
Death and disappearance: estimating mortality risks associated with philopatry and dispersal
7
Death and disappearance: estimating mortality risks associated with philopatry and dispersal Models dealing with the relationship of inbreeding to dispersal, the evolution of cooperative breeding, and the adaptive significance of reproductive suppression all involve a parameter describing die probability that emigration ends in death. Surprisingly, estimates of this parameter (or equivalently, of the probability of dispersing successfully) are virtually absent from the literature. There are formidable difficulties in estimating dispersal risk by direct observation of emigrants, but we contend that many investigators have already collected demographic information that can be used to estimate it indirectly. The root problem is that the proportion of the population diat emigrates from its natal group or home range, e, is not generally observable. Nevertheless, for each value of e diere is only one possible survival rate of emigrants (s t ) and of philopatric animals (s f ) diat can explain the overall survival rate. For study sites Uiat are neither sources nor sinks, we describe a graphical framework within which census data and behavioral observations can be used to place bounds on e, s,, and s p . We use data from a Serengeti population of the dwarf mongoose (HelogaU parvula) to illustrate how different types of data and assumptions allow us to construct better estimates of these parameters. For dwarf mongooses, die approach reveals unexpected differences in dispersal risk among sex and age classes. Key words: cooperative breeding, dispersal, dispersal cost, dispersal risk, emigration, HelogaU, Herpestidae, immigration, inbreeding avoidance, mongoose, mortality, Serengeti. [Behav Ecol 5:135-141 (1994)] Peter M. Waser Scott R. Creel Jeffrey R. Lucas Department of Biological Sciences, Purdue University, West Lafayette, IN 47907, USA ' I 'he probability diat an emigrant will survive to A. immigrate into a new home range or social group is a critical parameter in die evolution of dispersal and related phenomena. Is emigration a voluntary act by a young animal, or is it forced by parents? Anderson (1989) argued that an emigrat- ing rodent's presumed low probability of survival favors the latter interpretation. Is dispersal driven by the costs of inbreeding? Bengtsson (1978) be- lieved that mortality due to emigration would rarely outweigh the fitness costs of close inbreeding, so diat selection would generally promote emigration. What ecological conditions set the stage for co- operative breeding? Brown (1974) promoted the idea that habitat saturation, in effect an ecological circumstance diat decreases die probability of suc- cessful dispersal, can favor delayed emigration and set die stage for "helping" in groups of relatives. How much should dominant individuals suppress die reproductive attempts of subordinates? Veh- rencamp (1983) devised a model that predicts die answer in terms of (among odier diings) die mor- tality risk associated widi dispersal. These and many odier questions remain topics of strenuous debate in part because we still lack estimates of dispersal risk. A recent and extensive review of dispersal in small mammals (Anderson, 1989) summarized many data suggesting diat pop- ulations are relatively impervious to immigrants, but tiiere were no explicit estimates of die proba- bility diat an emigrant successfully immigrates. No measures of dispersal risk appear in a recent com- pendium of case studies in mammalian dispersal (Chepko-Sade and Halpin, 1987). An earlier survey of correlates of philopatry in solitary mammals (Waser and Jones, 1983) uncovered only two data sets from which such estimates could be extracted, and a survey by Johnson and Gaines (1990) found only diree more. Brown (1987) explicidy addressed die possible involvement of risky dispersal in die evolution of avian cooperative breeding, but re- ported no estimates of dispersal risk. At first glance, die best way to estimate the prob- ability that a disperser survives would seem to be through direct observation of emigrating individ- uals. However, there are formidable logistic diffi- culties in tracking individuals continuously from the act of emigration until they either die or suc- ceed in settling in a new home range or group. Few studies have succeeded in radio tracking more than a handful of dispersers to settlement or death (Boyce and Boyce, 1988; Garrett and Franklin, 1988; Holekamp, 1984; Jones, 1982; Leuze, 1980; van Vuren and Armitage, in press; Wiggett and Boag, 1989; see also Pollock et al., 1989, for methods to estimate survival rates of radio tracked dispersers). Radiotclemetry may bias mortality rates; even if it docs not, for many species die rate or scale of dispersal movements is simply too great to make continuous tracking feasible. An alternative is to estimate die proportion of emigrants diat survive from census data, widiout following dispersing animals. If we could accurately estimate die number of animals, E, diat emigrate from their groups or territories, and die number, /, diat successfully immigrate into or establish new groups (territories), die ratio 1/E would provide a straightforward approximation of s,, an emigrant's chance of surviving dispersal (Table 1). S. R. Creel is also affiliated with the Wildlife Conservation Re- search Unit, Department of Zo- ology, University of Oxford, South Parks Road, Oxford OX1 3PS, UK, and the African Wild Dog Project, Selous Conserva- tion Programme, Dar es Sa- laam, Tanzania, East Africa Received 5 August 1992 Revised 10 February 1993 Accepted 26 February 1993 1045-2249/94/$5.00 © 1994 International Society for Behavioral Ecology Waser et al. • Dispersal risk 135 at Montana State University Library on September 8, 2015 http://beheco.oxfordjournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Death and disappearance: estimating mortality risks associated with philopatry and dispersal

Death and disappearance: estimatingmortality risks associated withphilopatry and dispersal
