
guidelines-for-positioning-and-fixation-in-pcb-less-assembly ...
Upload
independentCategory
view
0download
0
World Journal of Microbiology and Biotechnology 9, 444-454
Special Topic Review
Control of the expression of bacterial genes involved in symbiotic nitrogen fixation
M. Megias,* J.L. Folch and C. Sousa
Several genera of NJixing bacteria establish symbiotic associations with plants. Among these, the genus Rhizobiurn has the most signiticant contribution, in terms of yield, in many important crop plants. The establishment of the Rhizobiunrlegume symbiosis is a very complex process involving many genes which need to be co-ordinately regulated. In the lirst instance, plant signal molecules, known to be flavonoids, trigger the expression of host-specific genes in the bacterial partner through the action of the regulatory NodD protein. In response to these signak, RGzo6ium bacteria synthesize llpo-oligosaccharide molecules which in turn cause cell differentiation and nodule development. Once the nodule has formed, Rhizobium cells differentiate into bacteroids and another set of genes is activated. These genes, designated nif and fix, are responsible for N, fixation. In this system, several regulatory proteins are involved in a complex manner, the most important being NifA and a two component (FixK and FixL) regulatory system. Our knowledge about the establishment of these symbioses has advanced recently, although there are many questions yet to be solved.
R$ WOT&: Bacteria, expression, genes, nitrogen fixation, symbiotic
Bacteria-plant interactions range from the pathogenic, in which bacteria parasitize a plant, to the symbiotic, in which two organisms help each other to survive under otherwise difficult environmental conditions. Bacteria of the genus Rhizobium are able to interact symbiotically with specific legume host plants, leading to the formation of N,-fixing root nodules. Nodules are specialized organs, within which the rhizobia, differentiated into bacteroids, utilize plant- provided C compounds and reduce atmospheric N, to ammonia for its assimilation by the plant (Long 1989). In return, the bacteria obtain C sources (citric acid cycle intermediates like malate and succinate). As this association is normally of mutual benefit to both the invader (Rhizobium cells) and the invaded (plant host), it is called symbiosis.
The incorporation of atmospheric N, into organic material which results from the Rhizobiurn-legume symbiosis is estimated to account for one third of the total N needed for world agriculture (Economou & Downie 1992). This unique intracellular association contributes significantly towards agricultural yield. The agronomic importance of this symbiotic association has thus justified extensive research
M. Meglas, J. L. Folch and C. Sousa are with the Departamento de Microbiologla y Parasitologia, Univenidad de Sevilla, Apdo 674, 41012 Sevilla, Spain; fax: +34 64 626162. ‘Corresponding author.
@I 7993 Rapid Communications of Oxford Ltd
over the past 100 years and the association is probably one of the best known of all plant-microbe interactions (Economou & Downie 1992; Fischer & Long 1992).
Nitrogen-fixing bacteria are taxonomically diverse and occupy various ecological niches. Some genera in the Rhizobiaceae (Rhizobium, Brudyrhizobitrm and Az.orhizobium) nodulate legumes whereas other members (Agrobacterium) are plant pathogens (Young & Johnston 1989). In an agricultural context, the most important association is that between members of the Leguminosae and Rhizobium, so not surprisingly this has received the most detailed study and will be the subject of this review. Each bacterial species is restricted to its specific group of host plants, thus defining ‘cross-inoculation groups’ in which the bacteria are classified according to their ability to establish symbiosis with a particular group of plants.
Successful nodulation is a host-specific process in the sense that Pisum and Viciu species are host plants for R. leguminosarum biovar (bv.) viciae, Medicago and Melilotus are hosts for R. meliloti, and Trifolium spp. are hosts for R. legtlminosamm bv. trifolii. Members of the genus Rhizobittm are relatively fast growing strains and have a narrow host range. On the other hand, the genus Bradyrhizobium contains a group of slow-growing rhizobia that generally nodulate quite a wide range of host plants.

Regulation of symbiotic nitrogen fixation
al. 1988; Barbour et al. 1991). Chemoattraction by flavonoids has also been reported (Caetano-Anolles et al. 1988; Barbour et al. 1991). The bacteria attach to the root hair surface probably via the specific Cat+-binding protein rhicadhesin and then more bacteria accumulate on the ones which have already adhered, this process being mediated by bacterial cellulose fibrils (Smit et al. 1987). On the other hand, such an attachment may not be necessary for nodulation to proceed, at least in soybeans.
Subsequently, the root hairs curl (Hat) into a so-called ‘shepherd’s crook’ shape (Smit et al. 1987), thus entrapping the bacteria within the curl (Yao & Vincent 1969; Bhuvaneswari et al. 1980), and the plant responds by forming thick and short roots (Tsr) (Van Brussel et al. 1990).
The infection starts with a very localized hydrolysis of fhe plant cell wall in the root hair curl (Ridge & Rolfe 1986). At the site of cell wall hydrolysis, bacteria enter the root hair cell by invagination of the plasma membrane. Around the invaginated membrane, the plant forms a hollow ingrowth which develops into a tubular tunnel-like structure, the infection thread, which penetrates further into the root hair cell. The bacteria multiply along the growing infect-ion thread and ultrastructural studies show them to be surrounded by a matrix-glycoprotein (plant-derived), their own extracellular polysaccharides and a mucigel composed of cell-wall polysaccharides (VandenBosch et al. 1989;
Economou & Downie 1992). The plant proteins that are specifically synthesized during the formation and function of a root nodule are called nodulins. During these early stages of infection, early nodulins (ENOD) are expressed (Nap & Bisseling 1990). Simultaneously with the growth of the infection thread, some cortex cells start to divide, resulting in a nodule primordium. The infection threads carry
In this symbiosis an exchange of signals between the host plant which releases flavonoids, and the bacterium, which releases so-called Nod metabolites, plays a crucial role. Lately, the Rhizobium-legume interaction has received much attention because of the finding of a small lipo- oligosaccharide molecule (Figure 1 and Table 1) which alone, at a very low concentration (lo-l1 M), causes nodule development in alfalfa plants (Lerouge et al. 1990). The molecule is synthesized in response to flavonoid compounds exuded by the root.
The development of symbiosis fakes place in several stages, confrolled by genes in the bacterium as well as in the plant. Molecular, genetic and cell biological studies revealed three major, somewhat independent series of events: (1) bacterial attachment to the root hairs, followed by root hair deformation and curling; (2) formation and growth of infection threads in which the bacteria multiply; and (3) induction of meristematic activity in the root cortex, leading to nodule development which culminates in N,-fixing symbiosis.
The sequence of events leading to a nodule starts with chemotactic attraction. Amino acids and dicarboxylic acids are probably the most important chemoattractants under field conditions (Giitz et al. 1982; Caetano-Anolles et
Figure 1. Basic structure of Nod factors of Rhizobium strains.
