degli ovopositori di tre specie di Melanius ci ha permesso di riscontrare significative differenze di sviluppo e struttura dei « terminali a »:
in anthracinum (fig. XVII b) le dimensioni relative degli stili e dei valviferi sono minori, gli articoli terminali degli stili sono corti e spatoliformi, l'insieme dei sensilli chetici è piuttosto regredito, a testimonianza della perdita di funzione dell'ovopositore. Non così in Pl. nigrita (fig. XVII c), gracile e minus dove dimensioni, sclerificazione e densità dei sensilli sono inalterate, come inalterata è probabilmente la funzionalità dell'apparato in questione. La veglia sulle uova in celle sotterranee di Pl. anthracinum è forse un caso isolato in
tutto il genere: Pl. vulgare e nigrum possiedono infatti ovodeposizione normale (THIELE, 1968 rispettivamente WITZKE, 1974).
Gen. Pterostichus
Pterostichus (Alecto) justusi W. Redtenbacher.
Abbiamo potuto raccogliere il 26 e 27/VI/1976 una dozzina di esemplari di questo interessante endemite stenotopo delle estreme propaggini orientali delle Alpi. È presente su due soli gruppi montuosi (per la distribu
zione geografica vedasi HOLDHAUS, 1954, p. 78 e tav. XVII): il Pohorie (m. 1543, a W. di Maribor, Slovenia) e la Koralpe (m 2140, ai confini tra Carinzia

- 304-
e Stiria). Gli esemplari sui quali si è basato l'allevamento provengono da quest'ultima località, un massiccio di rocce metamorfiche (gneiss e micascisti, vedi KLEINSCHMIDT e RITTER, 1976; BECK-MANNAGETTA, 1975) a sviluppata idrografia superficiale e ricco di falde acquifere affioranti. Pterostichus justusi, igrofilo e sistematicamente affine al ripicolo Pt. fasciatopunctatus, vi è presente dal piano alto-montano a quello alpino, concentrandosi particolar-
Fig. XVII
Ovopositori di Pterostichus e Platysma. a, Pterostichus (Cheporus) metallicus, specie che veglia sulle uova: le dimensioni sono praticamente normali, sensilli e fanere nel complesso un po' diradati; b, Platysma anthracinum, stilo e valvifero destro (contorni): articolo distale smussato ed accorciato rispetto a quello di altri Platysma, lobo posteriore del vf. ridotto, con poche spine e pochi sensilli; c, idem di Pl. nigrita, raffigurato come esempio di ovopositore non ridotto di Platysma.
mente lungo le rive dei torrentelli, nei canaloni umidi e dovunque l'elevata umidità del terreno non si accompagni ad una coltre umosa o torbosa troppo spessa. Gli esemplari provengono soprattutto dalla cresta che conduce al grande Speikkogel. Suddivisi in due gruppi di 3 d' d' e 3 ~ ~, sono stati posti in cristallizzatori di 20 cm di diametro, con per substrato argilla sabbiosa del luogo e scaglie di gneiss per riparo, alla temperatura di 16 - 17°C ed LD 15/9. La specie è particolarmente sensibile alla mancanza d'acqua, e molti individui affondano le mandibole nel terreno, bevendo avidamente,

- 305-
dopo ogni annaffiatura. Le osservazioni sul comportamento parentale si possono riassumere come segue:
una sola delle ~ ~ raccolte è risultata matura; l'ovodeposizione avviene nella fase oscura del ciclo giornaliero mediante la costruzione di capsule ovigere, che vengono attaccate ai lati o alla parete inferiore delle pietre;
tutti gli altri esemplari dei due sessi erano immaturi e per di più nella fase di formazione del corpo adiposo: si sono dovuti allontanare perchè distruggevano sistematicamente le capsule ovigere, divorandone l'uovo;
le uova complessivamente deposte dall'unica femmina-madre assommano ad almeno 25 in un periodo di circa due settimane;
lo sviluppo embrionale dura 15 giorni a 16 "C; la durata del primo stadio larvale è di circa 18 giorni alla medesima
temperatura.
Le capsule hanno un diametro massimo di 4-5 mm ed una forma non dissimile da quelle di altri Pterostichini, soprattutto del gen. Abax, cioè interamente chiuse, ad eccezione talora del lato verso il sostegno, che viene cosÌ a costituire una parete della cavità contenente l'uovo.
Si tratta del primo caso di costruzione di capsule ovigere noto per il gen. Pterostichus (sensu JEANNEL), e non è escluso che ricerche future rivelino anche in questo gruppo, come in Abax, una maggioranza di specie costruenti involucri terrosi a protezione delle uova. Il subgen. Cheporus (metallicus F.) è invece caratterizzato da cure parentali postnatali (WEIDEMANN, 1971), come pure gli affini Oreophilus. Uno sguardo all'apparato ovopositore di Pt. metallicus (fig. XVII a) e di Oreophilus multipunctatus permette di affermare che in queste specie la veglia sulle uova non è accompagnata dalle vistose regressioni di stili e valviferi osservate in M%ps. Questi anzi sono praticamente normali o tutt'al più impercettibilmente rimpiccioliti. f!. probabile dunque che, come in Abax, anche in Pterostichus le cure parentali evolute siano un'acquisizione secondaria, posteriore cioè alla formazione delle specie che le presentano.
