
Delayed autonomous self-pollination in two Japanese varieties of E pipactis helleborine...
11
Delayed autonomous self-pollination in two Japanese varieties of Epipactis helleborine (Orchidaceae) KENJI SUETSUGU* Graduate School of Human and Environmental Studies, Kyoto University, Yoshida-Nihonmatsu-cho, Sakyo, Kyoto 606-8501, Japan Received 22 December 2012; revised 5 July 2013; accepted for publication 16 August 2013 The terrestrial orchid Epipactis helleborine is a morphologically variable species with a wide geographical distribution. It is found throughout Europe and continues eastwards to Siberia, China and Japan. It is usually pollinated by social wasps and displays the morphological characteristics of an outcrossing species. In warm, temperate areas of Japan, E. helleborine often appears in alpine or subalpine regions, and has never been found in low-altitude forests, except for coastal pine forests. The coastal population of E. helleborine is often classified at the variety level, as E. helleborine var. sayekiana, and the inland populations are known as E. helleborine var. papillosa. It is possible that E. helleborine var. sayekiana possesses a distinctive selfing strategy, as its autonomous self-pollination has evolved in dry habitats, such as coastal dunes. The present study investigated the pollination biology of E. helleborine var. sayekiana and var. papillosa to detect differences in their reproductive systems. Unexpectedly, both E. helleborine var. papillosa and E. helleborine var. sayekiana were found to possess a self-pollination strategy and were therefore rarely visited by insects. Self-pollination occurs at the end of the flowering season and probably acts to ensure pollination even if insect-mediated pollination fails. Moreover, there are no floral differences between E. helleborine var. papillosa and E. helleborine var. sayekiana. These observations suggest that it is not necessary to distinguish between these two varieties. Furthermore, E. helleborine var. papillosa was pollinated by hoverflies, suggesting that E. helleborine var. papillosa is probably biologically distinct from its mother species, E. helleborine. © 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 733–743. ADDITIONAL KEYWORDS: autogamy – gynostemium morphology – pollination biology – reproductive assurance – reproductive biology. INTRODUCTION It has been noted that some taxa may be accorded protection unjustified by the biological reality (Pillon & Chase, 2007). This occurs more frequently for endangered species in taxonomically complicated groups (Pilgrim, Crawley & Dolphin, 2004; Ennos et al., 2012). One example is the genus Epipactis Zinn (Orchidaceae), with 25 (Richards, 1982), 36 (Delforge, 1995) or up to 56 (Delforge, 2001) species. Breeding systems, which include anatomical, morphological and biological aspects of the generative reproduction of individuals and populations, are an important characteristic used for the identification of such prob- lematic taxa (Pedersen & Ehlers, 2000; Squirrell et al., 2002; Bonatti, Sgarbi & Del Prete, 2006; Talalaj & Brzosko, 2008; Jakubska-Busse et al., 2012). Incomplete understanding of the breeding system of Epipactis is one of the main reasons for the taxonomic difficulties in this group (Talalaj & Brzosko, 2008). This phenomenon is exemplified by the Epipactis helleborine (L.) Crantz complex, which has radiated extensively throughout Eurasia and North Africa and is a widely established alien in North America. It is one of the most widespread orchid species (Luer, 1975; Delforge, 1995). This species is the progenitor of many problematic allogamous and autogamous sub- groups distinguished by early systematic monographs (Keller & Schlechter, 1940), which are still being modified today (Harris & Abbott, 1997; Squirrell et al., 2002). Despite the consensus that allogamy is predominant, the details of the breeding system for *Corresponding author. E-mail: [email protected] Botanical Journal of the Linnean Society, 2013, 173, 733–743. With 5 figures © 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 733–743 733
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Delayed autonomous self-pollination in two Japanese varieties of E pipactis helleborine...

Delayed autonomous self-pollination in two Japanesevarieties of Epipactis helleborine (Orchidaceae)
KENJI SUETSUGU*
Graduate School of Human and Environmental Studies, Kyoto University, Yoshida-Nihonmatsu-cho,Sakyo, Kyoto 606-8501, Japan
Received 22 December 2012; revised 5 July 2013; accepted for publication 16 August 2013
The terrestrial orchid Epipactis helleborine is a morphologically variable species with a wide geographicaldistribution. It is found throughout Europe and continues eastwards to Siberia, China and Japan. It is usuallypollinated by social wasps and displays the morphological characteristics of an outcrossing species. In warm,temperate areas of Japan, E. helleborine often appears in alpine or subalpine regions, and has never been foundin low-altitude forests, except for coastal pine forests. The coastal population of E. helleborine is often classified atthe variety level, as E. helleborine var. sayekiana, and the inland populations are known as E. helleborine var.papillosa. It is possible that E. helleborine var. sayekiana possesses a distinctive selfing strategy, as its autonomousself-pollination has evolved in dry habitats, such as coastal dunes. The present study investigated the pollinationbiology of E. helleborine var. sayekiana and var. papillosa to detect differences in their reproductive systems.Unexpectedly, both E. helleborine var. papillosa and E. helleborine var. sayekiana were found to possess aself-pollination strategy and were therefore rarely visited by insects. Self-pollination occurs at the end of theflowering season and probably acts to ensure pollination even if insect-mediated pollination fails. Moreover, thereare no floral differences between E. helleborine var. papillosa and E. helleborine var. sayekiana. These observationssuggest that it is not necessary to distinguish between these two varieties. Furthermore, E. helleborine var.papillosa was pollinated by hoverflies, suggesting that E. helleborine var. papillosa is probably biologically distinctfrom its mother species, E. helleborine. © 2013 The Linnean Society of London, Botanical Journal of the LinneanSociety, 2013, 173, 733–743.