Table 1. Structures of Nod factors produced by Rhizobium strains.
Rhizobium strain Substitutions* References
4
R. me/i/o0 2011 C16:2
R. leguminosarum bv. C18:4 or C16:l viciae RBL5560
R. melilofi AK41 C16:3
Rhizobium sp. NGR234
C18: 1 or C16:O
6. japonicum USDA110 C18:1, C16:O or C16:l and USDA135
A. caulinodans C18:O or C18: 1 ORS571
R2 R3 X Y
-H or -CO-CH, -SO,H -H -H
-CO-CH, -H -H -H
-H --SOaH -H -H
-CO-CH, 2-CTMethylfucose, -CONH? -CH, 3-Gsulphated or CO-acetylated
-H or -CO-CH, P-O-Methylfucose -H -H or -H
-CONH, or -H o-Arabinose or -H -H -CH,
n
2, 3 Lerouge et al. (1990)
2, 3 Spaink et a/. (1992)
1,2, 3 Schultze et al. (1992)
3 Demont et a/. (1992)
3, 4 Sanjuan et a/. (1992)
2, 3 Mergaert et al. (1992)
l &-Fatty acyl residue: vaccenic acid (C18:1), stearic acid (C18:0), palmitic acid (C16:O); R, and Rx-hydrogen or a different substitution; X-hydrogen or carbonyl residue; Y-hydrogen or Kmethyl residue; n-number of glucosamine units.
World Journal of Microbiology and Biotechndo~y, Vol 9, 1993 445

M. Meg&, JL. Folch and C. Sousa
the bacteria from the root hair to the nodule primordia formed in the root cortex (Dudley et al. 1987).
The bacteria are released from the tips of the infection threads into the host cytoplasm by a process that resembles endocytosis. After their release into the cytoplasm of the plant cell, rhizobia differentiate into bacteroids. The bacteroids lie alone or together, surrounded by a plant membrane derived from the Golgi and endoplasmic reticulum, this membrane is termed the peri-bacteroid membrane (PBM). The bacteroids are pleirnorphic and the fate of the PBM varies from legume to legume. In the case of the pea, the PBM has a volume approximately T-fold that of free-living bacteria. This membrane forms an interface of key importance, essential for the transfer of metabolites between the plant and bacteria. The mature bacteroids are now capable of using nitrogenase to reduce N,, so fixing N in the form of ammonia. Around the start of N, fixation, plant genes encoding so-called late nodulins are expressed. The best known and the most abundant of these is leghaemoglobin (Lb). Lb is an oxihaemoprotein which is localized in the peribacteroid space, with a high 0, affinity, and which resembles the vertebrate globins. Lb controls the concentration of free 0, in the nodule and forms a filter protecting the very O,-sensitive nitrogenase (Nap & Bisseling 1990).
Bacterial genes involved in the nodulation process are: (I) the nod genes (genes involved in host recognition and nodule formation) and also nol genes (for nodulation related); (2) ezo genes, encoding exopolysaccharides, and lipopolysaccharide-encoding genes; and (3) fiz and nif genes, involved in carrying out and supporting N, fixation.
Nodulation Genes
Nodulation genes in Rhizobium are involved in the early steps of the nodulation process. These genes are regulated
by plant-derived signal molecules called flavonoids, through their interaction with the nodD gene product (Rossen et al. 1985; Djordjevic et al. 1987; Horvath et al. 1987).
In Rhizobitrm species, the majority of genes for nodulation and nitrogen fixation are located on high molecular weight Sym (for symbiotic) plasmids aohnson et al. 1978; Nuti et
al. 1979). Other symbiotic loci map on the chromosome (Dylan et al. 1986) or in other plasmids (Toro & Olivares 1986). In contrast, in Bradyrhizobium and Azorhizobittm species, the nod genes are correlated with the chromosome (Van Den Eede et al. 1987).
The organization of nod and no1 genes in several species and biovars is shown in Figure 2. Nodulation genes are involved in the synthesis and transport of small lipo-oligosaccharide molecules which are recognized in a specific manner by the host plant and which by themselves start the organogenesis of the nodule (Lerouge et al. 1990; Spaink et al. 1991, 1992) (Figure 1 and Table 1). In some cases, complete nodule development can be achieved simply by adding these purified factors to the roots of uninoculated alfalfa plants.
The nodABCIj genes were designated as ‘common’ because they can be complemented by the corresponding genes from other strains or species. All the other nod genes, which, unlike the common nod genes, are not’ found in all strains and species and cannot be complemented by nod genes from other species, were designated host-specific nodulation genes (hsn) (Kondorosi et al. 1984). Table 2 presents a summary of the roles that many of the nod genes have.
Regulation of the nodI3 Gene
The nod genes are arranged in several different transcriptional units and these operons are co-ordinately regulated by the nodD product and specific plant signal
Rhizobium melilori
0 TNM. L EF. 0 ASCIJ -ml-n- .,,,,I, .m
+ -d--c-
Rhizobium leguminosarum bv. viciee
X NM, LREF 0 A8CIJ T I I I I II I, ,,.(gn.r I I I I ,I I - -cIcp-
Rhizobium legominosarum bv. trifolii
L Bradyrhizobium japonicum IWIA &I$, 01 rA 6 csu I JNNO Z v w
I I I II I I - C +-L---C----L ---t” -
Figure 2. Genetic organization of the nod and nol genes of R. meliloti, R. leguminosarum bv. viciae, R. leguminosarum bv. trifolii and Bradyrbizobium japonicum. Arrows indicate orientation of nod and no/ gene expression. l Position of the nod boxes; 0 nod genes; n no/ genes; q transcriptional regulatory nodD and syrM genes.
446 World ]oumd of Microbiology and Biotechnology. VOI 9, 1993

Regulation of symbiotic nitrogen fixation
Table 2. Predlcted biochemical function of nod and no/ genes.