Models dealing with the relationship of inbreeding to dispersal, the evolution of cooperative breeding,and the adaptive significance of reproductive suppression all involve a parameter describing die probabilitythat emigration ends in death. Surprisingly, estimates of this parameter (or equivalently, of the probabilityof dispersing successfully) are virtually absent from the literature. There are formidable difficulties inestimating dispersal risk by direct observation of emigrants, but we contend that many investigators havealready collected demographic information that can be used to estimate it indirectly. The root problemis that the proportion of the population diat emigrates from its natal group or home range, e, is notgenerally observable. Nevertheless, for each value of e diere is only one possible survival rate of emigrants(st) and of philopatric animals (sf) diat can explain the overall survival rate. For study sites Uiat are neithersources nor sinks, we describe a graphical framework within which census data and behavioral observationscan be used to place bounds on e, s,, and sp. We use data from a Serengeti population of the dwarfmongoose (HelogaU parvula) to illustrate how different types of data and assumptions allow us to constructbetter estimates of these parameters. For dwarf mongooses, die approach reveals unexpected differencesin dispersal risk among sex and age classes. Key words: cooperative breeding, dispersal, dispersal cost,dispersal risk, emigration, HelogaU, Herpestidae, immigration, inbreeding avoidance, mongoose, mortality,Serengeti. [Behav Ecol 5:135-141 (1994)]
Peter M. WaserScott R. CreelJeffrey R. LucasDepartment of BiologicalSciences,Purdue University,West Lafayette, IN 47907,USA
' I 'he probability diat an emigrant will survive toA. immigrate into a new home range or social
group is a critical parameter in die evolution ofdispersal and related phenomena. Is emigration avoluntary act by a young animal, or is it forced byparents? Anderson (1989) argued that an emigrat-ing rodent's presumed low probability of survivalfavors the latter interpretation. Is dispersal drivenby the costs of inbreeding? Bengtsson (1978) be-lieved that mortality due to emigration would rarelyoutweigh the fitness costs of close inbreeding, sodiat selection would generally promote emigration.What ecological conditions set the stage for co-operative breeding? Brown (1974) promoted theidea that habitat saturation, in effect an ecologicalcircumstance diat decreases die probability of suc-cessful dispersal, can favor delayed emigration andset die stage for "helping" in groups of relatives.How much should dominant individuals suppressdie reproductive attempts of subordinates? Veh-rencamp (1983) devised a model that predicts dieanswer in terms of (among odier diings) die mor-tality risk associated widi dispersal.
These and many odier questions remain topicsof strenuous debate in part because we still lackestimates of dispersal risk. A recent and extensivereview of dispersal in small mammals (Anderson,1989) summarized many data suggesting diat pop-ulations are relatively impervious to immigrants,but tiiere were no explicit estimates of die proba-bility diat an emigrant successfully immigrates. Nomeasures of dispersal risk appear in a recent com-pendium of case studies in mammalian dispersal(Chepko-Sade and Halpin, 1987). An earlier survey
of correlates of philopatry in solitary mammals(Waser and Jones, 1983) uncovered only two datasets from which such estimates could be extracted,and a survey by Johnson and Gaines (1990) foundonly diree more. Brown (1987) explicidy addresseddie possible involvement of risky dispersal in dieevolution of avian cooperative breeding, but re-ported no estimates of dispersal risk.
At first glance, die best way to estimate the prob-ability that a disperser survives would seem to bethrough direct observation of emigrating individ-uals. However, there are formidable logistic diffi-culties in tracking individuals continuously fromthe act of emigration until they either die or suc-ceed in settling in a new home range or group. Fewstudies have succeeded in radio tracking more thana handful of dispersers to settlement or death (Boyceand Boyce, 1988; Garrett and Franklin, 1988;Holekamp, 1984; Jones, 1982; Leuze, 1980; vanVuren and Armitage, in press; Wiggett and Boag,1989; see also Pollock et al., 1989, for methods toestimate survival rates of radio tracked dispersers).Radiotclemetry may bias mortality rates; even if itdocs not, for many species die rate or scale ofdispersal movements is simply too great to makecontinuous tracking feasible.
An alternative is to estimate die proportion ofemigrants diat survive from census data, widioutfollowing dispersing animals. If we could accuratelyestimate die number of animals, E, diat emigratefrom their groups or territories, and die number,/, diat successfully immigrate into or establish newgroups (territories), die ratio 1/E would provide astraightforward approximation of s,, an emigrant'schance of surviving dispersal (Table 1).
S. R. Creel is also affiliated withthe Wildlife Conservation Re-search Unit, Department of Zo-ology, University of Oxford,South Parks Road, Oxford OX13PS, UK, and the African WildDog Project, Selous Conserva-tion Programme, Dar es Sa-laam, Tanzania, East Africa
Received 5 August 1992Revised 10 February 1993Accepted 26 February 19931045-2249/94/$5.00© 1994 International Societyfor Behavioral Ecology
Waser et al. • Dispersal risk 135
at Montana State U
niversity Library on Septem
ber 8, 2015http://beheco.oxfordjournals.org/
Dow
nloaded from

Table 1Variables referred to in the text
E Number of animals that emigrate from their natalgroup or territory; where behavioral observationsexist, E must equal the number of observedemigrations plus some fraction of the number ofunexplained disappearances
/ Number of emigrants that succeed in immigratinginto a new group or territory, either on or off thestudy area. Can be estimated by counting thenumber of animals in new territories or groupsfrom one census to the next (Ic) or by countingthe number of animals observed to move into anew group or territory (/,)
N Number of animals on the study site during acensus
P Number of animals present on the study siteduring a census at time t that remain in theiroriginal group or territory at time / + 1
D Number of animals present on the study siteduring a census at time t that are no longerpresent in their original group or territory at time( + 1; where behavioral observations exist, D isthe sum of the number of observed emigrations,the number of known deaths in the group, andthe number of unexplained disappearances
e Proportion of the study population that leaves itsnatal group or territory between times t and ( + 1
s. Proportion of emigrants that survive to time f + 1sf Proportion of philopatric animals that survive to
time ( + 1
This approach has not been widely applied be-cause of the well-known problem of unexplaineddisappearances. Even when populations are underclose observation, many individuals disappear un-der unknown circumstances. These individuals mayor may not have emigrated, and if they emigrated,they may have died during emigration or may havesuccessfully immigrated into a group or territoryoff the study site. In this paper, we suggest a partialsolution to this problem.