ADDITIONAL KEYWORDS: autogamy – gynostemium morphology – pollination biology – reproductiveassurance – reproductive biology.
INTRODUCTION
It has been noted that some taxa may be accordedprotection unjustified by the biological reality (Pillon& Chase, 2007). This occurs more frequently forendangered species in taxonomically complicatedgroups (Pilgrim, Crawley & Dolphin, 2004; Ennoset al., 2012). One example is the genus Epipactis Zinn(Orchidaceae), with 25 (Richards, 1982), 36 (Delforge,1995) or up to 56 (Delforge, 2001) species. Breedingsystems, which include anatomical, morphologicaland biological aspects of the generative reproductionof individuals and populations, are an importantcharacteristic used for the identification of such prob-lematic taxa (Pedersen & Ehlers, 2000; Squirrell
et al., 2002; Bonatti, Sgarbi & Del Prete, 2006; Tałałaj& Brzosko, 2008; Jakubska-Busse et al., 2012).
Incomplete understanding of the breeding system ofEpipactis is one of the main reasons for the taxonomicdifficulties in this group (Tałałaj & Brzosko, 2008).This phenomenon is exemplified by the Epipactishelleborine (L.) Crantz complex, which has radiatedextensively throughout Eurasia and North Africa andis a widely established alien in North America. It isone of the most widespread orchid species (Luer,1975; Delforge, 1995). This species is the progenitor ofmany problematic allogamous and autogamous sub-groups distinguished by early systematic monographs(Keller & Schlechter, 1940), which are still beingmodified today (Harris & Abbott, 1997; Squirrellet al., 2002). Despite the consensus that allogamy ispredominant, the details of the breeding system for*Corresponding author. E-mail: [email protected]
bs_bs_banner
Botanical Journal of the Linnean Society, 2013, 173, 733–743. With 5 figures
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 733–743 733

this species are still debated (Ehlers & Pedersen,2000; Pedersen & Ehlers, 2000; Squirrell et al., 2002;Bonatti et al., 2006; Tałałaj & Brzosko, 2008), withsome reports asserting that E. helleborine subsp.neerlandica (Verm.) Buttler s.l. exhibits autonomousself-pollination (hereafter self-pollination; Claessens,Kleynen & Wielinga, 1998; Pedersen & Ehlers, 2000).
Epipactis helleborine occurs in a broad range ofhabitats, such as dense forest floors, urban areas,open grasslands with scattered trees and calcareoussoils ranging from the temperate to boreal zones(Salmia, 1986; Buttler, 1991; Delforge, 1995;Hollingsworth & Dickson, 1997). Japanese E. helle-borine typically appears in cool mountainous areas,although it also occurs occasionally in coastal dunes(Weijer, 1953; Pedersen & Ehlers, 2000). Mainly as aresult of habitat differences, the coastal populationsof E. helleborine are classified at the variety level asE. helleborine var. sayekiana (Makino) Hashimoto,whereas the inland populations are known as E. hel-leborine var. papillosa (Franch. & Sav.) Hashimoto(Hashimoto, 1987).
Ogura-Tsujita & Yukawa (2008) investigated therole of mycorrhizal associations as adaptive mecha-nisms that could enable such habitat diversity. Theirstudy revealed that E. helleborine var. papillosa has astrong preference for several taxa of pezizalean ecto-mycorrhizal fungi (Ogura-Tsujita & Yukawa, 2008).In contrast, Ogura-Tsujita & Yukawa (2008) also dis-covered that E. helleborine var. sayekiana was exclu-sively associated with Wilcoxina spp., ectomycorrhizalfungi connected with pine trees growing in coastaldunes. Considering these facts, the ability to associatewith a wide range of ectomycorrhizal fungi appears tobe one of the mechanisms for E. helleborine to suc-cessfully colonize diverse habitats (Ogura-Tsujita &Yukawa, 2008).
However, this trait can only be adaptive when pol-lination is successful in such environments. It isknown that pollinium removal is significantly lower indrier habitats than in mesic habitats, suggesting thatthe rate of pollinator visitation may be lower in theformer (Pedersen & Ehlers, 2000; Ehlers, Olesen &Ågren, 2002; Squirrell et al., 2002). Facultative andobligate autogamous subspecies and varieties of E. hel-leborine are known to exist in dry habitats, such as thesand dunes along the coastline of the North Sea of theNetherlands and Denmark [i.e. E. helleborine subsp.neerlandica (Vermeulen) Buttler and E. helleborinevar. renzii (Robatsch) J.Claess., Kleynen & Wielinga(Claessens et al., 1998)]. The ability to self-pollinatecould potentially be selected for pollinator-limited con-ditions (Baker, 1955; Wyatt, 1988). Therefore, it hasbeen proposed that selfing E. helleborine evolvedthrough the paedomorphosis of outcrossing flowers inlocalized environments (Claessens et al., 1998). Fur-
thermore, in the open sand dunes of northern Jutland,aerial shoots of the obligate selfing E. helleborine var.renzii often die from desiccation, sometimes evenbefore anthesis (Ehlers & Pedersen, 2000). The harshhabitats of sand dunes could provide the selectionpressure for selfing, both as a result of pollinatorscarcity and because selfing shortens the time spanfrom anthesis to fruit set, thereby increasing thechance of a plant surviving long enough to set fruit(Ehlers & Pedersen, 2000). Therefore, in Japan, it ispossible that E. helleborine in sand dune populations(i.e. E. helleborine var. sayekiana) adopts a self-pollination strategy.