nodF””
nodG” nodH”’
nodLmvf
nodM”* nodNmVt nodO’
nodPOmP”
nodT* nods@ no/R”
Predicted function
Proposed to determine the production of Nod factor precursor
Polymerizes the oligosaccharide backbone Transcriptional activator of inducible nod genes Proposed to synthesize Nod factor acyl chain
Proposed to synthetize Nod factor acyl chain
Involved to modify Nod factor fatty acyl side chain Proposed to attach activated sulphate to the
Nacetylglucosamine polymer Proposed to be involved in the excretion of the
signalling molecule Proposed to add an acetyl group to Nod factor
Proposed to synthesize Nod factor sugar subunits Involved in Vicia hirsu~a nodulation Proposed to intern the lipo-oligosaccharide into the
plant cell They could function in the production of an activated
form of sulphate for transfer to Nod factor Proposed to be membrane protein Proposed to add a methyl group to Nod factor Repressor of nodD
Reference
Schmidt et al. (1988), Vargas et a/. (1990), VIzquez et a/. (1991)
Bulawa 8 Wasco (1991), Rodriquez-Quifiones .et a/. (1989) Schlaman et a/. (1992) D&bell& & Sharma (1986). Bibb et a/. (1989),
Spaink et a/. (198ga), Vargas et al. (1990) Shearman et a/. (1986), Sheldon et a/. (1990),
Geiger et al. (1992) Spaink et al. (1991) Schwedock & Long (1990)
Evans & Downie (1986), VAzquez et a/. (1992)
Downie (1989), Bloemberg et a/. (1992), Baev & Kondorosi (1992)
Baev et a/. (1991) Fisher & Long (1992) Downie et a/. (1992)
DBbell& 8. Sharma (1986), Cervantes et a/. (1989), Schwedock & Long, (1990), Folch ef a/. (1992)
Canter-Cremers et a/. (1989) Holsters et al. (1992), Geelen et a/. (1992) Schlaman et al. (1992)
* nod genes are present in R. melilofi (“), R. leguminosarum bv. viciae (‘), R. leguminosarum bv. frifolii (‘), R./eguminosarum bv. phaseoli (‘). R. fropici (“), R. fredii (‘), 6. japonicum (I) and A. caulinodans (“). nodY and nodZ are found in 6. japonicum, nodX has only been found in R. leguminosarum bv. viciae strain TOM. Other genes, including nodK, nodU, no/A, no/E, no/F, no/G, no/H, no// and no/P, have been identified but their possible functions have not been described.
molecules. The nodD gene has been detected in all rhizobia tested so far. In some Rhizobium strains nodD is present as a single copy whereas in others, multiple (two or three) different alleles were found (Rodriguez-Quiiiones et al. 1987). The NodD protein is a transcriptional activator of the nod genes and has different specificity for flavonoids depending on the Rhizobium species from which it comes. This protein shows homology with the LysR family of regulatory proteins. On its amino terminal region there is a sequence which resembles a DNA-binding domain and also a so-called receiver module domain which could allow protein-protein interactions. The carboxy terminus, which varies more than the amino terminus in different species, has been reported to be involved in flavonoid recognition. There is evidence, however, which suggests that this protein does not have two separate functional domains, but rather that the whole tertiary structure is involved in nod gene activation (Spaink et al. 1989b; Sousa et al. 1993).
Transcription of the nodD gene is usually constitutive, but several factors, including combined N, plant signal compounds and regulatory proteins [e.g. the nod repressor protein (NolR), the syrA4 gene product and NodD itself], can regulate its expression (Figure 3) (Rossen et al. 1985;
Horvath et al. 1987; Banfalvi et al. 1988; Dusha et al. 1989;
Kondorosi et al. 1989; Davis & Johnston 1990; Maillet et al. 1990). In this respect, we still know little about the complex regulatory system that controls nodD expression. For example, in R. meliloti groEL mutants (a molecular chaperon) the expression of nodD3 is prevented and that of the nodD1 and nodD2 is reduced when they are expressed constitutively (Fischer & Long 1992).
Most of the different nod operons have a 50 bp (Schofield & Watson 1986) conserved sequence upstream of the promoter region which is the nod-box and to which the nodD protein binds in order to activate transcription. It has been shown by Long (1992) that the nodD protein binds only to one side of the double helix on the nod boxes and causes DNA to bend, resembling the action of the IHF regulatory protein found in other bacterial species (Kustu et al. 1992). It is still not clear whether the NodD protein acts as a monomer or as a multimer in order to activate transcription, although there is some evidence that indicates it could interact with other proteins or with nodD itself to do so (Long 1992). In this respect, a characteristic structure has been found in all nod boxes studied which favours the hypothesis that NodD binds as a tetramer to the nod-box. Other studies involving nod-box deletion mutants also support this model.
World Journal of Microbiology and Bmtechndogy, Vol 9, 1993 447

- Nod factors -
~I”\ ,erJ independent
Figure 3. Cascade regulation of nod gene expression. Arrows indicate regulatory routes. +-Positive controls; controls (Fisher & Long 1992).
NodD protein activity as a tr~s~ptioM1 activator of the nod regulon depends on the protein’s interaction with flavonoids, although it has not been proven that these molecules bind directly to the NodD proteins (Firmin et nl. 1986; Redmond et al. 1986; Bassam et al. 1988; Peters & Long 1988; Zaat et af. 1989; Davis & Johnston 1990). As distinct plant species release different sets of signal molecules (flavones, isoflavones, flavanones and chalcones), NodD proteins can only activate transcription when the right host plant is present, so determining the host specificity (Gyargypal ef al. 1988; Spaink et af. 1989b). Other factors, such as combined nitrogen, can also influence nod gene expression, thus regulating the Rhizobitrtn- legume interaction (Dusha et al. 1989).
Nitrogen Fixation Genes
The genetic basis of symbiotic N, fixation in the Rhiiobiaceae has been the objective of extensive research. The ability of rhizobia to induce N,-fixing nodules requires the specific expression of certain bacterial genes. Those genes homologous to N, fixation genes in Klebsiella peumoniae are referred to as nifgenes. Others, also essential for symbiotic N, fixation but sharing no homology to PC ~~~~iae, are called f;x genes. The presence of these /ix genes correlates with the specific environmental conditions in which symbiotic nitrogen-fixing microorganisms act. This suggests that N, fixation within a root nodule requires a regulatory track different from that in K. ~eumuniae.
The structure and the biochemical properties of the nitrogenase complex are highly conserved, though a marked characteristic of diazotrophs is that they differ in the physiological conditions under which they fix N,. Thus, K. pn~~niae is an enteric bacterium able to use atmospheric N, for growth in the absence of 0, and combined N. On
--negative
the other hand, bacteria of the genus ~~jz~b~u~ fix N, only in symbiotic conditions.
Studies on N, fixation in Rhizobium and Bradyrhizobittm species were facilitated by the previous identification and characterization of 21 contigous nif genes in I( pneumoniae (De Bruijn et al. 1990). These genes are grouped on the chromosome or pSym in Rhizobictm and are organized in different transcriptional units. nif genes of Rhizobitrm which are homologous to those of Klebsiella include nifHDK, the structural genes that encode for the polypeptide subunits of the nitrogenase complex; nifA, the product of which is a positive transcriptional activator for nifHDK and other nif
and fix genes (Ruvkun & Ausubel 1980; Szeto ef al. 1984); nip, nifnr and nifE, genes are involved in the synthesis and processing of the nitrogenase iron-molybdenum cofactor CFeMocof (Norel et al. 1984; Hennecke et al. 1985); and nifF (Hontelez et af. 1984) and nifl (Norel et al. 1984), are genes which encode for nitrogenase-specific electron transport proteins.