CONCLUSIONI
Le conoscenze attuali sui comportamenti studiati sono ben lungi dall'essere complete. Nel corso di questa ricerca si sono comunque venuti delineando alcuni punti di rilievo, che si possono considerare una risposta, anche se provvisoria, ai quesiti formulati nell'introduzione (pag. 276) e soprattutto un punto di partenza per indagini future.

- 306-
Per quanto riguarda le relazioni tra cure parentali ed ecologia, l'esempio di Abax, che è sinora il più documentato, farebbe pensare ad una maggior capacità di adattamento ad ambienti diversi dei costruttori di capsule ovigere rispetto alle specie che vegliano sulle uova. Infatti il « primo gruppo », che sembra caratterizzato per intero dal primo comportamento, è costituito tanto da specie silvicole che da praticole. Tale varietà di scelte ecologiche potrebbe essere la conferma, estesa a tutto il genere, delle' minori esigenze edafiche osservate da LasER (1972) in A. ater rispetto ad A. parallelus. Essa sembra inoltre connessa, almeno nelle specie da noi studiate, a due significative varianti del comportamento nei confronti degli annessi del suolo: i praticoli infatti depongono le uova di preferenza sulla superficie inferiore delle pietre, in accordo con le caratteristiche ecoclimatiche delle formazioni aperte. La disparità numerica tra i due gruppi principali del genere (il secondo gruppo di FIORI, 1896, comprende solo 4 spp., tutte silvicole) ci invita per ora alla prudenza.
Paragonare l'etologia parentale di generi diversi, ponendola in relazione alle preferenze ambientali può essere scarsamente significativo, non solo perchè le conoscenze sulle esigenze edafiche di Molops, Oreophilus e Pterostichus sono molto scarse, ma anche perchè con il genere variano strutture ed adattamenti, cioè la « Lebensform» (14).
Resta comunque il fatto che le cure parentali evolute sembrano interessare soprattutto specie silvicole, o le cui popolazioni presentino perlomeno un massimo di abbondanza in biotopi boschivi, a distribuzione altitudinale montana o subalpina. La maggior parte degli Oreophilus è silvico!.o-euriecia, anzi negli orizzonti alto-montano e subalpino essi danno origine a popolazioni molto dense, al punto che AMIET (1967) ha chiamato « Oreophileti » i popolamenti a coleotteri terricoli di peccete e lariceti delle Alpi Marittime. L'eccezione più vistosa è forse costituita da PIatysma anthracinum, paludicolo e presente anche in pianura, prevalentemente silvicolo solo nelle parti meridionali o centrali del suo areale (MuLLER, 1926; LEHMANN, 1965; BRANDMAYR, 1975) ma che in quelle settentrionali (GERSDORF, 1937; LINDROTH, 1945) sembra tutt'al più favorito da un certo grado di ombreggiamento.
Il tratto ecologico unificante di questi Carabidi non è però tanto la formazione vegetale di appartenenza, o le condizioni edafiche, quanto forse il fatto che si tratta di specie attere o tutt'al più pteridimorfe (PI. anthracinum) viventi in ambienti (boschivi) stabili e duraturi. In questo contesto la veglia sulle uova può essere considerata un caso molto spinto di « selezione per K »,
(14) Anche altri AA. vedono ad esempio dn Molops un genere ad abitudini fossorie (AMIET, 1967, su M. medius). .

- 307-
ovvero una strategia adottata da popolazioni che per mantenersi alla densità di saturazione (15) nei loro biotopi elettivi puntano più su accorgimenti etologici tendenti alla diminuzione della mortalità preimmaginale che non su un alto potenziale riproduttivo. Già LAMPE (1975) ha posto l'accento sul fatto che specie esercitanti cure parentali postnatali depongono un numero di uova minore. Per restare nell'ambito dei Pterostichini basti ricordare le 153-216 uova di Calathus melanocephalus (VAN DIJK, 1972), le 50 e più uova che possono essere deposte da lj1 lj1 di Bothriopterus adstrictus in soli lO giorni (GOULET, 1974, p. 16, Tav. 7), il centinaio e passa di uova deposte da lj1 lj1
di Calathus fuscipes ed erratus (KURKA, 1972). Anche Platysma nigrita può totalizzare numeri elevati di uova deposte (THIELE, ex verbo in LAMPE, op. cit.). A tali cifre si contrappongono quelle molto meno elevate dei Pterostichini a cure parentali evolute: 15 in media per A. ovalis; un massimo di 30 per Pterostichus metallicus (WEIDEMANN, 1971) e quelle di 10-20 al massimo in prevalenza osservate o citate da BRANDMAYR e BRANDMAYR-ZETTO (1974, Tab. 1) ed in questo lavoro. Numeri d'uova particolarmente bassi sembrano caratterizzare Molops. Le scarse osservazioni sulla fertilità delle lj1 lj1 si possono così riassumere: M. piceus 12,12,10,1; austriacus 7; ovipennis 7; plitvicensis 5; striolatus 11, 5. Un tasso di natalità così basso permette di supporre che il tasso intrinseco di accrescimento (r) delle popolazioni di Molops sia pure basso, e ciò costituisce un argomento a favore di una strategia ecologica K-selezionata dei Pterostichini a cure parentali evolute. Quanto osservato si può facilmente porre in relazione con la longevità particolare di Molops (ciclo biennale) e con la forte regressione del loro ovopositore (comparsa meno recente delle cure parentali).