Both E. helleborine var. sayekiana and var. papil-losa have also decreased considerably as a result ofhabitat destruction and excessive collection. In Japan,the status of E. helleborine var. sayekiana has beenlisted as ‘vulnerable’, which means that the risk ofextinction is increasing (Environment Agency ofJapan, 2000). The extinction probability of thisspecies in the next 100 years is estimated to be 90%(Environment Agency of Japan, 2000). Epipactis hel-leborine var. papillosa has also been categorized asendangered in many prefectures, although it is notyet designated as an endangered taxon at thenational level (Environment Agency of Japan, 2000).Because breeding systems in plants often profoundlyaffect the levels and partitioning of genetic structureof populations and genetic diversity, information onpollination biology is also important for conservation(Hamrick & Godt, 1997; Gale, 2007).
I conducted the survey on two Japanese varieties ofE. helleborine to characterize their reproductivesystem. Insect visitors were investigated to determinewhether some floral visitors work as effective pollina-tors. Hand pollination experiments were conducted toexamine whether self-pollination occurs in these vari-eties. The feature that distinguishes the outcrossingand selfing flowers of E. helleborine is the rostellum,which serves as a barrier between pollen and stigmain outcrossing flowers, but is reduced or absent in theselfing variants (Pedersen & Ehlers, 2000; Ehlerset al., 2002; Bonatti et al., 2006; Tałałaj & Brzosko,2008). Therefore, the floral architecture (especiallyrostellum morphology) was examined in order todescribe how it promotes the identified breedingtypes.
MATERIAL AND METHODSSTUDY SPECIES
Epipactis helleborine is a self-compatible, long-livedherb. Its geographical distribution is wide, includingEurope and continuing eastwards to Siberia, Chinaand Japan. Coastal Japanese E. helleborine is often
734 K. SUETSUGU
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 733–743

separated at the variety level as E. helleborine var.sayekiana (Fig. 1A), whereas inland populations arerecognized as E. helleborine var. papillosa (Fig. 1B).However, this distinction is made despite the coastalpopulations not being genetically different from theinland populations at the sequence level for the trnLintron (Ogura-Tsujita & Yukawa, 2008). The coastalpopulations occur in Japanese black pine (Pinus thun-bergii Parl.) forests (Maekawa, 1971; Satomi, 1982).These forests are naturally widespread, but are alsoplanted along coastal sand dunes (Satake, 1989). Inwarm, temperate areas of Japan, E. helleborine oftenappears in alpine or subalpine regions, and has neverbeen found in low-altitude forests, except for coastalpine forests (Ogura-Tsujita & Yukawa, 2008).Although E. helleborine var. papillosa and E. helle-borine var. sayekiana are also reported to be distin-guishable by their sepal coloration (var. papillosa isgreenish and var. sayekiana is pinkish; Makino,1918), both varieties have a wide range of coloration(Fig. 2A–F). Individual flowers of both varietiesremain open for 6–8 days unless the flowers are pol-linated by insects (K. Suetsugu, pers. observ.).
INSECT VISITORS AND NATURAL FRUIT SET
The pollination biology of these two varieties wasinvestigated in central and northern Japan from 2009to 2012 (Table 1). I carried out the field studies forE. helleborine var. sayekiana in mid-June from 2009to 2010, at population S1. The study site was an opencoastal dune forest dominated by the sparse Japaneseblack pine, Pinus thunbergii Parl. Flower visitors
were observed, with a total observation time of > 50 h.Observations were made over a span of 2–6 h, cover-ing daylight hours (07.00–18.00 h). I also carried outfield studies for E. helleborine var. papillosa duringmid- to late July in 2009–2012, at population P1. Thestudy site was a coniferous plantation dominated byPicea glehnii Mast. Insect visitor observations weremade over a 4–6-h period, covering daylight hours(07.00–18.00 h), with a total observation time of> 70 h. In addition, the pollination biology of E. hel-leborine var. papillosa was investigated in populationP2 during mid-July in 2011. This study site was aconiferous forest dominated by larch, Larix kaempferi(Lamb.) Carrière. Insect visitor observations weremade over a 3–6-h period, covering daylight hours(07.00–18.00 h), with a total observation time of> 15 h. The natural fruit set ratio was also recorded inpopulations P3 and P4 of E. helleborine var. papillosain 2010 and 2012, respectively.
POLLINATION EXPERIMENT
Pollination experiments were conducted to determinethe ability for and extent of self-fertilization in bothE. helleborine var. sayekiana (population S1) in 2009and E. helleborine var. papillosa (population P1) in2009 and 2011. Twenty inflorescences at the samephenological stage (just before anthesis) were used forthese experiments. Hand pollination experimentswere performed to determine whether fruit set waslimited by the receipt of pollen. Each flower in theexperiment was assigned to one of four treatments.(1) Self-pollination treatment: flowers were bagged
Figure 1. Epipactis helleborine inflorescences: A, flowering E. helleborine var. sayekiana; B, flowering E. helleborine var.papillosa; C, fruiting E. helleborine var. papillosa.
AUTOGAMY IN EPIPACTIS HELLEBORINE 735
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 733–743

Figure 2. Flowers and insect visitors of two varieties: A–C, Epipactis helleborine var. sayekiana; D–F, Epipactishelleborine var. papillosa; rostella are indicated by arrows; G, Chyliza vittata visiting E. helleborine var. papillosa; H, I,Episyrphus balteatus visiting E. helleborine var. papillosa, with viscidium indicated by arrow; J, K, leaf mining ofE. helleborine var. papillosa by C. vittata, eggs and larva, respectively, indicated by arrows.