Divergent to nigh and upstream of nifA in the R. meli~~fi megaplasmid (pSym1) are the MBCX genes which are transcribed as a single operon from firA to fix (Better et al. 1985; Earl et al. 1987). The fixABCX genes do not exhibit homology to Klebsialla nif genes, and their products may be involved in nitrogenase-specific electron transport. The R. ~elilofj m promoter contains the characteristic nif consensus promoter sequence (Better et a?. 1985) which indicates that it is probably recognized by the product of ntrA and is positively controlled. The f;xX gene is highly homologous to ferredoxin found in other nitrogen-fixing species such as ~ofobacfer vine~an~i~ (Buck & Cannon 1987).
A second fir gene cluster of R. melilofi constitutes the four operons fixL], firK, fixNOQP and fixGH1.S (David et al. 1988;
Batut et al. 1989; Kahn et al. 1989); firLJ and fi& are regulatory genes. fiGHIS and ~x~OQP gene products are thought to be membrane located. FixG has homology with
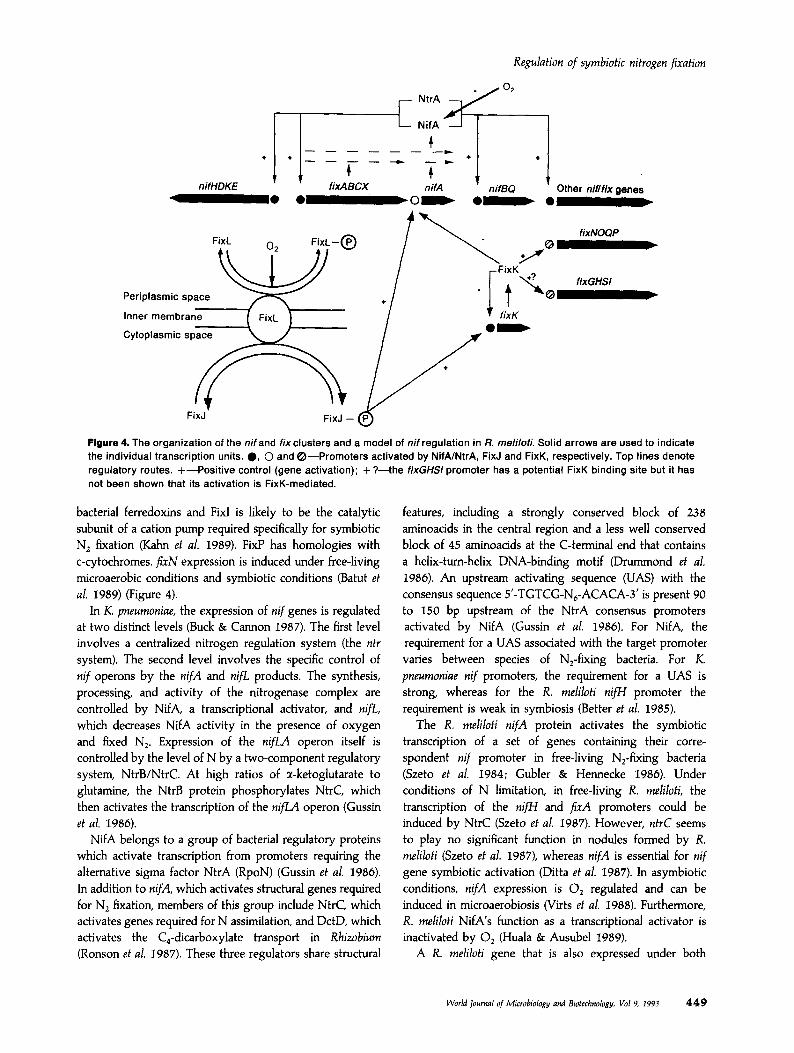
Regulation of symbiotic nitrogen fixation
nifHDKE I I- NifA --I
k -------) +------ -w+ +
k k fixABCX nifA I
nifBQ ’ Other niflfix genes l -o- o-o-
4\ FixL
t\ Y2
Periplasmic space
Inner membrane
Cytoplasmic space
o fixGH$J
FixJ
Figure 4. The organization of the nif and fixclusters and a model of nif regulation in R. meliloti. Solid arrows are used to indicate the individual transcription units. 0, 0 and 0-Promoters activated by NifAINtrA, FixJ and FixK, respectively. Top lines denote regulatory routes. +-Positive control (gene activation); + ?-the fixGHS/ promoter has a potential FixK binding site but it has ._ not been shown that its activation is FixK-mediated.
bacterial ferredoxins and Fix1 is likely to be the catalytic subunit of a cation pump required specifically for symbiotic N, fixation (Kahn et al. 1989). FixP has homologies with c-cytochromes. f;N expression is induced under free-living microaerobic conditions and symbiotic conditions (Batut et al. 1989) (Figure 4).
In K. pneumoniae, the expression of nif genes is regulated at two distinct levels (Buck & Cannon 1987). The first level involves a centralized nitrogen regulation system (the ntr system). The second level involves the specific control of nif operons by the nifA and nifL. products. The synthesis, processing, and activity of the nitrogenase complex are controlled by NifA, a transcriptional activator, and nip, which decreases NifA activity in the presence of oxygen and fixed N,. Expression of the nifLA operon itself is controlled by the level of N by a two-component regulatory system, NtrB/NtrC. At high ratios of a-ketoglutarate to glutamine, the NtrB protein phosphorylates NtrC, which then activates the transcription of the nifL4 operon (Gussin et al. 1986).
NifA belongs to a group of bacterial regulatory proteins which activate transcription from promoters requiring the alternative sigma factor NtrA (RpoN) (Gussin et al. 1986). In addition to nifA, which activates structural genes required for N, fixation, members of this group include NtrC, which activates genes required for N assimilation, and DctD, which activates the C,-dicarboxylate transport in Rhizobium (Ronson et al. 1987). These three regulators share structural
features, including a strongly conserved block of 238 aminoacids in the central region and a less well conserved block of 45 aminoacids at the C-terminal end that contains a helix-turn-helix DNA-binding motif (Drummond et al.
1986). An upstream activating sequence (UAS) with the consensus sequence 5’-TGTCG-N,-ACACA-3’ is present 90 to 150 bp upstream of the NtrA consensus promoters activated by NifA (Gussin et al. 1986). For NifA, the requirement for a UAS associated with the target promoter varies between species of N,-fixing bacteria. For K. pneumoniae nif promoters, the requirement for a UAS is strong, whereas for the R. meliloti nigh promoter the requirement is weak in symbiosis (Better et al. 1985).