Ciclo biologico e cure parentali postnatali evolute sembrano strettamente connessi soprattutto nel genere Molops, la cui fenologia immaginale comprende un'estivazione, durante la quale ha luogo la veglia sulle uova in celle sotterranee. Altre specie, come Abax ovalis e Pterostichus metallicus, appartenenti alla medesima categoria etologica, non presentano però una marcata scomparsa estiva, (LAMPE, WEIDEMANN, op. cit.) ma tutt'al più un calo della popolazione attiva in superficie. In Molops infatti le cure sono primarie, formatesi cioè con i primi rappresentanti del genere stesso, forse in connessione con i primi mutamenti climatici del Pleistocene inferiore, e l'estivazione potrebbe quindi considerarsi un adattamento a climi più freddi dell'attuale.
Il ciclo di sviluppo biennale è di gran lunga il più diffuso nelle specie
(") L'indice K si riferisce all'equazione logistica di accrescimento delle popolazioni; per maggiori particolari si veda ad es. OnUM, Principi di Ecologia, Piccin, p. 259.

- 308-
a cure evolute sinora studiate (12 su 14) e6). Costituiscono eccezione PI. anthracinum ed A. parallelus, entrambi a riproduzione primaverile e comparsa autunnale della nuova generazione.
Gli studi morfofunzionali hanno permesso in numerosi casi di dedurre anche a priori se una specie faccia uso o meno dell'ovopositore. La costruzione di capsule ovigere può essere effettuata con un apparato « normale », non vistosamente modificato cioè rispetto a quello di altri Carabidi « Conchiferi» o « Balteiferi » (sensu JEANNEL, 1941-42). Nel genere Pereus questo comportamento sembra collegato ad un notevole sviluppo e spinosità del lobo posteriore del valvifero, importante per la raccolta del terriccio alla sua estremità e per il modellamento della cavità trasversale dell'uovo. Il gruppo siculo-sardo-corso dello strietus però ha un ovopositore parzialmente ridotto, e solo future indagini potranno stabilire se anche in questo genere a distribuzione paleotirrenica sia comparsa la veglia delle uova in celle sotterranee.
Le cure parentali evolute sono collegate a due diversi gradi di regressione dei terminalia femminili, in quanto implicanti la caduta in disuso dell'ovopositore. In certi casi si osserva una riduzione biometricamente accertabile di stili e valviferi (Abax, Platysma, Pereus gr. strietus), talora però solo la riduzione del corredo sensoriale (Oreophilus, Pterostiehus, Tanythrix pro parte). Molto più vistosi invece i casi di regressione avanzata di Tanythrix senilis e del gen. Molops, nei quali detto apparato sembra aver perso ogni qualsivoglia funzionalità, e dove gli stili possono anche essere fusi in un unico articolo (M. pieeused austriaeus). La tabella II illustra l'epoca relativa di comparsa della veglia sulle uova (immaginando che la velocità di regressione dell'apparato sia uguale in tutte le specie) ed il grado di riduzione dell'ovopositore nei Pterostichini le cui cure parentali evolute siano accertate. L'aggettivo « secondarie» indica naturalmente che le cure in quel caso sono posteriori alla formazione della specie. L'epoca di comparsa del fenomeno è a nostro avviso il Pleistocene, soprattutto per i motivi indicati nei precedenti capitoli a proposito della fenologia. Non è escluso però che nuove osservazioni (Pereus gr. strietus!) ci obblighino in futuro a prendere in considerazione datazioni più antiche. Gli aggettivi più e meno recenti si riferiscono quindi sempre a quest'era geologica.
La veglia sulle uova in celle sotterranee sembra essere una peculiarità dei Pterostichini. Il diverso grado di riduzione dell'ovopositore nelle specie che la presentano, il caso di A. ovalis e parallelus e quello isolato di Platysma
(16) L'alto numero di larve al III stadio di Oreophilus da noi rinvenute in alta montagna nel mese di giugno ci permette di attribuire anche a questo genere un ciclo di sviluppo biennale.

- 309-
anthracinum fanno ritenere che l'insorgenza polifiletica sia tutt'ora in pieno svolgimento. All'interno del gruppo esse sembrano caratterizzare le subtribus Molopina e Pterostichina, aventi in comune quindi una medesima «predisposizione genetica ». :t interessante notare inoltre come proprio nei generi
TABELLA Il
epoca di rapporti
Specie comparsa filetici
Ovopositore ± recente
Platysma anthracinum + secondarie lievemente ridotto
Pterostichus metallicus (*) + secondarie quasi normale, pochi sensilli
Oreophilus multipunctatus + secondarie quasi normale, pochi sensilli Oreophilus morio + secondarie quasi normale, pochi sensilli Oreophilus yvani + secondarie quasi normale, pochi sensilli
Abax ovalis + secondarie sclerificazione e dimensioni ri· dotte.
Abax parallelus + secondarie sclerificazione e dimensioni ri· dotte.