First, we adopt the common assumption that thestudy area is neither a source nor a sink, so thatthe number of animals that originate off the studysite but successfully settle within it is an estimateof the number of successful off-site dispersers (wecomment in the Discussion on the validity of thisassumption). Making this assumption allows us toestimate / from censuses, even when the populationunder study is not closed, but E remains unknown:some of the remaining unexplained disappearancesrepresent animals that died after emigrating, butothers represent deaths among philopatric individ-uals. Nevertheless, we show that it is generally pos-sible to learn something about these parametersbecause census data tell us the overall survival rate.Our approach is to plot feasible values of the sur-vival rate of emigrants s,, and the survival rate ofphilopatric individuals sp, as functions of the pro-portion of the study population that emigrates, t.Although e, st, and S/, are unknown, census or be-havioral data can be used to set bounds on theirpossible values. The investigator can further nar-row those bounds if the assumption is made thatS/, S s,. In some situations, the investigator mayprefer to use the mortality data from a class of
nondispersing individuals (where sp is known) toestimate e and s,. The consequences of such as-sumptions are easily visualized using our approach.
In this paper, we illustrate the indirect estimationof e, Sp, and s, using data from a long-term studyof dwarf mongooses (HelogaU parvula) and showhow it can be used to detect differences in dispersalrisk among age classes, sexes, or populations. Else-where (Creel and Waser, 1991, submitted), we useour estimates of mongoose dispersal risk to inves-tigate the evolution of dispersal patterns and re-productive suppression in these gregarious ani-mals.
METHODS
CensusesOur estimates of dispersal risk are based on datafrom the 5 km x 5 km Sangere study area in theSerengeti National Park, Tanzania (2°20' S, 34°50'E), in which the late Jon Rood began freeze-mark-ing and following dwarf mongooses in 1974. Ser-engeti dwarf mongooses live in groups of 2-21 inwhich the oldest, dominant pair produce mdst orall young; subordinates may wait in a queue for thebreeding position, attempt reproduction as a sub-ordinate, or disperse to another group (Creel andWaser, 1991, in press; Rood, 1983, 1987, 1990).
Each marked animal in the study area was locatedin November-December, the beginning of thebreeding season, by Rood in 1975-1986, by Scottand Nancy Creel in 1987-1989, and by Lee Elliottin 1990. Most unmarked animals were captured,sexed, aged by toothwear, marked, and released atthis time. The number of individuals under obser-vation ranged from 81 to 161, distributed in 8—20social groups. From censuses, we calculated threenumbers: N, the number of individuals present onthe study site during a census; P, the number ofindividuals that were philopatric and survived tothe next census; and /c, the number of individualsthat moved into (or formed) new groups on thestudy site and survived to the next census. In de-termining Ic, we included unmarked animals thatimmigrated from off the site and excluded animalsthat left the study site even if they were seen ingroups elsewhere. For this reason, and because an-imals may in principle immigrate more than oncebetween censuses, It is an estimate, rather than adirect measure of /.
Behavioral observationsIf only census data are available, animals must bedefined as dispersing or philopatric based solely ontheir relative locations in two successive censuses.Many investigators, however, will also have obser-vations of individual histories between censuses.For dwarf mongooses, behavioral observations con-tinued throughout most of each year (mean 7.2months, range 2-12). Deaths, immigration and em-igration attempts, and unexplained disappearanceswere recorded ad libitum, as were sightings of tran-sient individuals. Observations of additional groupsoutside the study area were useful in confirmingsome cases of emigration.
From these more detailed individual histories,we counted the number of animals that disap-
1S6 Behavioral Ecology Vol. 5 No. 2
at Montana State U
niversity Library on Septem
ber 8, 2015http://beheco.oxfordjournals.org/
Dow
nloaded from

peared, D. Some disappearances were known to beemigrations, others were known to be deaths in thegroup, and still others could not be allocated toeither category and so were scored as "unex-plained."
We scored an emigration under two circumstanc-es: when a recognizable individual disappeared fromone group and was subsequently found as a mem-ber of another, or when an individual disappearedafter having been sighted outside its group (at leastseveral hundred meters away and, usually, outsideits group's normal home range). Emigrants definedby these criteria returned to their groups on onlyfour occasions, with the shortest period spent as atransient exceeding 2 weeks.
We scored a death when mortality by naturalcauses (predation, fire) was observed, when an iden-tifiable animal was found dead, or when a dominantanimal of either sex disappeared and was not seenagain (only 7 of 135 dominant animals that dis-appeared were subsequently found in anothergroup). We scored an unexplained disappearancewhenever a recognizable subordinate was not sight-ed for a month or more but the criteria for emi-gration were not met.