The R. melilofi nifA protein activates the symbiotic transcription of a set of genes containing their corre- spondent nif promoter in free-living N,-fixing bacteria (Szeto et al. 1984; Gubler & Hennecke 1986). Under conditions of N limitation, in free-living R. meliloti, the transcription of the nigh and @A promoters could be induced by NtrC (Szeto et al. 1987). However, ntrC seems
to play no significant function in nodules formed by R. meliloti (Szeto et al. x987), whereas nifA is essential for nif gene symbiotic activation (Ditta et al. 1987). In asymbiotic conditions, nifA expression is 0, regulated and can be induced in microaerobiosis (Virts et al. 1988). Furthermore, R. melilofi NifA’s function as a transcriptional activator is inactivated by 0, (Huala & Ausubel 1989).
A R. meliloti gene that is also expressed under both

M. Megias, J.L. Folch and C. Sousa
microaerobic and symbiotic conditions, fixK, represses nifA transcription and is a positive regulator of the jixN operon (Batut et al. 1989). firK is homologous to the E. coli regulatory gene frzr which controls genes expressed under anaerobic conditions. Homologues of the R. meliloti JixK gene have been identified in R. leguminosarum bv. viciae (Colonna-Roman0 et al. 1990), A. caulinodans (De Bruijn el al. 1990) and B. japonicum (Anthamatten & Hennecke 1991). However, unlike these homologues, R. meliloti has FixK which lacks the N-terminal sequence which is thought to be responsible for the 0, sensitivity of other Fnr-like proteins (Batut et al. 1989).
The symbiotic and microaerobic expression of firK and n&A in R. meliloti is positively controlled by a pair of regulatory proteins, FixL and FixJ (David et al. 1988; De Philip et al. 1990). FixJ, the response regulator, is activated by FixL, the 0, sensor, in response to conditions of low 0, tension and then activates expression of nif and fix genes. Sequence data have led to the prediction that FixL and FixJ belong to a family of two-component regulatory systems [see review by Stock et al. (1989)]. FixL is an oxygen-binding haemoprotein and senses oxygen through its haem moiety, which is probably contained between residues 86 and 219 of the polypeptide (Gilles-Gonzhlez et al. 1991). This part of the R. meliloti FixL polypeptide is very well conserved in A. caulinodans (Kaminski & Elmerich 1991) and in B. japonicum (Anthamatten & Hennecke 1991). In addition, in its N-terminus, FixL has two sequences which represent potential transmembrane helices as found in many sensor components of two-component regulatory systems. Look- ing at the properties of most members of the two- component systems (Ronson et al. 1987; Albright et al.
1989), FixL could be proposed as a transmembrane sensor protein, which reacts to a change in the bacterial environment by phosphorylating the cognate activator protein FixJ, which in turn promotes the transcription of nifA and fixK genes. A model of the regulation of N, fixation in R. meliloti is presented in Figure 4. FixJ shows similarity to the N terminus of the regulator class components over the first 120 aminoacids (David et al. 1988; Albright et al.
1989). In E. coli, in an heterologous system, fixK requires higher
levels of the activator FixJ than nifA for activation of transcription. However, the biological significance, if any, of the differential expression of nifA and fixK remains to be elucidated (De Philip et aI. 1990). In vitro transcription studies of fixK promoter showed that it was dependent on 07’ holoenzyme. Therefore, the activity of R. meliloti nifA promoter is also probably 0”-dependent (Batut et al. 1991).
Oxygen tension seems to be the main signal for nitrogen fixation gene expression within the nodule. However, there are indications that other signals are needed for effective symbiotic gene expression. Thus R. meliloti jixA promoter is weakly activated in microaerobiosis but strongly in
symbiosis (Ditta et al. 1988; A. Cebolla, personal communication). Although induction of nigh expression in microaerobiosis is strongly dependent on UAS, this sequence is not necessary to achieve high levels of expression during symbiosis (Ditta et al. 1988). No activation of nif gene expression was found, either in free-living microaerobic or symbiotic R. meliloti, when a gene involved in the transcriptional activation of the C,-dicarboxylate transport gene was mutated (Birkenhead et al. 1990). On the other hand, in free-living R. meliloti, nifA expression is sensitive to fixed N, levels but not fixK (Nooman et al. 1992). Therefore the expression of structural genes for nitrogen fixation in R. meliloti may then be controlled through other physiological signals in addition to the oxygen status.
In summary, many signals are involved in the expression of the bacterial genes required for the establishment of a mature nodule. Some of them have been elucidated, but others remain to be found. Once all the signals have been discovered a more integrated view of the complex phenomenon of Rhizobium-legume symbiosis will be possible.
Acknowledgements
The authors are grateful to A. Cebolla for critically reading the manuscript. This work was supported, in part, by grants from DGICYT, Spain (BIO 90-S20-c02-01). JLF and CS were supported by fellowships from Ministerio de Asuntos Exteriores, Spain, and Consejeria de Educaci6n y Ciencia, Junta de Andalucia, Spain, respectively.
References Albright, C.M., Huala, E. & Ausubel, F.M. 1989 Procaryotic signal
transduction mediated by sensor and regulatory protein pairs. Annual Reviezo of Genetics 23, 311-336.
Anthamatten, D. & Hennecke, H. 1991 The regulatory status of the /i.rL- and jixJ-like genes in Bradyrhizobium juponicum may be different from that in Rhizobium melilofi. Molecular and General Genetics 225, 38-48.
Baev, N., Endre, G., Petrovics, G., Banfalvi, Z. & Kondorosi, A. 1991 Six nodulation genes of nod box locus 4 in Rhizobium melilofi are involved in nodulation signal production: nodM codes D-glucosamine synthetase. Molecular and General Genetics 228, 113-124.
Baev, N. & Kondorosi, A. 1992. Nucleotide sequence of the R. melilofi nodL gene located in locus n5 of the nod regulon. Plant Molecular Biology 18, 843-846.
Banfalvi, Z., Nieuwkoop, A., Schell, M., Best, L. & Stacey, G. 1988 Regulation of nod gene expression in Brudyrhizobium juponicum. Molecular and General Genefics 214, 420-424.
Barbour, W.M., Hattermann, D.R. & Stacey, G. 1991 Chemotaxis of Bradyrhizobium juponicum to soybean exudates. Applied and Environmenfal Microbiology 57, 2635-2639.
450 World ]oumal of Microbiology and Biotechnology, Vol 9. 1993

Regulation of symbiotic nitrogen fixation
D.&bell&, F. & Sharma, S.B. 1986 Nucleotide sequence of Rhiwbium meliloti RCRLOIl genes involved in host specificity of nodulation. Nucleic Acids Research 14, 7453-7472.
De Bruijn, F., Hilgert, U., Stigter, J., Schneider, M., Meyer, H., Klosse, U. & Pawlowski, K. 1990 Regulation of nitrogen fixation and assimilation genes in the free-living versus symbiotic state. In Nitrogen Fixation: Achievements and Objectives, eds Gresshof, P.M., Roth, L.E., Stacey, G. & Newton, W.E. pp. 33-44. New York: Chapman and Hall.