736 K. SUETSUGU
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 733–743

with a fine-meshed net before anthesis to excludepollinators (N = 5). This treatment was used to testwhether fruit set could occur by self-pollination. (2)Agamospermy treatment: the entire pollinaria wereremoved before anthesis using forceps, and theflowers were then bagged (N = 5). This treatment wasused to test for agamospermy. (3) Artificial self-pollination treatment: the pollinaria were removedand used to hand pollinate the same flower beforebeing bagged (N = 5). (4) Artificial cross-pollinationtreatment: the same as treatment (3), but using pol-linia from a different plant (N = 5). To avoid samplingfrom a genetically identical plant, all plants werekept at least 1 m from their nearest neighbour duringthe cross-pollination experiments. In addition to thefour treatment regimens, flowering individuals wererandomly tagged and allowed to develop fruit bynatural pollination conditions. Differences betweentreatments were analysed by Fisher’s exact test.
SELF-POLLINATION MECHANISM
Well-developed rostellum structure is considered to bethe most important physical barrier between the maleand female parts of the flower, preventing self-fertilization (Pedersen & Ehlers, 2000; Ehlers et al.,2002; Bonatti et al., 2006; Tałałaj & Brzosko, 2008).In most self-pollinated orchids, this structure does notdevelop, develops incompletely or sometimes disinte-grates during flowering (Catling, 1990). Flowers werethus examined in terms of whether their architecturemight promote or protect against spontaneous auto-gamy. In both varieties, randomly chosen inflores-cences at different developmental stages (at c. 1 day,3 days, 1 week and 2 weeks after anthesis) werechecked for the presence of the rostellum and thecontact between stigma and pollinia. At least 20flowers from five individuals at each developmentalstage were observed in every population of bothvarieties.
Because bagging experiments and column morphol-ogy investigations suggested that self-pollinationoccurred, I then observed pollen tube elongation usinganiline blue staining to confirm the self-pollination
strategy. Each of the ten columns with nylon-meshedbagging of populations S1, P1 and P2 at c. 1 day,3 days, 1 week and 2 weeks after flower opening wereexcised and fixed in 3 : 1 ethanol : acetic acid for30 min and then softened in 1 M KOH for 1 h at 60 °Cusing a block heater. The columns were washed indistilled water for a few minutes and stained with0.1% aniline blue in K3PO4 buffer, pH 8.5, for 2 h atroom temperature. The samples were rinsed brieflyin distilled water and mounted in 50% glycerol.The samples were squashed and visualized by UVmicroscopy.
RESULTSINSECT VISITORS
The frequency of flower visits by any insect wasextremely low in both varieties. No insect visitorscarrying pollinia were observed throughout theintensive monitoring period in E. helleborine var.sayekiana. The fly Japanagromyza tokunagai (Agro-myzidae), the larvae of which feed on orchid seeds,occasionally visited the plants to lay eggs on theyoung ovaries. However, they never went inside theinner portion of the flower or touched the gynoste-mium. In population P1, E. helleborine var. papillosawas visited by Chyliza vittata (Psilidae), which laideggs on flowers and leaves (Fig. 2G). They have beenrecorded as the leaf, stem and root miner of diverseorchids, including E. helleborine (Robbins, 1991), andthus can be considered as an antagonist (Fig. 2J, K).Although they occasionally went inside the innerportion of the flower, there was no pollinium removal/deposit observed during their visitations. Four indi-viduals of the syrphid fly Episyrphus balteatus visitedthe flowers and landed on the labellum (Fig. 2H, I).Among them, two were observed to have removedpollinia, which were attached to the head during thevisitation. In population P2, E. helleborine var. pap-illosa was also visited by C. vittata, which laid eggson the flowers (Fig. 2G). However, they never wentinside the inner portion of the flower or touched thegynostemium.
Table 1. Study populations of two varieties of Epipactis helleborine. Population size is given as number of floweringplants