Finally, we scored an immigration whenever agroup was joined for at least 1 month by a markedindividual from another group or by an unmarkedindividual whose age precluded its having been bomin the group. We also scored an immigration foreach of two or more opposite-sexed emigrating in-dividuals that initiated an association on the studysite lasting at least a month, established a territory,and thus formed a new group. We summed all casesof immigration into groups on the study area toobtain a "behavioral" measure of the number ofimmigrants, lb. Like Ic, lb is an estimate of / andassumes that the number of emigrants that succeedin joining groups off the study site equals the num-ber of unmarked animals successfully immigratingonto the site. Ib may be larger than /,. (for mon-gooses, it is 3% larger) because it includes animalsthat immigrated but died before the next census.
Bracketing «„ «, and sp
From annual censuses, we can measure the overallsurvival rate, (P + Ie)/N, but we cannot determinethe survival rates associated with dispersal (s,) orphilopatry (s^) unless we know the proportion ofthe population that emigrates («). We do know,however, that e, se and s, are related:
s,= N-e
W ~ e)
(1)
(2)
We therefore have two equations describing rela-tionships among the three unknowns.
For mongooses, we plotted s, and sp as a functionof e. We then estimated bounds on s,, e, and sp giventhe assumptions that neither of the two survivalrates can exceed one, and that individuals in fa-miliar groups or home ranges survive at least aswell as those that are dispersing.
We also used behavioral observations to bracketthe proportion of emigrants. If all dispersal at-
tempts had been detected, so that all unexplaineddisappearances were deaths in the group, then Ewould be simply the number of observed emigra-tions. On the other hand, if deaths in the grouphad been reliably detected so that unexplained dis-appearances were in fact emigrations, then E wouldbe die sum of observed emigrations and unex-plained disappearances. In either case, we can cal-culate e = E/N. The assumption that unexplaineddisappearances are deaths in die group leads to alower bound for 5,, while the assumption that theyare emigrations leads to an upper bound.
We bracketed sr for the sexes separately; withineach sex, we also looked at the effects of age. Todo this we broke the male sample into three ageclasses: yearlings, 2-3 year olds, and older animals.Because emigration was rare among older females,we compared yearlings with all odier age classes.These breakdowns gave us sample sizes of 163—461animals, 16-43 emigrants, and 40-85 unexplaineddisappearances per analysis.
Estimating survival rates using a subset of thepopulation that rarely emigrates
To estimate survival rates, we assumed that philo-patric mongooses of all age-sex classes had thesame annual mortality rate and measured that ratein a class of individuals that rarely or never emi-grated. In dwarf mongooses, Rood (1987) identi-fied three classes of individuals that rarely emigrate:females more than 3 years old, dominant females(of any age), and dominant males. All three classesof individuals had annual mortality rates of ap-proximately 0.3 (Rood, 1990). We therefore allo-cated 0.3 of the D disappearing animals to philo-patric mortality, assumed the balance to haveemigrated, and calculated s, as Ib/Q.7D. We choseto use /t instead of /,. in this analysis because wehad determined D using behavioral as well as censusobservations.
We determined confidence limits around this lastestimate of s, using a bootstrap procedure (Efronand Gong, 1983). To do this, we treated each an-nual pair of /» and E estimates as one data point.The bootstrap procedure estimates the distributionof / , /£ from these data. We wrote a SAS programto draw a "bootstrap sample": 14 random draws,with replacement, from the 14 annual values of Itand E. The program then calculated the "bootstrapreplication," the value of It/E for this sample, andrepeated this procedure 1000 times. The 5th and95th percentiles of the histogram of 1000 boot-strap replications provide 95% confidence intervalsaround /»/£ (the "percentile method," Efron andGong, 1983).
RESULTS
Characteristics of dispersal
Over the entire study period, including both censusand behavioral observations, we detected 160 im-migrations, 187 emigrations, 137 deaths, and 341unexplained disappearances. The general charac-teristics of dwarf mongoose dispersal over the en-tire study period did not differ from those de-scribed by Rood (1987). Males and females exhibitedqualitatively similar patterns of emigration widi re-