Demont, N., Roche, P., Aureole, H., Talmont, F., Prom&, D., Prom& J.C., Price, N.P.J., Relic, B., Broughton, W.J., DCbellC F., Ardourel, M.Y., Maillet, F., Rosenberg, C., Truchet, G. & DbnariC, J. 1992 Rhizobium nodulation factors: variations on a theme. In Molecular Genetics of Plant Microbe Interactions, eds Nester, E.W. & Verma D.P.S. pp. 133-141. Dordrecht: Kluwer Academic.
DePhilip, P., Batut, J. & Boistard, P. 1990 Rhizobium meliloti FixL is an oxygen sensor and regulates R. meliloti nifA and fink genes differently in Escherichia coli. ]ournal of Bacteriology 172, 4255-4262.
Ditta, G., Virts, E. & Helinski, D. 1988 Oxygen regulation of nif genes in Rhizobium meliloti. In Molecular Plant-Microbe Interaclions, eds Verma, D.P.S. & Palacios, R. pp. 109-110. St. Paul MN: APS Press.
Ditta, G., Virts, E., Palomares, AJ. & Kim, C.H. 1987 The nifA gene of Rhiwbium meliloti is oxygen regulated. ]oumal of Bacteriology 169, 3217-3223.
Djordjevic, M.A., Redmond, J.W., Batley, M. & Rolfe, B.G. 1987 Clovers secrete specific phenolic compounds which either stimulate or repress nod gene expression in Rhiwbium trifolii. EMBO ]ournal6, 1173-1179.
Downie, J.A. 1989 The nodL gene from Rhizobium leguminosarum is homologous to the acetyl transferases encoded by 1acA and cysE. Molecular Microbiology 3, 1649-1651.
Downie, J.A., Sutton, J.M. & Lea, E.J.A. 1992 nodO: a nodulation protein that forms pores in membranes. In 6th International Symposium on the Molecular Genetics of Plant-Microbe Interactions, Seattle, Washington, USA, 11-16 July 1992 p. S13.
Drummond, M., Whitty, P. & Wooton, J. 1986 Sequence and domain relationships of ntrC and nifA from Klebsiella pneumoniae: homologies to other regulatory proteins. EMBO ]oumal 5, 441-447.
Dudley, M.E., Jacobs, T.W. & Long, S.R. 1987 Microscopic studies of cell divisions induced in alfalfa roots by Rhiwbium meliloti. Planta 171, 28+301.
Dusha, I., Bakos, A., Kondorosi, A., De Bruijn, F. & Shell, J. 1989 The Rhizobium meliloti early nodulation genes (nodABC) are nitrogen regulated: isolation of a mutant strain with efficient nodulation capacity on alfalfa in the presence of ammonium. Molecular and General Genetics 219, 89-96.
Dylan, T., Ielpi, L., Stanfield, S., Kashyap, L., Douglas, C., Yanosfsky, M., Nester, E., Helinski, D.R. & Ditta, G. 1986 Rhiwbium meliloti genes required for nodule development are related to chromosomal virulence genes in Agrobacterium tumefaciens. Proceedings of the National Academy of Sciences of the United States of America 83, 4403-4407.
Earl, C.D., Ronson, C.W. & Ausubel, F.M. 1987 Genetic and structural analysis of the Rhizobium meliloti fixA, fixB, frxC, and fxX genes. Journal of Bacferiology 169, 1127-1136.
Economou, A. & Downie, J.A. 1992 The nodulation of legumes by rhizobia. In Nitrogen Fixation and its Research in China, ed Hong, G.-F. pp. 315-339. Berlin and Heidelberg: Springer- Verlag.
Evans, LJ. SK Downie, J.A. 1986 The nod1 gene product of Rhizobium leguminosarum is closely related to ATP-binding bacterial
Bassam, B.J., Djorjevic, M.A., Redmond, J.W., Batley, M. & Rolfe, B.G. 1988 Identification of a nodD-dependent locus in the Rhiwbium strain NGR234 activated by phenolic factors secreted by soybeans and other legumes. Molecular Plant-Microbe Interactions I, 161-168.
Batut, J., Daveran-Mingot, M.L., David, M., Jacobs, J., Gamerone, A.M. & Kahn, D. 1989 FixK a gene homologous with fnr and crp from Escherichia coli, regulates nitrogen fixation genes both positively and negatively in Rhizobium meliloti. EMBO Journal 8, 1279-1286.
Batut, J., Santero, E. & Kustu, S. 1991 In vitro activity of the nitrogen fixation regulatory protein FixJ from R. meliloti. ]ournal of Bacferiology 173, 5914-5917.
Bhuvaneswari, T.V., Turgeon, G. & Bauer, W.D. 1980 Early stages in the infection of soybean (Glycine max L Merr) by Rhizobium
japonicum. 1. Localization of infective root cells. Plant Physiology 66, 1027-1031.
Better, M., Lewis, B., Corbin, D., Ditta, G. & Helinski, D.R. 1985 Structural relationships among Rhiwbium meliloti symbiotic promoters. Cell 35, 479-485.
Bibb, MJ., Biro, S., Motamedi, H., Collins, J.F. & Hutchinson, C.R. 1989 Analysis of the nucleotide sequence of the Streptomyces glaucescens tml genes provides key information about enzymology of poliketide antibiotic biosynthesis. EMBO ]ournal 8, 2727-2736.
Birkenhead, K., Nooman, B., Reville, W.J., Boesten, B., Manian, S.S. & O’Gara, F. 1990 Carbon utilization and regulation of nitrogen fixation genes in Rhizobium melilofi. Molecular Plant-Microbe lnteracfions 3, 167-173.
Bloemberg, G.V., Lugtenberg, B.J.J. & Spaink, H.P. 1992 In vitro transacetylating activity of the nodL protein of Rhizobium leguminosarum bv. viciae. In 6th International Symposium on the Molecular Genetics of Plant-Microbe Interactions, Seattle, Wash- ington, USA. 11-16 ]uly 1992.
Buck, M. & Cannon, W. 1987 Frameshifts close to the Klebsiella pnetrmoniae promoter prevent multicopy inhibition by hybrid nigh plasmids. Molecular and General Genetics 207, 492-498.
Bulawa, C.E. & Wasco, W. 1991 Chitin and nodulation. Nature 353, 710.
Caetano-Anolles, G., Crist-Estes, D.K. & Bauer, W.D. 1988 Chemotaxis of Rhiwbium meliloti to the plant flavone luteolin requires functional nodulation genes. ]oumal of Bacteriology 170, 3164-3169.
Canter-Cremers, H.C.J., Spaink, H.P., Wijfjes, A.H.M., Pees, E., Wijffelman, C.A., Okker, R.J.H. & Lubtenberg, B.J.J. 1989 Additional nodulation genes on the Sym plasmid of Rhiwbium leguminosarum biovar viciae. Plant Molecular Biology 13, 163-174.