Plant species Population Site Habitat type Population size
var. sayekiana S1 Fujisawa, Kanagawa Coastal dunes forest c. 200var. papillosa P1 Ebetsu, Hokkaido Coniferous plantation c. 70var. papillosa P2 Koumi, Nagano Natural coniferous forest 18var. papillosa P3 Chino, Nagano Natural coniferous forest 23var. papillosa P4 Sappro, Hokkaido Coniferous plantation 15
AUTOGAMY IN EPIPACTIS HELLEBORINE 737
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 733–743
POLLINATION EXPERIMENT
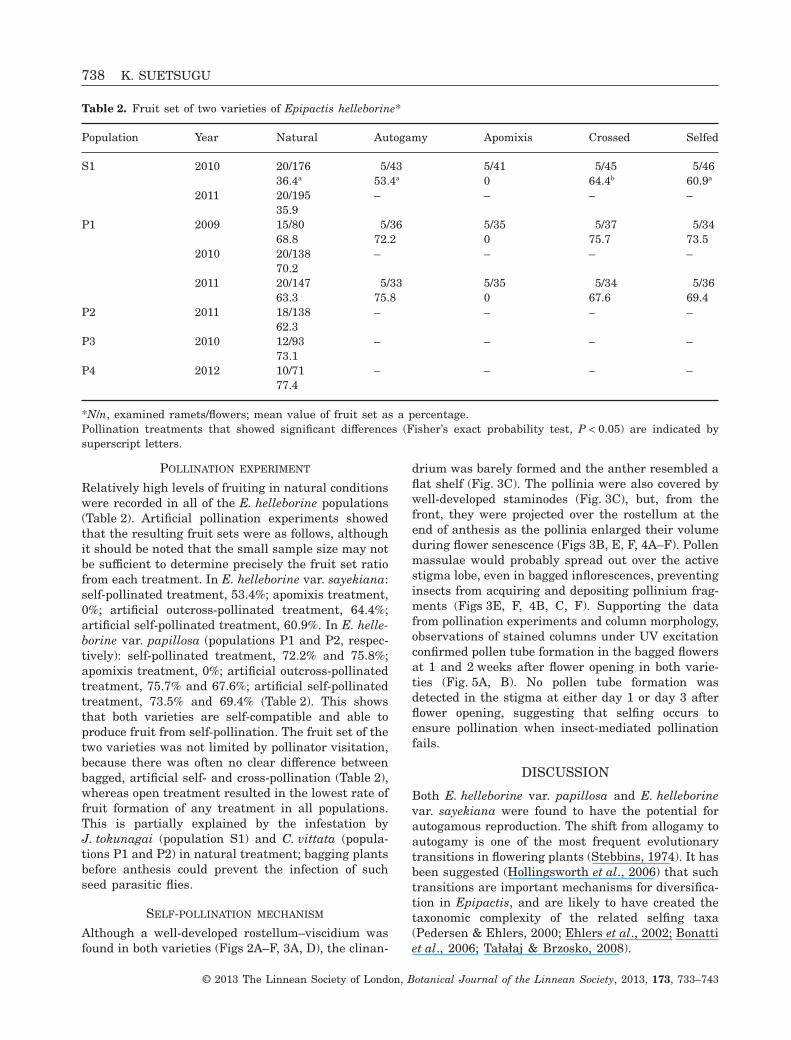
Relatively high levels of fruiting in natural conditionswere recorded in all of the E. helleborine populations(Table 2). Artificial pollination experiments showedthat the resulting fruit sets were as follows, althoughit should be noted that the small sample size may notbe sufficient to determine precisely the fruit set ratiofrom each treatment. In E. helleborine var. sayekiana:self-pollinated treatment, 53.4%; apomixis treatment,0%; artificial outcross-pollinated treatment, 64.4%;artificial self-pollinated treatment, 60.9%. In E. helle-borine var. papillosa (populations P1 and P2, respec-tively): self-pollinated treatment, 72.2% and 75.8%;apomixis treatment, 0%; artificial outcross-pollinatedtreatment, 75.7% and 67.6%; artificial self-pollinatedtreatment, 73.5% and 69.4% (Table 2). This showsthat both varieties are self-compatible and able toproduce fruit from self-pollination. The fruit set of thetwo varieties was not limited by pollinator visitation,because there was often no clear difference betweenbagged, artificial self- and cross-pollination (Table 2),whereas open treatment resulted in the lowest rate offruit formation of any treatment in all populations.This is partially explained by the infestation byJ. tokunagai (population S1) and C. vittata (popula-tions P1 and P2) in natural treatment; bagging plantsbefore anthesis could prevent the infection of suchseed parasitic flies.
SELF-POLLINATION MECHANISM
Although a well-developed rostellum–viscidium wasfound in both varieties (Figs 2A–F, 3A, D), the clinan-
drium was barely formed and the anther resembled aflat shelf (Fig. 3C). The pollinia were also covered bywell-developed staminodes (Fig. 3C), but, from thefront, they were projected over the rostellum at theend of anthesis as the pollinia enlarged their volumeduring flower senescence (Figs 3B, E, F, 4A–F). Pollenmassulae would probably spread out over the activestigma lobe, even in bagged inflorescences, preventinginsects from acquiring and depositing pollinium frag-ments (Figs 3E, F, 4B, C, F). Supporting the datafrom pollination experiments and column morphology,observations of stained columns under UV excitationconfirmed pollen tube formation in the bagged flowersat 1 and 2 weeks after flower opening in both varie-ties (Fig. 5A, B). No pollen tube formation wasdetected in the stigma at either day 1 or day 3 afterflower opening, suggesting that selfing occurs toensure pollination when insect-mediated pollinationfails.
DISCUSSION
Both E. helleborine var. papillosa and E. helleborinevar. sayekiana were found to have the potential forautogamous reproduction. The shift from allogamy toautogamy is one of the most frequent evolutionarytransitions in flowering plants (Stebbins, 1974). It hasbeen suggested (Hollingsworth et al., 2006) that suchtransitions are important mechanisms for diversifica-tion in Epipactis, and are likely to have created thetaxonomic complexity of the related selfing taxa(Pedersen & Ehlers, 2000; Ehlers et al., 2002; Bonattiet al., 2006; Tałałaj & Brzosko, 2008).
Table 2. Fruit set of two varieties of Epipactis helleborine*
Population Year Natural Autogamy Apomixis Crossed Selfed
S1 2010 20/176 5/43 5/41 5/45 5/4636.4a 53.4a 0 64.4b 60.9a
2011 20/195 – – – –35.9
P1 2009 15/80 5/36 5/35 5/37 5/3468.8 72.2 0 75.7 73.5
2010 20/138 – – – –70.2
2011 20/147 5/33 5/35 5/34 5/3663.3 75.8 0 67.6 69.4
P2 2011 18/138 – – – –62.3
P3 2010 12/93 – – – –73.1
P4 2012 10/71 – – – –77.4
*N/n, examined ramets/flowers; mean value of fruit set as a percentage.Pollination treatments that showed significant differences (Fisher’s exact probability test, P < 0.05) are indicated bysuperscript letters.