T anythrix edurus + secondarie quasi normale, spine e sensilli ridotti
T anythrix senilis secondarie fortemente regredito
Malaps piceus primarie fortemente regredito Malaps austriacus primarie fortemente regredito Molops avipennis primarie fortemente regredito Malaps plitvicensis primarie fortemente regredito Malaps strialatus primarie fortemente regredito
(*) Dobbiamo al Prof. FRIEDRICH WEBER (Miinster, Germania Federale) un completamento della tabella a manoscritto inoltrato: Pt. (Cheparus) muhlfeldi, endemico delle Alpi Orientali, esercita pure la veglia sulle uova. Il sottogenere bibasico Cheparus è quindi interamente dedito a cure parentali evolute.
Abax e Molops, etologicamente complessi, anche gli organi genitali interni presentino un alto grado di differenziazione, in particolare nei vasi deferenti (LAMPE, 1968; LaSER e LAMPE, 1973).
La veglia sulle uova rappresenta in questo gruppo di Carabidi evoluti (i Pterostichini), attualmente in fase di espansione e rappresentato a livello mondiale da ben 5.000 specie (JEANNEL, in GRASSÉ, 1949) un mutamento della strategia riproduttiva. Basato su una modificazione del comportamento

- 310-
materno, nelle specie che lo presentano da un certo tempo (Molops) esso si accompagna ad un ciclo fenologico peculiare, comprendente un'estivazione. Ricerche eco-fisiologiche e di dinamica di popolazioni, unitamente all'estensione delle osservazioni sull'ovodeposizione di altri Pterostichini si rendono ora necessarie per il completamento di un quadro che è delineato solo in alcuni suoi tratti essenziali.
RINGRAZIAMENTI
L'autore ringrazia quanti hanno in vario modo collaborato al buon esito della ricerca. Si sente obbligato in particolare verso il Prof. ATHOS GOIDANICH (Torino) ed il Prof. HANS ULRICH THIELE (K61n) per i continui incitamenti allo studio delle cure parentali dei Carabidi, verso la moglie TULLIA per l'aiuto prestato negli allevamenti e la dott.a FABIOLA DANELUTTO per i dati sul gen. Abax. Esprime inoltre la sua gratitudine al prof. SANDRO RUFFo. direttore del Museo Civico di Storia Naturale, Verona, per il copioso materiale messo a disposizione ed all'ing. GUIDO CALLIGARIS, con il quale ha potuto discutere vari aspetti della biologia dei Pterostichini. E infine riconoscente al prof. ELVEZIO GHIRARDELLI, direttore dell'Istituto di Zoologia ed Anatomia Comparata di Trieste.
RIASSUNTO
Ricerche sul comportamento mediante allevamenti sono state affiancate da uno studio morfofunzionale degli apparati ovopositori per chiarire alcuni aspetti delle cure parentali nei Carabidi Pterostichini. Delle 5 specie di Abax che si è riusciti a portare all'ovodeposizione, A. carinatus interra semplicemente le uova, A. exaratus, biinningeri ed oblongus risultano costruttori di capsule ovigere di argilla a difesa di uova singole, A. ovalis (popolazioni alpino-orientali) esercita cure postnatali evolute, consistenti nella veglia sulle uova in celle sotterranee, come aveva già osservato LAMPE (1975) per la stessa specie nello Schiefergebirge.
Sono stati approfonditi alcuni aspetti della costruzione di capsule ovigere, quali la sequenza degli atti materni, la forma degli involucri, la scelta del substrato e la differente disposizione delle uova sulla superficie superiore (silvicoli) o inferiore (praticoli) delle pietre. Gli studi sull' ovopositore di sostituzione di questi Coleotteri dimostrano che l'insorgenza della costruzione di capsule ovigere nei Pterostichini (ed in altri gruppi dei Conchifera e Baltei/era di JEANNEL, 1941-42) può essere interpretata anche come preadattamento morfofunzionale (mancanza del lobo posteriore del valvifero ad es. nei Carabini). Nel gen. Abax inoltre si è potuto accertare biometricamente che solo due specie su 17, essendo dotate di cure evolute, presentano un apparato ovopositore ridotto (ovalis e parallelus) e si è chiarita la distribuzione dei comportamenti in tutto l'albero filogenetico, ponendoli in relazione all'ecologia, alla sistematica ed alla storia evolutiva.

- 311-
Molops striolatus ha rivelato cure parentali postnatali evolute come gli altri rappresentanti del genere sinora studiati. Nei Molops la veglia sulle uova si accompagna ad un ciclo fenologico peculiare, comprendente un'estivazione degli adulti (durante la quale le
'i? 'i? «covano» le uova). La fenologia cosi modificata, testimonianza forse di epoche più fredde durante le quali il genere si sarebbe differenziato, si può facilmente porre in relazione con il fatto che in questi Carabidi le cure parentali sono primarie, presenti cioè già nei rappresentanti ancestrali, come si può desumere dal forte grado di regressione di stili e valviferi. Lo studio morfologico comparato dei segmenti genitali di altri Molopini, anche delle forme relitte derivanti da linee paleotirreniche premioceniche (es.: Speomolops) o egeiche (Stenochoromus, Typhlochoromus) ha permesso di formulare l'ipotesi che le cure parentali evolute siano una conquista dei Molopina (s. ]EANNEL) recenti, ad espansione tardo-pliocenica e pleistocenica, mentre le forme antiche, ad ovopositori non ridotti, sarebbero ad etologia più semplice.