Waser ct al. • Dispersal risk 1S7
at Montana State U
niversity Library on Septem
ber 8, 2015http://beheco.oxfordjournals.org/
Dow
nloaded from
40 r
• Females• Males
0 2 4 6 8Age (years)
10 12 14
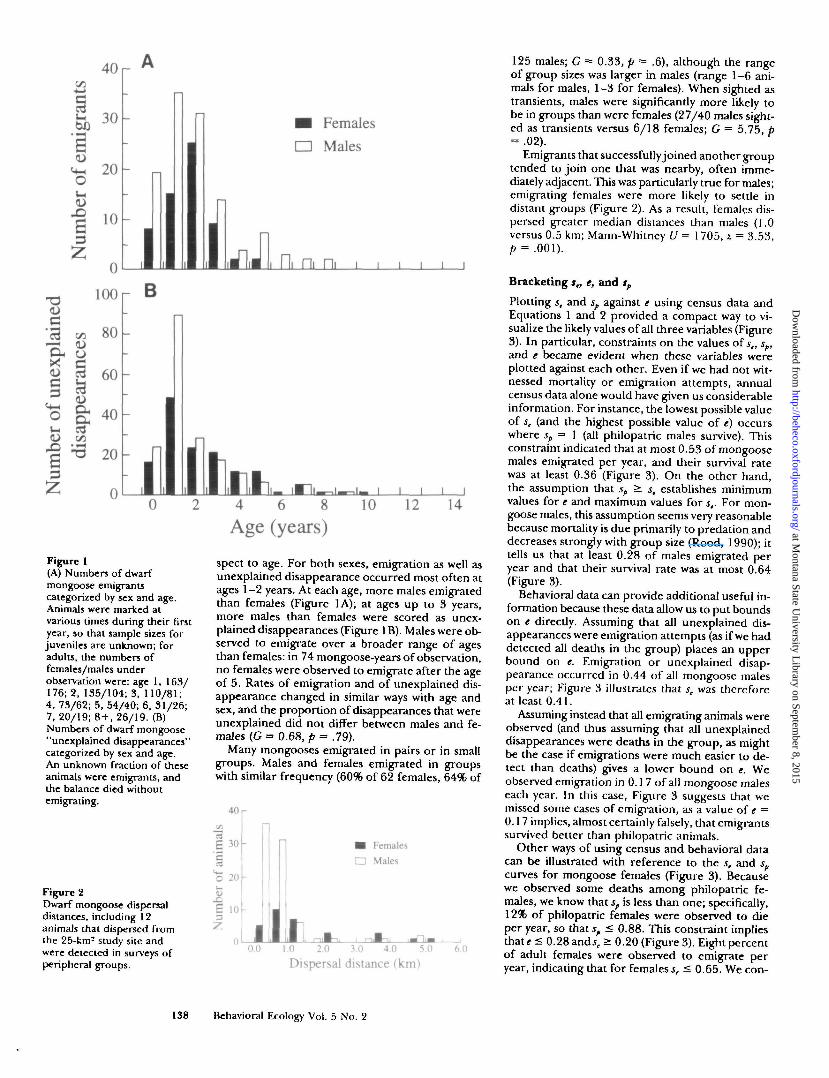
Figure 1(A) Numbers of dwarfmongoose emigrantscategorized by sex and age.Animals were marked atvarious times during their firstyear, so that sample sizes forjuveniles are unknown; foradults, the numbers offemales/males underobservation were: age 1, 163/176; 2, 135/104; 3, 110/81;4, 73/62; 5, 54/40; 6, 31/26;7, 20/19; 8 + , 26/19. (B)Numbers of dwarf mongoose"unexplained disappearances"categorized by sex and age.An unknown fraction of theseanimals were emigrants, andthe balance died withoutemigrating.
spect to age. For both sexes, emigration as well asunexplained disappearance occurred most often atages 1-2 years. At each age, more males emigratedthan females (Figure 1A); at ages up to 3 years,more males than females were scored as unex-plained disappearances (Figure 1B). Males were ob-served to emigrate over a broader range of agesthan females: in 74 mongoose-years of observation,no females were observed to emigrate after the ageof 5. Rates of emigration and of unexplained dis-appearance changed in similar ways with age andsex, and the proportion of disappearances that wereunexplained did not differ between males and fe-males (G = 0.68, p = .79).
Many mongooses emigrated in pairs or in smallgroups. Males and females emigrated in groupswith similar frequency (60% of 62 females, 64% of
4 0 r
c30 -
20 -
Figure 2Dwarf mongoose dispersaldistances, including 12animals that dispersed fromthe 25-km? study site andwere detected in surveys ofperipheral groups.
2
z10 -
11
• Females• Males
II . .0.0 1.0 2.0 3.0 4.0 5.0
Dispersal distance (km)6.0
125 males; G = 0.33, p = .6), although the rangeof group sizes was larger in males (range 1-6 ani-mals for males, 1-3 for females). When sighted astransients, males were significandy more likely tobe in groups than were females (27/40 males sight-ed as transients versus 6/18 females; G = 5.75, p= .02).
Emigrants that successfullyjoined another grouptended to join one that was nearby, often imme-diately adjacent. This was particularly true for males;emigrating females were more likely to settle indistant groups (Figure 2). As a result, females dis-persed greater median distances than males (1.0versus 0.5 km; Mann-Whitney U = 1 705, z = 3.53,p = .001).
Bracketing sa e, and sp
Plotting s, and sp against e using census data andEquations 1 and 2 provided a compact way to vi-sualize the likely values of all three variables (Figure3). In particular, constraints on the values of s,, sp,and e became evident when these variables wereplotted against each other. Even if we had not wit-nessed mortality or emigration attempts, annualcensus data alone would have given us considerableinformation. For instance, the lowest possible valueof s, (and the highest possible value of e) occurswhere sp = 1 (all philopatric males survive). Thisconstraint indicated that at most 0.53 of mongoosemales emigrated per year, and their survival ratewas at least 0.36 (Figure 3). On the other hand,the assumption that sp > s, establishes minimumvalues for e and maximum values for s,. For mon-goose males, this assumption seems very reasonablebecause mortality is due primarily to predation anddecreases strongly with group size (Rood, 1990); ittells us that at least 0.28 of males emigrated peryear and that their survival rate was at most 0.64(Figure 3).
Behavioral data can provide additional useful in-formation because these data allow us to put boundson e directly. Assuming that all unexplained dis-appearances were emigration attempts (as if we haddetected all deaths in the group) places an upperbound on e. Emigration or unexplained disap-pearance occurred in 0.44 of all mongoose malesper year; Figure 3 illustrates that s, was thereforeat least 0.41.
Assuming instead that all emigrating animals wereobserved (and thus assuming that all unexplaineddisappearances were deaths in the group, as mightbe the case if emigrations were much easier to de-tect than deaths) gives a lower bound on e. Weobserved emigration in 0.1 7 of all mongoose maleseach year. In this case, Figure 3 suggests that wemissed some cases of emigration, as a value of e =0.17 implies, almost certainly falsely, that emigrantssurvived better than philopatric animals.