738 K. SUETSUGU
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 733–743

Figure 3. Column morphology and autogamy in Epipactis helleborine var. sayekiana. Side view: A, 24 h after anthesis;B, 1 week after anthesis. C, Dorsal view showing clinandrium and rostellum. Lower view: D, 24 h after anthesis; E,1 week after anthesis; F, 2 weeks after anthesis. CC, clinandrium cavity (a cavity behind the stigma, surrounding thebasal part of the fertile anther); RS, rostellum; ST, staminodes (wing-like remnants of two anthers, which have lost theiroriginal function to produce pollen); VR, rostellum–viscidium (morphological modification of the median stigmatic lobesecreting a sticky substance usually responsible for sticking the pollinium to the body of the pollinator and for limitingthe possibility of self-pollination).
AUTOGAMY IN EPIPACTIS HELLEBORINE 739
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 733–743

The well-developed rostellum of E. helleborineusually serves as a barrier separating the male andfemale parts of the flower, and the viscidium exudesfrom the tip of the rostellum (Richards, 1982) andacts as a glue to bind the pollinia to visiting insects.
However, most taxonomic problems in Epipactiscentre on the large number of named taxa in whichthe viscidium and rostellum are highly reduced orless persistent (Claessens et al., 1998; Pedersen &Ehlers, 2000; Ehlers et al., 2002; Bonatti et al., 2006;
Figure 4. Column morphology and autogamy in Epipactis helleborine var. papillosa. All photographs represent theflowers from 2 weeks after anthesis. A–C, Front view. D, Lateral view. E, Upper view. F, Lower view. Rostellum indicatedby arrows. Sepal and petal were removed from E and F.
740 K. SUETSUGU
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 733–743

Tałałaj & Brzosko, 2008). Any reduction of the ros-tellum diminishes the physical barrier betweenthe anthers and the stigma, whereas reduction ofthe viscidium lowers the likelihood of the polliniaattaching to insect visitors (Claessens et al., 1998;Pedersen & Ehlers, 2000; Ehlers et al., 2002);instead, the pollinia remain in situ and fall down-wards directly onto the stigmatic surface, resulting inself-pollination.
However, the current study found that both E. hel-leborine var. papillosa and E. helleborine var. sayeki-ana possess a well-developed rostellum (Figs 1A–F,3A, D), although it did not seem to be a significantbarrier to self-fertilization. Instead, self-pollination inthese varieties seemed to be a consequence of a poorlydeveloped clinandrium, which resulted in the pollengrains being pushed forward, especially during flowersenescence (Figs 3B, E, F, 4A–F) when the polliniabecome enlarged and less compact. A similar systemof self-fertilization has also been reported forE. palustris (L.) Crantz (Godfery, 1933; Tałałaj &Brzosko, 2008). In this case, pollen is prevented fromfalling straight onto the stigma zone, which is locatedat the back of the projecting rostellum, with theviscidium and pollinia. This positioning results in thepollen tetrads falling near the rostellum/viscidium.During wilting, the viscidium dries and the rostellum/
viscidium barrier becomes weaker, allowing thepollen tetrads, which are no longer trapped in thesticky glue, to be transferred to the upper edge ofthe stigma (Tałałaj & Brzosko, 2008). Furthermore, itis likely that exudates from the stigma promote thespread of pollen grains over the stigma lobe. In thisstudy, observations of the stained columns under UVexcitation confirmed that pollen falls from the fragilepollinia onto the upper edges of the stigma at the endof anthesis in both E. helleborine var. papillosa andE. helleborine var. sayekiana (Fig. 5A, B). However,although both varieties had the potential for self-pollination, their average fruit set was much lowerthan that of obligate autogamous taxa, which producealmost 100% fruit set (Tremblay et al., 2005). Thissuggests that both varieties employ a facultativeautogamous strategy. Indeed, both varieties can beinsect pollinated, with self-pollination occurring pre-dominantly at the end of the flowering season(Figs 3B, E, F, 4A–F). The evolution of functioningautogamy in Japanese populations of E. helleborinecan therefore still be considered to be in its earlystages (H. Æ. Pedersen, pers. comm.).
It has been suggested that self-pollination in flow-ering plants is an adaptation that ensures reproduc-tion when pollinators are scarce (Lloyd, 1992;Vaughton, Ramsey & Simpson, 2008; Zhang & Li,
Figure 5. Epifluorescence images of two varieties of Epipactis helleborine column from 1 week after anthesis, stainedwith aniline blue: A, var. sayekiana; B, var. papillosa. Po, pollen grains appressed to the stigma; Pt, pollen tubes emergingfrom germinating pollen grains. Scale bars, 100 μm.
AUTOGAMY IN EPIPACTIS HELLEBORINE 741
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 733–743

2008). It has been noted that the removal of polliniafrom flowers of E. helleborine in drier habitats issignificantly lower than in mesic habitats, which sug-gests that the rate of pollinator visits is also lower(Ehlers et al., 2002). Furthermore, populations ofE. helleborine in mesic deciduous and coniferousforests are often outcrossing, whereas those in drierhabitats are predominantly self-fertilizing varietiesand subspecies (Robatsch, 1988; Claessens et al.,1998; Pedersen & Ehlers, 2000). It was thereforesurprising that the current study found that theinland var. papillosa and the coastal var. sayekianaboth exhibited a selfing strategy. However, observa-tion of the plants in their natural environmentrevealed that there were few insect visitors, suggest-ing that it is not only the environmental conditionsper se that promote self-pollination in these varieties,but also the scarcity of pollinators.