Cervantes, E., Sharma, S.B., Maillet, F., Vasse, J., Truchet, G. & Rosenberg, C 1989 The Rhiwbium meliloti host range nodQ gene encodes a protein which shares homology with translation elongation and initiation factors. Molecular Microbiology 3, 745-755.
Colonna-Romano, S., Arnold, S., Schluter, A., Boistard, P. & Puhler, A. 1990 An Fnr-like protein encoded in Rhiwbium leguminosarum biovar viciae shows structural and functional homology to Rhizobium meliloti FixK. Molecular and General Genetics 223, 138-147.
David, M., Daveran, M.-L., Batut, J., Dedieu, A., Domergue, O., Ghai, J., Hertig, C., Boistard, P. & Kahn, D. 1988 Cascade regulation of nif gene expression in Rhiwbium meliloti. Cell 54, 671-683.
Davis, E.O. & Johnston, A.W.B. 1990 Regulatory functions of the three nodD genes of Rhizobium leguminosarum bv. phaseoli. Molecular Microbiology 3, 1567-1577.
Worki ]oumal of Microbiology and Biotechnology, Vol 9, 1993 451

M. Megias, ].L. Folch and C. Sousa
transport proteins; nucleotide sequence analysis of the nod1 and nod] genes. Gene 43, 95-101.
Firmin, J.L., Wilson, K.E., Rossen, L. & Johnston, A.W.B. 1986 Flavonoid activation of nodulation genes in Rhizubium reversed by other compounds present in plants. Nature 324, 90-92.
Fischer, R.F. & Long, S.R. 1992 Rhizobium-plant signal exchange. Nature 357, 655-660.
Folch, J.L., Spaink, H.P., Sousa, C., Quinto, C. & Megias, M. 1992 Rhizobirtm tropici CIAT899 nodulation genes and signals. In 6th International Symposium on the Molecular Genetics of Plant-- Microbe Interactions, Seattle, Washington, USA, II-16 July, 1992 p. 67.
Geelen, D., Goormachtig, S., Van Den Eede, G., Van Montagu, M. & Holster, M. 1992 Organization and functional analysis of the nod locus I genes from Azorhitobium caulinodans strain ORS571. In 6th international Symposium on the Molecular Genetics of Plant-Microbe Interactions, Seattle, Washington, USA, 11-16 July, 1992, p. 68.
Geiger, O., Spaink, H.P. & Lugtenberg, B.J.J. 1992 Phospholipids of Rhizobium contain nodE-determined highly unsaturated fatty acid moieties. In 6th international Symposium on the Molecular Genetics of Plant-Microbe Interactions, Seattle, Washington, USA, 11-16 July, 1992, p. 69.
Gilles-Gonzalez, M.A., f)‘ltta, G.S. & Helinski, D.R. 1991 A haemoprotein with kinase activity encoded by the oxygen sensor of Rhizobium meliloti. Nature 350, 170-172.
Giitz, R., Limmer, N., Ober, K. & S&nit, R. 1982 Motility and chemotaxis in two strains of Rhizobium with complex flagella. Journal of General Microbiology 128, 789-798.
Gubler, M. h Hennecke, H. 1986 @A, B and C genes are essential for symbiotic and free-living, microaerobic nitrogen fixation. European Journal of Biochemistry 146, 193-199.
Gussin, G.N., Ronson, C.W. & Ausubel, F.M. 1986 Regulation of nitrogen fixation genes. Annual Review of Genetics 20,567-591.
Gyiirgypal, Z, Iyer, N. & Kondorosi, A. 1988 Three regulatory nodD alleles of diverged flavonoid-specificity are involved in host-dependent nodulation by Rhizobium meliloti. Molecular and General Genetics 212, 85-92.
Hennecke, H., Alvarez-Morales, A., Betancourt-Alvarez, M., Ebeling, E., Filser, M., Fischer, H.-M., Gubler, M., Hahn, M., KaIusa, K., Lamb, J.W., Meyer, L., Regensburger, B., Studer, D. & Weber, J. 1985 Organization and regulation of symbiotic nitrogen fixation genes from Brudyrhizobium japonicum. In Nitrogen Fixation Research Progress, eds Evans, H.J., Bottomley, P.J. & Newton, W.E. pp. 157-163. The Hague: Nihjoff/Junk.
Holsters, M., Geelen, D., Goethals, K., Van Montagu, M., Geremia, R., Prom& J.C. & Mergaert, P. 1992 Nod factor production by Azorhizobium caulinodans strain ORS571. In Nau Horizons in Nitrogen Fixation, eds Palacios, R., Mora, J. & Newton, W.E. pp. 191-196. Dordrecht : Kluwer Academic.
Hontelez, J.G.J., Mol. P., Van Dun, C., Schetgens, R., Van Kammen, A. & Van Den Bos, R.C. 1984 Expression of sym-plasmid genes in bacteroids of Rhizubium leguminosarum. In Advances in Nitrogen Fixation Research, eds Veeger, C. & Newton, W.E. p. 686. The Hague: Nihjoff/Junk.
Horvath, B., Bachem, C.W.B., Schell, J. & Kondorosi, A. 1987 Host specific regulation of nodulation genes in Rhizobium is mediated by a plant signal, interacting with the nodD gene product. EMBO ]ournal6, 841-848.
HuaIa, E, & Ausubel, F.M. 1989 The central domain of Rhizobium meliloti NifA is sufficient to activate transcription from the R. meliloti nigh promoter. ]oumal of Bacteriology 171, 3354-3365.
Johnston, A.W.B., Beynon, J.L., Buchanan-Wollaston, A.V., Setchell, S.M., Hirsch, P.R. & Beringer, J.E. 1978 High frequency transfer
of nodulating ability between strains and species of Rhizobium. Nature 276, 634-636.
Kahn, D., David, M., Domergue, O., Daveran, M.-L., Ghai, J., Hirsch, P.R. & Batut, J. 1989 Rhizobium meliloti firGH1 sequence predicts involvement of a specific cation pump in symbiotic nitrogen fixation. ]oumal of Bacteriology 171, 929-939.
Kaminski, P.A. & Elmerich, C. 1991 Involvement of fixL] in the regulation of nitrogen fixation in Azorhizobium caulinodans. Molecular Microbiology 5, 665-673.
Kondorosi, E., Banfalvi, Z. & Kondorosi, A. 1984 Physical and genetic analysis of a symbiotic region of Rhizobium meliloti: identification of nodulation genes. Molecular and General Genetics 193, 445-452.
Kondorosi, E., Gyuris, J., Schmidt, J., John, M., Duda, E., Hoffman, B., Schell, J. & Kondorosi, A. 1989 Positive and negative control of nod gene expression in Rhizubium meliloti is required for optimal nodulation. EMBO ]ournal 8, 1331-1340.