Other ways of using census and behavioral datacan be illustrated with reference to die s, and spcurves for mongoose females (Figure 3). Becausewe observed some deaths among philopatric fe-males, we know that sp is less than one; specifically,12% of philopatric females were observed to dieper year, so that s, < 0.88. This constraint impliesthat* < 0.28 ands, > 0.20 (Figure 3). Eight percentof adult females were observed to emigrate peryear, indicating that for females se < 0.65. We con-
138 Behavioral Ecology Vol. 5 No. 2
at Montana State U
niversity Library on Septem
ber 8, 2015http://beheco.oxfordjournals.org/
Dow
nloaded from

elude drat, for adult male mongooses, 0.41 < sc <0.64, and for adult females, 0.20 < s. < 0.65.
Figure 4 illustrates the ranges of possible valuesof s,, sf, and e for mongoose males and females ofdifferent age classes. Breaking the data down byage suggested that emigrating yearling males hadlow survival rates and that survival rate increasedwith age. For females, bounds on s, were too broadto suggest variation in survival rate with age. Aparallel analysis suggests that survival rates for phil-opatric animals, sp, were similar for both sexes andall ages. Proportions of animals that emigrated, e,declined with age for both sexes and were generallyhigher for males than for females (Figure 4).
Estimating s, using a subset of thepopulation that rarely emigrates
Assuming that 0.3 of the disappearing animals diedat home and the balance emigrated, we estimateddiat mongoose male emigrants survived at a rateof 0.49 and females at a rate of 0.22. The 95%confidence limits did not overlap (males 0.37-0.62,females 0.12-0.33).
Assuming the mortality rate of philopatric ani-mals is independent of age and using die sameapproach, we found patterns parallel to those ob-tained by bracketing s, estimates. Male and femaleyearlings survived at virtually identical rates; thesurvival rate of male emigrants increased with age,but that of older females did not (Table 2).
DISCUSSION
Though many studies have produced the kinds ofdata used in the analyses suggested here, few haveestimated dispersal risk using these or similar medi-ods (but see Bowen et al., 1989; Fitzpatrick andWoolfenden, 1986; Koenig and Mumme, 1987;Wiggett and Boag, 1989). We argue that, althoughdie details of die approach may vary with the spe-cies and estimates may never be precise, we maybe able to glean more about st, sp, and e than hasgenerally been appreciated. Simply knowing theupper and lower bounds of s, will permit a sensi-tivity analysis, and simple census data are oftenenough to substantially narrow diese bounds. Spe-cies for which s, can be tightly bracketed will includethose widiin which a large proportion of a partic-ular age class immigrates: s, is relatively insensitiveto the value of e when Ic/N is large (Equation 1).For instance, s, for female mongooses is highly sen-sitive to the estimate of e; s, for male mongooses,which immigrate more often, is less sensitive (Fig-ure 3). It will also be possible to put reasonablytight bounds on the probability of surviving dis-persal for species in which emigration is a conspic-uous act (so that most disappearances are ex-plained).
Our results for mongooses suggest that differenttypes of census and behavioral data can be com-bined to narrow the bounds on s,. Some investi-gators may have data (or be willing to make as-sumptions) that let them place limits on e, otherson sp; our approach has die advantage of makingthe consequences of such information visualizablein a common framework. Often, an upper or lowerlimit of s, alone will be of value. For instance, givendiat we know diat at most 0.63 of emigrating male
1.0
0.8
0.6
0.4
0.2
0.01.0
0.8
0.6
0.4
0.2
Males
i i i i i i i i
00.
-
-"A
i i i i i i i
Females
0.0 0.2 0.4 0.6 0.8 1.0
Proportion emigrating (e)
mongooses survive, we could conclude that selec-tion would not favor dispersal for inbreeding avoid-ance alone unless die cost of inbreeding exceeded0.37 (Bengtsson, 1978).
There are many species for which die distinctionbetween "philopatry" and "emigration" is not aneat dichotomy; young animals may engage in re-peated forays away from die natal site, searchingfor opportunities to immigrate. This problem hasparticularly plagued investigators of cooperativelybreeding birds, for which such forays are commonbut inconspicuous. For example, Filzpatrick andWoolfenden (1986) estimate a parameter Do similar
2 s
1.0 -
0.8 -
? -^ 0.6 \-
3 £ 0.4
0.2
0.0
B e 0.82 S« -g 0.6> w
I g. 0.4" 1 0.2
0.0
10c st A Io c u-8
I 1 0.40.2
0.0
-
•
- •
•
D
Males
D
I
a
i
a
i
•
i
a
i
DI
Females
•i
•
i
•
*
ai
D
1
0
Figure 3Estimated survival rates foremigrants (i,) and philopatricanimals (J,) as a function ofthe proportion of individualsthat emigrated (*), calculatedfrom annual census data usingEquations 1 and 2. Data forall age classes combined. Asdescribed in the text, thesegraphs may be used todetermine the bounds on e, sf,and sr.
1 2 - 3 4 + 1 2 +
Age (years)
Figure 4Upper bounds (circles) andlower bounds (squares) of thesurvival rate of emigrants (j,)and of nondispersers (s,), aswell as the proportion ofanimals that emigrate (e), as afunction of sex and age class.
Waser ct al. • Dispersal risk 1S9
at Montana State U
niversity Library on Septem
ber 8, 2015http://beheco.oxfordjournals.org/
Dow
nloaded from

Table 2Probabilities of surviving dispersal (»,) as a function of age and sex, assuming that 0.7 of all disappearances aremigration attempts
Age
Sex Yearlings 2-3 years 4+ years
MalesFemales
0.28 (0.13-0.42)"0.31 (0.16-0.51)
0.51 (0.36-0.68)0.21 (0.11-0.32)
0.59 (0.40-0.77)
" Bootstrapped 95% confidence limits in parentheses.
to our s, by dividing I, by the total number of non-breeders in the population. The justification forthis approach is that it indicates what the conse-quences would be if all nonbreeders tried to dis-perse; it would equally be appropriate if all non-breeders were continuously monitoring othergroups for breeding vacancies. Under such circum-stances, it might make sense to assume that mostunexplained disappearances are emigration at-tempts; the consequences of this assumption wouldbe clear on plotting s, against e.