Epipactis helleborine is mainly pollinated by socialwasps, including Vespula vulgaris and V. germanica,and is a prime example of a wasp-pollinated flower(Judd, 1971, 1979; Jakubska et al., 2005). Further-more, despite its large nectar reward, this species isknown to be almost entirely overlooked by otherpotential pollinators (Darwin, 1862; Brodmann et al.,2008). Brodmann et al. (2008) revealed that the floralscent of E. helleborine plays a key role in the attrac-tion of prey-hunting social wasps by mimicking theso-called green leaf volatiles produced by plants wheninfested with herbivorous insects. Although both vari-eties failed to attract wasp pollinators, the inlandvariety E. helleborine var. papillosa was pollinated byhoverflies (Fig. 2H–I). The pollination rate was verylow, but this may provide evidence that E. helleborinevar. papillosa can produce different attractants tolure specific pollinators.
We could not detect any floral differences betweenthese two varieties. In addition, Ogura-Tsujita &Yukawa (2008) demonstrated that E. helleborine var.sayekiana does not differ genetically from E. helle-borine var. papillosa at the level of sequences of thetrnL intron. Furthermore, their range of coloration iscontinuous and the differentiation is vague (Fig. 2A–F), although both varieties are reportedly differenti-ated by sepal coloration (Makino, 1918). Theseobservations suggest that it may not be necessary todistinguish these two varieties. In contrast, the factthat E. helleborine var. sayekiana was pollinated byhoverflies suggests that E. helleborine var. sayekianais biologically distinct from the progenitor E. helle-borine because they utilize different pollinators. Moreintensive study of these two varieties, including pol-linator observations, analyses of chemical attractantsfor insect visitors and genetic analysis, may be nec-essary to understand the biological and taxonomicalcomplexity of these two varieties.
ACKNOWLEDGEMENTS
The author thanks Michiya Murakawa, Kiyoshi Kawa-guchi, Katsuro Hasegawa and Hideo Shirane for fieldassistance and/or habitat information. The author alsothanks Dr Henrik Æ. Pedersen and the anonymousreviewer for their constructive comments on the manu-script. This research was partially supported by aJapan Society for the Promotion of Science ResearchFellowships for Young Scientists Grant.
REFERENCES
Baker HG. 1955. Self-compatibility and establishment after‘long-distance’ dispersal. Evolution 9: 347–348.
Bonatti PM, Sgarbi E, Del Prete C. 2006. Gynostemiummicromorphology and pollination in Epipactis microphylla(Orchidaceae). Journal of Plant Research 119: 431–437.
Brodmann J, Twele R, Francke W, Hölzler G, Zhang Q,Ayasse M. 2008. Orchids mimic green-leaf volatiles toattract prey-hunting wasps for pollination. Current Biology18: 740–744.
Buttler KP. 1991. Field guide to orchids of Britain andEurope. Marlborough: Crowood Press.
Catling PM. 1990. Auto-pollination in the Orchidaceae.Orchid Biology: Reviews and Perspectives 5: 121–158.
Claessens J, Kleynen J, Wielinga R. 1998. Some notes onEpipactis helleborine (L.) Crantz ssp. neerlandica (Vermeu-len) Buttler and Epipactis renzii K. Robatsch. Eurorchis 10:55–64.
Darwin C. 1862. The various contrivances by which orchidsare fertilised by insects. London: John Murray.
Delforge P. 1995. Orchids of Britain and Europe. London:Harper Collins.
Delforge P. 2001. Guide des Orchidées d’Europe, d’Afrique duNord et du Proche-Orient. Lausanne: Delachaux et Niestlé.
Ehlers BK, Olesen JM, Ågren J. 2002. Floral morphologyand reproductive success in the orchid Epipactis helleborine:regional and local across-habitat variation. Plant Systemat-ics and Evolution 236: 19–32.
Ehlers BK, Pedersen HA. 2000. Genetic variation in threespecies of Epipactis (Orchidaceae): geographic scale andevolutionary inferences. Biological Journal of the LinneanSociety 69: 411–430.
Ennos RA, Whitlock R, Fay MF, Jones B, Neaves LE,Payne R, Taylor I, de Vere N, Hollingsworth PM. 2012.Process-based Species Action Plans: an approach to conservecontemporary evolutionary processes that sustain diversityin taxonomically complex groups. Botanical Journal of theLinnean Society 168: 194–203.
Environment Agency of Japan. 2000. Threatened wildlifeof Japan: red data book, 2nd edn. Tokyo: Japan WildlifeResearch Center [in Japanese].
Gale S. 2007. Autogamous seed set in a critically endangeredorchid in Japan: pollination studies for the conservation ofNervilia nipponica. Plant Systematics and Evolution 268:59–73.
742 K. SUETSUGU
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 733–743

Godfery MJ. 1933. Monograph and iconograph of nativeBritish Orchidaceae. Cambridge: Cambridge UniversityPress.
Hamrick JL, Godt MJW. 1997. Conservation genetics. In:Avise JC, Hamrick JL, eds. Conservation genetics. NewYork: Chapman Hall, 281–304.
Harris SA, Abbott RJ. 1997. Isozyme analysis of thereported origin of a new hybrid, orchid species, Epipactisyoungiana (Young’s helleborine), in the British Isles. Hered-ity 79: 402–407.
Hashimoto T. 1987. Our recent knowledge of the Japaneseorchid flora. In: Proceedings of the 12th World Orchid Con-ference. Tokyo: Nippon Tosyo Tsushin Hanbai, 118–126.
Hollingsworth PM, Dickson JH. 1997. Genetic variation inrural and urban populations of Epipactis helleborine (L.)Crantz. (Orchidaceae) in Britain. Botanical Journal of theLinnean Society 123: 321–331.
Hollingsworth PM, Squirrell J, Hollingsworth ML,Richards AJ, Bateman RM. 2006. Taxonomic complexity,conservation and recurrent origins of self-pollination inEpipactis (Orchidaceae). In: Bailey JP, Ellis RG, eds.Current taxonomic research on the British and Europeanflora. London: Botanical Society of the British Isles, 27–44.