Nel terzo vasto genere dei Molopini, Percus, un'indagine morfologica a scopo predittivo sui genitali esterni femminili permette di individuare almeno due modalità di ovodeposizione: la più diffusa sarebbe la costnlzione di capsule ovigere, già constatata da vari AA. in due specie e corrispondente ad un notevole sviluppo del lobo posteriore del valvifero. Nel gr. strictus però ci sono ovopositori ridotti, che fanno pensare a cure parentali evolute.
Veglia sulle uova è stata riscontrata anche nella ssp. italiana (hespericum) di Platysma anthracinum. Sembrerebbe però trattarsi di un caso isolato all'interno del genere, com'è confermato dall'esame morfologico comparato dei genitali femminili.
Pterostichus justusi infine, endemismo della Koralpe (Carinzia), si è rivelato costruttore di capsule ovigere e non si esclude che tale modalità di ovodeposizione sia molto diffusa in specie e sottogeneri affini.
Sono inoltre approfonditamente discusse le questioni riguardanti il significato delle cure parentali evolute in relazione a ciclo biologico, ecologia, epoca di comparsa, grado di riduzione dei « terminalia » ed ai rapporti filetici. La veglia sulle uova compare polifileticamente negli evoluti Pterostichina e Molopina e sembra rappresentare un mutamento di strategia riproduttiva selezionato in ambienti stabili (soprattutto silvicoli) e tendente alla diminuzione della mortalità preimmaginale. Il fatto che si tratti di specie K-selezionate troverebbe la sua riprova nella generale longevità (prevalenza di cicli biennali) e soprattutto nel basso numero di uova deposte, corrispondente ad un tasso intrinseco di accrescimento molto basso delle popolazioni. La veglia sulle uova è inoltre da considerarsi secondaria e più recente in Abax, Platysma. Oreophilus (?), Tanythrix edurus; secondaria e meno recente in Tanythrix senilis; primaria e meno recente in Molops.
SUMMARY
ETHOLOGICAL AND MORPHO·FUNCTIONAL INVESTIGATIONS ON PARENTAL BEHAVIOUR IN PTEROSTICHINE CARABIDS (COLEOPTERA, CARABIDAE:
PTEROSTICHINI).
To elucidate many problems of parental care in Pterostichini ethological research by means of rearing was carried out togheter with morpho·functional studies on non-gonapophyseal ovipositors. The ovipositions have been observed in five Abax species: A. carinatus shows primitive egg-Iaying behaviour, i. e. the direct pushing of eggs in earth; A. exa-

- 312-
ratus, bi:inningeri and oblongus encase its eggs in mud cells which were attached to stones, mosses or other litter-components (fig. I); A. ovalis (fig. VI, populations of eastern Alps) shows high1y evolute brood care beyond oviposltion (<< Brutpf1ege »), as already observed by LAMPE (1975) on hills near K61n. Behavioral sequences shown by females and especially the shape of «ovigerous capsules» have been investigated (figs. I, II) and active choiee of c1ayey substrate rather than humus seems to be ascertained for A. exaratus. Alpine field-species as A. oblongus fix their capsules with preference to the lower surface of stones, silvieolous ones indifIerent1y, according to the ecoc1imatic difIerences between open and c10se vegetal formations. Studies on egg-Iaying apparatus show that construction of mud cells in Pterostichini (like in other groups of Conchifera and Balteifera of JEANNEL, 1941-42) can be explained in terms of morpho-functional pre-adaptation: in Carabini, for example, a posterior lobe of valvifer is not developed (strict1y «digging apparatus », fig. IV, 1, 2); on the contrary the ovipositor of Pterostichini, with well formed posterior lobes (fig. III, lb. post.), can be denominate a «moulding apparatus» (fig. III). In the whole genus Abax (17 spp.) only 2 species (ovalis and parallelus) show a biometrie reduction of styli-valviferi complex (fig. VII), according to their «Brutpf1ege », which implies inactivity of ovipositor. Distribution of the three behavioral patterns in phylogenetic tree was explained with regard to ecology, systematies and phyletic relationships, (figs. VIII, IX).
Brood care of Molops striolatus has been shown to be postnatal as well as in other species of this genus (figs. X, XI). In Molops the watch on eggs becomes a patt of a characteristic phenologieal pattern inc1uding imaginal aestivation (figs. XII-XIV). Modified phenology could be a record of cooler c1imatic stages, during which the genus has been differentiated, and seems to be in connectlOn with strong regression phenomena in female genital structures, i. e. with primary features of brood care in Molops (fig. XV). Comparative morphological analysis of terminalia has also been made on other Molopina, especially on paleotyrrhenian premiocenic (Speomolops) and aegeie relics (Stenochoromus, Typhlochoromus). This comprehensive view allows the hypothesis that postnatal brood care is an evolutionary target reached only by recent JEANNEL'S Molopina, whereas ancestors, with normal genitalia, show simpler behaviour.
In Percus (the third large genus of Molopina) a previsional study of female genital segments leads to two oviposition behavioral patterns. The more frequent mud celI construction is known already for 2 species and appears to be coupled with well developed posterior lobes of valvifera (fig. XVI). However P. strictus-group has reduced ovipositors and it suggests «Brutpf1ege ».
Guarding eggs of Platysma anthracinum and ltalian ssp. hespericum (hoc opus) seems to be unique within Platysma, as confirmed from ovipositor's evidence (fig. XVII).