Refining the estimate ois, using values of sp fromnondispersing subsets of the population requiresstrong support for the assumption that sp is con-stant across the relevant age-sex classes. One checkon the validity of this assumption for mongoosesis shown in Figure 4: upper and lower bounds forsp for all age-sex classes included the value ob-served in nondispersing age-sex classes, 0.7. Ingeneral, the approach will be most justifiable inspecies for which dispersal is confined to one sexor age class, and sources of mortality in the groupare similar among sexes (or age classes). In suchcases, bootstrap confidence intervals give a mea-sure of the certainty of the s, estimate given annualvariation in immigration, emigration, and deathrates.
Estimates of the probability of successful dis-persal will need correction if the study site as awhole is either a source or a sink for emigrants.The impact of this assumption is influenced by thesize of the study area: the higher the proportionof observed immigrants that originate within thestudy site, the lower the potential error in esti-mating 5,. In the worst case that none of the emi-grants from our study area had survived off thesite, our estimates of s, would be 40% too high.
In fact, emigrants from the study site were knownto settle successfully up to several kilometers away(Figure 2). Throughout the study, surrounding ar-eas sustained stable mongoose populations at den-sities similar to those under observation. The agestructure of three groups off the study site wasvirtually identical to that on the study site (G =2.19, df = 7, p = .9). Thus, it is likely that immi-gration and emigration were in balance.
We emphasize that the estimates of s, we reportare not estimates of the chances of "effective" dis-persal (Greenwood, 1980); many mongooses suc-cessfully establish themselves in a new group yetstill fail to breed. However, if effective dispersal isthe phenomenon of interest, the approach can eas-ily be modified by replacing fc with the number ofanimals that immigrate and breed.
The indirect approach is not applicable only togroup-living species. Emigration from and immi-gration into specific local areas, territories, orsleeping sites could be examined in the same waysas movements in and out of groups. What is re-quired is a site within which enough animals aremarked and monitored to allow reliable detectionof immigration and emigration.
For dwarf mongooses, the results of our dispersalrisk analyses immediately suggest interesting pat-terns that had not previously been apparent. Whilewe might have guessed that older, larger, moreexperienced mongooses would be more successfulat dispersing than juveniles and yearlings, there wasnothing to suggest that one sex might be more atrisk than the other (Figure 4 and Table 2). Dwarfmongooses are monomorphic and would seemequally vulnerable to predators or starvation dur-ing dispersal.
Several explanations are plausible. It is conceiv-able that transient males gain a survival advantagefrom their somewhat greater tendency to travel inpairs and small groups. Another possibility is thatmales are more successful than females at gainingentrance into new groups. This seems plausible,given that males immigrate in pairs or groups moreoften than females (Rood, 1987) and that immi-grating males are much more likely than femalesto kill or evict the same-sexed resident breeder(Rood, 1990). Emigrant survival rates would beexpected to increase with age if much of the riskinvolved is in overcoming the opposition of same-sexed residents during immigration. However, itdoes not necessarily follow that females are lesssuccessful in immigrating because they usually at-tempt to enter a new group singly. While residentmales (and especially the "beta" male) often force-fully resist male immigrants, interactions betweenimmigrating and resident females are generallypeaceful (Rood, 1986, 1987).
Another possibility is that females survive lesswell during dispersal because they are more selec-tive in the groups that they attempt to join thanare males, thus prolonging the length of time theylive as vulnerable transients. The somewhat longerdispersal distances observed for females (Figure 2)are consistent with this possibility, as is the obser-vation that females rarely immigrate into groupscontaining more than one resident female (Rood,1987). Thirty-six percent of females that did im-migrate (N = 39) immediately achieved a breedingposition, versus only 20% of males (N = 117; G •=3.83,/) = .05). Dispersing females markedly shortenthe queue in which they must wait to acquire the
140 Behavioral Ecology Vol. 5 No. 2
at Montana State U
niversity Library on Septem
ber 8, 2015http://beheco.oxfordjournals.org/
Dow
nloaded from

breeding position, whereas dispersing males short-en it only slightly (Creel and Waser, in press). Theseresults raise intriguing questions about the payoffsto male and female residents of resisting immigra-tion, and the payoffs for selectivity among male andfemale dispersers.
Dispersal risk analyses also suggest interestingpossibilities for cross-species comparisons. The lowprobability of success associated with dwarf mon-goose emigration is consistent with the commonexpectation that risky dispersal or saturated habitatis associated with philopatry and cooperative rais-ing of young (Brown, 1974, 1987; Emlen, 1982;Woolfenden and Fitzpatrick, 1984). Is the proba-bility of successful dispersal equally low in othercooperative breeders, and is it higher in speciesdiat are less cooperative? The answers will comewhen parallel analyses are pursued on other spe-
We thank Hazel, Josephine, and Jon Rood for access totheir study site and data, for their generous logistic as-sistance in the field, and for their friendship. Many of therecent observations were contributed by Nancy Creel andLee Elliott. Norm Henderson suggested the possibility ofthe bootstrap procedure, and George McCabe assuredthat it came to fruition. The manuscript benefited fromthe comments of Kellie Herbst, Rick Howard, Brian Keane,Don Kramer, Kerry Rabenold, and two anonymous re-viewers. Karim Hirji, David Babu, B. Maragesi, the Tan-zania National Parks, the Tanzania National ResearchCouncil, and the Serengeti Wildlife Research Institutemade it possible to carry out this work. The work wasfunded in part by the National Geographic Society andthe National Science Foundation (BSR 88180040).