Jakubska A, Przado D, Steininger M, Aniolł-Kwiatkowska J, Kadej M. 2005. Why do pollinatorsbecome ‘sluggish’? Nectar chemical constituents from Epi-pactis helleborine (L.) Crantz (Orchidaceae). AppliedEcology and Environmental Research 3: 29–38.
Jakubska-Busse A, Procków J, Górniak M, Gola EM.2012. Is Epipactis pseudopurpurata distinct from E. purpu-rata (Orchidaceae)? Evidence from morphology, anatomy,DNA and pollination biology. Botanical Journal of theLinnean Society 170: 243–256.
Judd WW. 1971. Wasps (Vespidae) pollinating helleborine,Epipactis helleborine (L.) Crantz, at Owen Sound, Ontario.Proceedings of the Entomological Society of Ontario 102:115–118.
Judd WW. 1979. Arthropods associated with helleborineorchid, Epipactis helleborine (L.) Crantz., at Dunnville,Ontario. Entomological News 90: 41–44.
Keller G, Schlechter R. 1940. Monographie und Iconogra-phie der Orchideen Europas und des Mittelmeergebietes.Berlin: Dahlem bei, 329–350.
Lloyd DG. 1992. Self- and cross-fertilization in plants. II. Theselection of self-fertilization. International Journal of PlantSciences 153: 370–380.
Luer CA. 1975. The native orchids of the United States andCanada, excluding Florida. New York: New York BotanicalGarden.
Maekawa F. 1971. The wild orchids of Japan in color. Tokyo:Seibundo.
Makino T. 1918. New species, Epipactis sayekiana Makino.Journal of Japanese Botany 2: 22.
Ogura-Tsujita Y, Yukawa T. 2008. Epipactis helleborineshows strong mycorrhizal preference towards ectomycorrhi-zal fungi with contrasting geographic distributions inJapan. Mycorrhiza 18: 331–338.
Pedersen HAE, Ehlers BK. 2000. Local evolution of obligateautogamy in Epipactis helleborine subsp. neerlandica (Orchi-daceae). Plant Systematics and Evolution 223: 173–183.
Pilgrim ES, Crawley MJ, Dolphin K. 2004. Patterns ofrarity in the native British flora. Biological Conservation120: 165–174.
Pillon Y, Chase MW. 2007. Taxonomic exaggeration and itseffects on orchid conservation: research note. ConservationBiology 21: 263–265.
Richards AJ. 1982. The influence of minor structuralchanges in the flower on breeding systems and speciation inEpipactis Zinn (Orchidaceae). In: Armstrong JA, Powell JM,Richards AJ, eds. Pollination and Evolution. Sydney: RoyalBotanic Gardens, 47–53.
Robatsch K. 1988. Beiträge zur Kenntnis europäischerEpipactis-Arten. Linzer Biologische Beiträge 20: 161–172.
Robbins J. 1991. Leaf-mining insects in Warwickshire: part6. Further additions and revisions. Proceedings of the Bir-mingham Natural History Society 26: 84–87.
Salmia A. 1986. Chlorophyll-free form of Epipactis helle-borine (Orchidaceae) in SE Finland. Annales BotaniciFennici 23: 49–57.
Satake Y. 1989. Pinaceae. Wild flowers of Japan: woodyplants. Tokyo: Heibonsha.
Satomi N. 1982. Orchidaceae. In: Satake Y, Ohwi J,Kitamura S, Watari S, Tominari T, eds. Wild flowers ofJapan: herbaceous plants. Monocotyledoneae. Tokyo: Heibo-nsha, 187–235.
Squirrell J, Hollingsworth PM, Bateman RM, TebbittMC, Hollingsworth ML. 2002. Taxonomic complexity andbreeding system transitions: conservation genetics of theEpipactis leptochila complex (Orchidaceae). MolecularEcology 11: 1957–1964.
Stebbins GL. 1974. Flowering plants: evolution above thespecies level. Cambridge: The Belknap Press of HarvardUniversity Press.
Suetsugu K. 2013. Autogamous fruit set in a mycoheter-otrophic orchid Cyrtosia septentrionalis. Plant Systematicsand Evolution 299: 481–486.
Tałałaj I, Brzosko E. 2008. Selfing potential in Epipactispalustris, E. helleborine and E. atrorubens (Orchidaceae).Plant Systematics and Evolution 276: 21–29.
Tremblay RL, Ackerman JD, Zimmerman JK, Calvo RN.2005. Variation in sexual reproduction in orchids and itsevolutionary consequences: a spasmodic journey to diversi-fication. Biological Journal of the Linnean Society 84: 1–54.
Vaughton G, Ramsey M, Simpson I. 2008. Does selfingprovide reproductive assurance in the perennial herbBulbine vagans (Asphodelaceae)? Oikos 117: 390–398.
Weijer J. 1953. The colour-differences in Epipactis helle-borine (L.) Cr. Wats. & Coult., and the selection of thegenetical varieties by environment. Genetica 26: 1–32.
Wyatt R. 1988. Phylogenetic aspects of the evolution of self-pollination. In: Gottlieb LD, Jain SK, eds. Plant evolution-ary biology. London: Chapman and Hall, 109–131.
Zhang ZQ, Li QJ. 2008. Autonomous selfing provides repro-ductive assurance in an alpine ginger Roscoea schneideri-ana (Zingiberaceae). Annals of Botany 102: 531–538.
AUTOGAMY IN EPIPACTIS HELLEBORINE 743
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 733–743




