Mud cell construction has also been observed in Pterostichus justusi, which lives endemie on Koralpe (Carinthia) and Pohorie (Slovenia). This ovipositional pattern is probably very frequent in related species and subgenera.
The significance of post-natal maternalistie behaviour has been discussed with partieular reference to life cyc1e, ecology, geologie period of appearance, reduction degree of terminalia and phyletie relationships (see table II). Watch on eggs arises polyphyle
tically in evolute Pterostichina and Molopina and seems to be a change in reproduction strategy, whieh is due to selective pressure acting in stable forest habitats and tends to lower preimaginal death rate. Small number of eggs (= low intrinsic growth-rate of populations) and longevity (high frequence of 2 years life cyc1es) agrees with situation of these
K-selected species.

- 313-
Post-natal brood care is regarded as secondary and more recent in Abax, Platysma, Pterostichus (?), Oreophilus (?), Tanythrix edurus; secondary and less recent in Tanythrix senilis; primary and less recent in Molops. Future investigations however, especially on Percus, could partly modify these attainments.
BIBLIOGRAFIA
AMIET ].1., 1967 - Les groupements des wlcopteres terricoles de la haute vallée de la Vesubie (Alpes Maritimes). - Mem. Mus. Hist. Nat. Paris, S.A, Zoologie, 46 (2): 124-213.
BACCETTI B., 1964 - Considerazioni sulla costituzione e l'origine della fauna di Sardegna. - Lavv. Soc. It. Biogeografia, VIII (1963/64) - Arch. Bot. Biogeogr. It. 40, 4a S., 9 (4): 37-103.
BARGAGLI P., 1874 - Cenni biologici su due specie di Percus. - BolI. Soc. Ent. It. 6: 27-3l. BEcK-MANNAGETTA P., 1975 - Der geologische Aufbau des steirischen Anteils der Koralpe.
- Ber. wasserwirtsch. Rahmenplanung, 31:8-41 (con cartina 1:50.(00). BOLDORI 1., 1933 - Appunti biologici sul Ptetostichus multipunctatus. - Studi Trent. Sco
Nat., Trento, 14: 222-233. BRANDMAYR P., 1974 a - Auto- und Synokologische Untersuchungen iiber die Carabiden
zweier Vegetationseinheiten des Slowenischen Kiistenlandes: das Carici (humilis)Seslerietum juncifoliae und das Seslerio (autumnalis)-Fagetum (Coleopt. Carabidae). -Acta ent. Jugoslavica, lO (1-2): 15-40.
BRANDMAYR P., 1974b - Le cure parentali di Carterus (Sabienus) calydonius Rossi (Coleoptera, Carabidae). - Atti Acc. Sco Torino, 108: 811-818.
BRANDMAYR P., 1975 - Un gruppo di invertebrati del suolo, i Coleotteri Carabidi, in relazione al grado di trasformazione dei biotopi agrari e forestali del basso Friuli: sua importanza per la ricostruzione ambientale. Ecologia ed Agricoltura: Problemi delle aree abbandonate. - (Riunione del Gr. di lavo per l'ecologia della Soc. Bot. It., Firenze, 22-23/XIj1974, in: Inform. Bot. It., 7: 91-97.
BRANDMAYR P., 1976 - Revisione del gen. Abax Bonelli (Coleoptera, Carabidae) su base morfo-funzionale, etologica ed ecologica. In c. di st. su Atti XI Congr. Naz. Ent.
Portici-Sorrento, 10-15jV/1976. BRANDMAYR e BRANDMAYR-ZETTO, T., 1974 - Sulle cure parentali e su altri aspetti della
biologia di Carterus (Sabienus) calydonius Rossi, con alcune considerazioni sui fenomeni di cura della prole sino ad oggi riscontrati in Carabidi (Coleoptera, Carabidae). -Redia, 55: 143-175.
BURMEISTER F., 1939 - Biologie, (jkologie und Verbreitung der europilischen Kilfer. - I Band: Adephaga. 307 pp., Goecke, Krefeld.
CASALE A., 1972 - Visione d'insieme del complesso ecologico e faunistico della grotta del Bue Marino (Cala Conone, Dorgali, NU). - BolI. Soc. Sarda Sc. Nat., 10:3-28.
CLAASSEN P.W., 1919 - Life history and biological notes on the Chlaenius impunctifrons Say. (Coleoptera, Carabidae). - Ann. Ent. Soc. Amer. 12: 95-100.
DANELUTTO F. - Revisione del genere Abax Bonelli su base morfofunzionale ed etologica con particolare riguardo agli apparati genitale ed ovipositore. - Tesi di laurea (inedita) in Sc. Nat., Univo Trieste (1975-76).
DICKER G.H.L., 1951 - Agonum dorsale Pont. (Col., Carabidae): an unusal egg-laying habit and some biological notes. - Ent. Mon. Mag., 87: 33-34.

- 314-
DI]K Th. S. (VAN), 1972 - The signilicance 01 the Viversity in Age Composition 01 Calathus melanocephalus L. (Col. Car.) in Space and Time at Schiermonnikoog. - Oecologia, lO: 111-136.