REFERENCES
Anderson P, 1989. Dispersal in rodents: a resident fitnesshypothesis. Shippensburg, Pennsylvania: American So-ciety of Mammalogists Special Publication No. 9.
Bengtsson BO, 1978. Avoiding inbreeding: at what cost?J Theor Biol 73:439-444.
Bowen BS, Koford RR, Vehrencamp SL, 1989. Dispersalin the communally breeding groove-billed ani (Crotoph-aga sulcirostris). Condor 91:52-64.
Boyce CCK, Boyce JL III, 1988. Population biology ofMicrotus arvalis. J Anim Ecol 57:723-736.
Brown JL, 1974. Alternate routes to sociality in jays, witha theory for the evolution of altruism and communalbreeding. Am Zool 14:63-80.
Brown JL, 1987. Helping and communal breeding inbirds. Princeton, New Jersey: Princeton University Press.
Chepko-Sade BD, Halpin ZT (eds), 1987. Mammalian dis-persal patterns: the effects of social structure on pop-ulation genetics. Chicago: University of Chicago Press.
Creel SR, Waser PM, 1991. Failures of reproductive sup-pression in dwarf mongooses: accident or adaptation?Behav Ecol 2:7-16.
Creel SR, Waser PM, in press. Inclusive fitness and re-productive strategics in dwarf mongooses. Behav Ecol.
Efron B, Gong G, 1983. A leisurely look at the bootstrap,the jackknife, and cross-validation. Am Stat 37:36-48.
Emlen ST, 1982. The evolution of helping behavior anecological constraints model. Am Nat 119:29-39.
Fitzpatrick JW, Woolfenden GE, 1986. Demographicroutes to cooperative breeding in some new world jays.In; Evolution of animal behavior, paleontological andfield approaches (Nitecki MH, Kitchell JA, eds). NewYork: Oxford University Press; 137-160.
Garrett MG, Franklin WL, 1988. Behavioral ecology ofdispersal in the black-tailed prairie dog. J Mammal 69:236-250.
Greenwood PJ, 1980. Mating systems, philopatry and dis-
persal in birds and mammals. Anim Behav 28:1140-1162.
Holekamp K, 1984. Dispersal in ground-dwellingsciurids.In: The biology of ground-dwelling squirrels (MurieJO, Michener GR, eds). Lincoln: University of Ne-braska Press; 295-320.
Johnson ML, Gaines MS, 1990. Evolution of dispersal:uieoretical models and empirical tests using birds andmammals. Annu Rev Ecol Syst 21:449-480.
Jones WT, 1982. Natal nondispersal in kangaroo rats (PhDthesis). West Lafayette, Indiana: Purdue University.
Koenig WE, Mumme RL, 1987. Population ecology ofthe cooperatively breeding acorn woodpecker. Prince-ton, New Jersey: Princeton University Press.
Leuze CCK, 1980. The application of radio tracking andits effect on die behavioral ecology of die water vole.In: A handbook on biotelemetry and radio tracking(Amlaner CJ, MacDonald DW, eds). Oxford: PergamonPress; 361-366.
Pollock KH, Winterstein SR, Bunck CM, Curtis PD, 1989.Survival analysis in telemetry studies: the staggered en-try design. J Wildl Manage 53:7-15.
Rood JP, 1983. The social system of die dwarf mongoose.In: Recent advances in the study of mammalian behav-ior (Eisenberg JF, Kleiman DG, eds). Shippensburg,Pennsylvania: American Society of Mammalogists Spe-cial Publication No. 7; 454-488.
Rood JP, 1986. Ecology and social evolution in the mon-gooses. In: Ecological aspects of social evolution: birdsand mammals (Rubenstein DI, Wrangham RW, eds).Princeton, New Jersey: Princeton University Press; 131-152.
Rood JP, 1987. Dispersal and intergroup transfer in thedwarf mongoose. In: Mammalian dispersal patterns,the effects of social structure on populadon genetics(Chepko-Sade BD, Halpin ZT, eds). Chicago: Universityof Chicago Press; 85-103.
Rood JP, 1990. Group size, survival, reproduction, androutes to breeding in dwarf mongooses. Anim Behav39:566-572.
van Vuren D, Armitage KB, in press. Survival of dispers-ingand philopatric yellow-bellied marmots: what is thecost of dispersal? Oikos.
Vehrencamp SL, 1983. A model for the evolution ofdespotic versus egalitarian societies. Anim Behav 31:667-682.
Waser PM, Jones WT, 1983. Natal philopatry among sol-itary mammals. Q Rev Biol 50:355-390.
Wiggett DR, Boag DA, 1989. Intercolony natal dispersalin die Columbian ground squirrel. Can J Zool 67:42—50.
Woolfenden G, FitzpatrickJ, 1984. The Florida scrub jay:demography of a cooperatively breeding bird. Prince-ton, New Jersey: Princeton University Press.
Waser et al. • DUpersal risk 141
at Montana State U
niversity Library on Septem
ber 8, 2015http://beheco.oxfordjournals.org/
Dow
nloaded from




