ERVIN T.L., 1967 - Bombardier beetles (Coleoptera, Carabidae) 01 North America: Parto II. Biology and behavior 01 Brachinus palliJus Erwin in Calilornia. - Coleop. Bull., 21: 41-55.
FIORI A., 1896 - Note critiche sulle specie del primo gruppo del gen. Abax. - Atti Soc. Nat. Modena, Ser. II, 29: 28-42.
FIORI G., 1961 - Alcuni appunti sull'entomolauna cavernicola sarda e sui problemi concernenti il popolamento delle caverne della Sardegna. - Atti Ace. Naz. !tal. Entom., Rendiconti, 8: 307-316.
FOCARILE A., 1973 - Cenni sugli aspetti zoogeogralici della coleotterolauna orolila dell'Appennino centrale. - Atti Ace. Se. Torino, 1" Classe Se. Mat., Fis., Nat. 107: 565-588.
FRANZ H., 1929 - Morphologische und phylogenetische Studien an Carabus L. und deT niichstveTwandten Gattungen. - Z. wiss. Zool., 135: 163-213, 56 fìgg.
GERSDORF E., 1937 - dkologisch-Iaunistische Untersuchungen uber die Carabiden der mecklenburgischen Landschalt. - Zool. Jahrb., Abt. f. Syst., 70: 17-86.
GOULET H., 1974 - Biology and relationships 01 Pterostichus adstrictus Eschscholtz and Pterostichus pensylvanicus Le Conte (Coleoptera, Carabidae). - Quaest. EntomoI., lO (1): 3-33.
HOLDHAus K., 1954 - Vie Spuren der Eiszeit in der Tierwelt Europas. - Abhandl. Zool. Bot. Ges. Wien, 18; 493 pp., 52 tavv.
JEANNEL R., 1924 - Coléoptères nouveaux de Catalogne. - PubI. Junta Cjencias Nat. Barcellona, 4 (8): 3-11.
JEANNEL R., 1941-42 - Faune de France. Coléoptères Carabiques. - 39, premiere partie; 40, deuxieme partie. 1172 pp., Paris, Librairie Fac. S., reprint Kraus, 1975.
JEANNEL R., 1942 - La genèse des faunes terrestres. - Bibl. Inst. Maritime et Coloniale, Presses Univ., Paris, 514 pp., 213 fìgg., 8 tavv.
JEANNEL R., 1948 - Sur deux larves de Carabiques. - Rev. Frane. d'Entom., Paris, 15: 74-78. JEANNEL R., 1949 - Drdre de Coléoptères, in P. GRASSÉ; Traité de ZooI., Anat. Syst.
BioI., VoI. 9: 771-1077. JEANNEL R., 1950 - Sur deux Ptérostichides cavernicoles de Ma;orque. - Rev. Frane. d'En
tom., 17 (3): 157-165. KING J.L., 1919 - Notes on the bio!ogy 01 thc Carabid genera Brachynus, Galerita and
Chlaenius. - Ann. Amer. Ent. Soc. COlumbus, 12: 382-388. KLEINSCHMIDT G. e U. RITTER., 1976 - Geologisch-petrographischer Aulbau des Koral
penkristallins sudich von Soboth / Steiermark-Kiirnten (Raum Huhnerkogel-La(Jl~en). -Carinthia II, 86: 57-91.
KOMAREK J., 1954 - Materska pece u Molops piceus Panz. (Brutpllege bei Molops piceus Panz.). - Acta Soc. Ent. Cechoslov., 51: 132-134.
KRAUSE R., 1974 - Vie Laulkiiler der Siichsischen Schweiz, ihre Phiinologie, dkologie und Vergesellschaltung. I (Coleoptera, Ckindelidae et Carabidae). - Faun. AbhandI., 5 (1): 73-179.
KURKA A., 1972 - Bionomy 01 the Czechoslovak species 01 the genus Calathus Bon., with notes on their rearing (Coleoptera; Carabidae). - Vesto Cs. spoI. zooI. 36 (2): 101-114.

- 315-
LA GRECA M., 1961 - Considerazioni sull'origine e la costituzione della Fauna di Sicilia. -Arch. Bot. e Biogeogr. It., 37, Ser. 4a; 6 (4): 1-23.
LAMPE KH., 1968 - Untersuchungen zur Morphologie der inneren Geschlechtsorgane von Carabiden. Hausarbeit zur ersten Staatspru;ung fur das Lehramt an der Realschule. -Kéiln, Germania Federale.
LAMPE KH., 1975 - Die Fortpflanzungsbiologie und Okologie des Carabiden Abax ovalis DFT. und der Einfluss der Umweltfaktoren Bodentemperatur, Bodenfeuchtigkeit und Photoperiode auf die Entwicklung in Anpassung an die Jahreszeit. - Zoo1. Jb. Syst., 102: 128-170.
LEHMANN H., 1965 - Okologische Untersuchungen uber die Carabidenfauna des Rheinufers in der Umgebung von Koln. - Z. Morph. Oko1. Tiere, 55: 597.
LENGERKEN H. (VON), 1954 - Die Brutfursorge und Brutpflegeinstinkte der Kiifer. II ed., pp. 383, Geest & Portig, Leipzig.
LEONARDI c., 1969 - Descrizione delle larve di Molops senilis Schaum e Typhlochoromus stolzi (Mocz.) e note sulla sistematica larvale di questi due generi di pterostichini (Coleoptera, Carabidae). - Ann. Mus. Civ. St. Nat., Genova, 77: 609-625.
LINDROTH C.H., 1945 - Die Fennoskandischen Carabidae. Spezieller Teil. - Kung1. Vetenskap. Vitterh. Samh. Handling., Ser. B, 4 (1): 709 pp.
LINDROTH CH., 1946 - Inheritance of wing dimorphism in Pterostichus anthracinus 111. -Hereditas, Lund, 32.
LINDROTH C.H., 1957 - The principal termr used for male and female genitalia in Coleoptera. - Opuscula ent., 22: 241-256.
LaSER S., 1970 - Brutfursorge und Brutpflege bei Laufkiifern der Gattung Abax. Zoo1. Anz. Supp1. 33, Verh. Zoo1. Ges., 358-364.
LaSER S., 1972 - Art. und Ursachen der Verbreitung einiger Carabidenarten (Coleoptera) im Grenzraum Ebene-Mittelgebirge. - Zoo1. Jb. Syst., 99 (2): 213-262.
LaSER S. e K.H. LAMPE., 1973 - Die Morphologie und Histologie der Vasa deferentia von Abax ater ViU. (Col. Carab.) und die in ihnen stattfindende Spermiozeugmenbil. dung. - Verh. Deutsch. Zoo1. Ges., Mainz. 66. Jahresvers., pp. 83-87.
LUMARET J.P., 1971 - Cycle biologique et comportement de ponte de Percus (Pseudopercus) navaricus (Col. Carabique). - L'Entomologiste, 27 (3): 49-52.
MARCUZZI G., 1952/53 - Osservazioni sulla microsistematica di alcuni Coleotteri delle Dolomiti. - Atti 1st. Veneto Se. Lett. Arti, 3: 209.
MARCUZZI G., 1962 - L'endemismo nella fauna delle Alpi Orientali. - Arch. Bot. Biogcogr. It., 28: 1-23.
NooNAN G.R., 1973 - The Anisodactylines (Insecta: Coleoptera: Carabidae: Harpalini): Classification, evolution and zoogeography. - Quaest. Entomo1., 9 (4): 266-480.
MiiLLER G., 1926 - I Coleotteri della Venezia Giulia. Catalogo ragionato. Parte la, Adephaga. - Studi entomologici, Trieste, 1 (2): 1-306.
PASA A., 1953 - Appunti geologici per la pc:teogeografia delle Puglie. - Mem. Biogeogr. Adriatica, 2: 175-286.
PATRIZI S., 1955 - Nuovo genere e nuova specie di Pterostichide troglobio della Sardegna orientale (Coleoptera Carabidae, Fam. Pterostichidae, Trib. Molopini). - Fragm. Entomo1., 2 (4): 29-40.
PIGNATTI S., 1952/53 - Introduzione allo studio fitosociologico della pianura veneta orientale con particolare riguardo alla vegetazione litoranea. - Arch. Bot., 28: 265-329;
29: 1-25/65-98/129-174.

- 316-
PORRO E. e G. PORRO, 1968 - Contributo alla conoscenza del genere Typhlochoromus (Coleoptera, Carabidae). - BolI. Soc. Ent. It., 98 (1-2): 19-29.
SCHWARZBACH M .. 1974 - Das Klima der Vorzeit - Eine Einfuhrung in die Palaoklimatologie. 3' ed., 380 pp., 191 figg. - Enke Verlag, Stuttgart.
THIELE H.V., 1956 - Die Tiergesellschaften der Bodenstreu in den verschiedenen Waldtypen des Niederbergischen Landes. - Z. angew. Ent., 30: 316-367.
THIELE H.V., 1964 - Experimentelle Vntersuchungen uber die Ursachen der Biotopbindung bei Carabiden. - Z. Morph. okol. Tiere, 53: 387-452.
THIELE H.V., 1966 - Einflusse der Photoperiode auf die Diapause der Carabiden. - Z. angew. Ent., 58: 143-149.
THIELE H.V., 1968 - Zur Methode der Laboratoriumszucht von Carabiden. - Decheniana, 120: 335-34l.
THIELE H.V., 1971 - Die Steuerung der Jahresrhytmik von Carabiden durch exogene und endogene Faktoren. - Zool. Jb. Syst., 98: 341-37l.
THIELE H.V., 1973 - Physiologisch-okologische Studien an Laufkafern zur Kausalanalyse ihrer Habitatbindung. - Verh. Ges. f. Okol., Saarbriicken, pp. 39-54.
VIGNA-TAGLIANTI A., 1973 - Considerazioni sui Carabidi cavernicoli ed endogei dell'Asia Minore (Col. Car.). - Int. J. Speleol., 5 (3/4): 349-360.
WEIDEMANN G., 1971 - Zur Biologie von Pterostichus metallicus F. (Coleoptera, Carabidae). - Faun. Oekol. Mitt., 4 (1): 30-36.
WITZKE G., 1974 - Die Larve von Pterostichus (Platysma) niger (SCHALLER, 1783). - Entom. Blatter, 70 (1): 5-ll.
XAMBEU., 1900 -Moeurs et métamorphoses des Insectes. Rev. d'Ent., 19: 1-56.
Copyright © 2022 FDOKUMEN




