Kustu, S., Weiss, D.S., Klose, K., Porter, S., North, A. & Wedel, A. 1992 Prokaryotic enhancers and enhancer binding proteins. In 6th International Symposium on the Molecular Genetics of Plant-Microbe Interactions, Seattle, Washington USA, 11-16 July, 1992, p. 520.
Lerouge, P., Roche, P., Faucher, C., Maillet, F., Truchet, G., Prome, J.C. & Denarie, J. 1990 Symbiotic host-specificity of Rhizobium meliloti is determined by sulphated and acylated glucosamine oligosaccharide signal. Nature 344, 78X-784.
Long, S.R. 1989 Rhizobium-legume nodulation: life together in the underground. Cell 56, 203-214.
Long, S.R. 1992 Expression and action of Rhizobium nod genes: recent results. In 6th Infernational Symposium on the Molecular Genetics of Plant-Microbe Interactions, Seattle, Washington, USA, 11-16 July 1992, p. Sll.
Maillet, F., Dhbelle, F. & D&a&, J. 1990 Role of the nodD and SyrM genes in the activation of the regulatory gene nodD3 and of the common and host-specific nod genes of Rhizobium meliloti. Molecular Microbiology 4, 1975-1984.
Mergaert, P., Van Montagu, M., Prome, J.C. & Holsters 1992 The Nod factors of Azorhizobium cuulinodans strain ORS571. In Nao Horizons in Nitrogen Fixation, eds Palacios, R., Mora, J. & Newton, W.E. pp. 191-196. Dordrecht: Kluwer Academic.
Nap, J.-P. & Bisseling, T. 1990 Developmental biology of a plan-prokaryote symbiosis: the legume root nodule. Science 250, 948-954.
Nooman, B., Motherway, M. & O’Gara, F. 1992 Ammonia regulation of the Rhizobium meliloti nitrogenase structural and regulatory genes under free-living conditions-involvement of the fiL gene product. Molecular and General Genetics 234, 423-428.
Norel, F., Kush, A., Denefle, P., Charpin, N. & Elmerich, C. 1984 Nitrogen fixation in a tropical Rhizobium associated with Sesbania rostrata. In Advances in Nitrogen Fixation Research, eds Veeger, C. & Newton, W.E. p. 694. The Hague: Nihjoff/Junk.
Nuti, M.P., Lepidi, A.A., Prakash, R.K., Schilperoort, R.A. & Cannon, F.C. 1979 Evidence for nitrogen fixation genes on indigenous Rhizobium plasmids. Nature 282, 533-535.
Peters, N.K. & Long, S.R. 1988 Alfalfa root exudate and compounds which promote or inhibit induction of Rhizobium meliloti nodulation genes. Plant Physiology 88, 396-400.
Redmond, J.W., Batley, M., Djordjevic, M.A., Innes, R.W., Kuempel, P.L. & Rolfe, B.G. 1986 Flavones induce expression of nodulation genes Rhizobium. Nature 323, 632-635.
Ridge, R.W. & Rolfe, B.G. 1986 Rhizobium sp. degradation of legume root hair cell wall at the site of infection thread origin. Applied and Environmental Minobiology 50, 717-750.
452 World]oumalofh4icmbiology and Biotechnology, Vol 9. 1993

Regulation of symbiotic nitrogen fixation
De Sande, K., Van Spronsen, P., Wijfjes, A.H.M. & Lugtenberg, B.J.J. 1992 Rhizobial lipo-oligosaccharide signals: their biosynth- esis and their role in the plant. In Molecular Genetics of Plant Microbe Interactions, eds Nester, E.W. & Verma D.P.S. pp. 151-162. Dordrecht: Kluwer Academic.
Spaink, H.P., Okker, R.J.H., Wijffelman, C.A., Tak, T., Goosen-De Roo, L., Pees, E., Van Brussel, A.A.N. & Lugtenberg, B.J.J. 1989b Symbiotic properties of rhizobia containing a flavonoid independent hybrid nodD product. Journal of Bacteriology 171, 4045-4053.
Spaink, H.P., Sheeley, D.M., Van Brussel, A.A.N., Glushkai, J., York, W.S., Tak, T., Geiger, O., Kennedy, E.P., Reinhold, V.N. & Lugtenberg, B.J.J. 1991 A novel highly unsaturated fatty acid moiety of lipo-oligosaccharide signals determines host specificity of Rhizobium. Nature 354, 125-130.
Spaink, H.P., Winman, J., Djordjevic, M.A., Wijffelman, CA., Okker, R.J.H. & Lugtenberg, B.J.J. 1989a Genetic analysis and cellular localization of the Rhizobium host-specificity-determin- ing NodE protein. EMBO ]ournal 8, 2811-2818.
Stock, J.B., Ninfa, A.J. & Stock, A.M. 1989 Protein phosphorylation and regulation of adaptive responses in bacteria. Microbiological Reviews 53, 45&490.
Szeto, W.W., Nixon, B.T., Ronson, C.W. & Ausubel, F.M. 1987 Identification and characterization of the Rhizobium melilofi nfr gene: R. melilofi has separate regulatory pathways for activation of nitrogen fixation genes in tree-living and symbiotic cells. ]ournaf of Bacferiology 169, 1423-1432.
Szeto, W.W., Zimmerman, J.L., Sundaresan, V. & Ausubel, F.M. 1984 A Rhizobium mehlofi symbiotic regulatory gene. Cell 36, 1035-1043.
Toro, N. & Olivares, J. 1986 Analysis of R. melilofi sym mutants obtained by heat treatment. Applied nnd Environmental Microbiology 5 1, 1148-1150.
Van Brussel, A.N.N., Recourt, K., Pees, E., Spaink, H.P., Tak, Y., Wijffelman, C.A., Kijne, J.W. & Lugtenberg, B.J.J. 1990 A biovar-specific signal of Rhizobium leguminosarum bv. viciae induces increased nodulation gene-inducing activity in root exudate of Vicia safiva subsp. n&u. ]ournal of Bacteriology 172, 5394-5401.
VandenBosch, K.A., Bradley, D.J., Knox, J.P., Perotto, S., Butcher, G.W. & Brewin, NJ. 1989 Common components of the infection thread matrix and the intercellular space identified by immunocytochemical analysis of pea nodules and uninfected roots. EMBO Journal 8, 335-342.
Van Den Eede, G., Dreyfus, B., Goethals, K., Montagu, V.M. & Holsters, M. 1987 Identification and cloning of nodulation genes from stem-nodulating bacterium ORS571. Molecular and General Genefics 206, 291-299.
Vazquez, M., Davalos, A., De Las Peiias, A., Sanchez, F. & Quinto, C. 1991 Novel organization of the common nodulation genes in Rhizobium leguminosarum bv. phaseoli strains. ]ournal of Bacteriology 173, 125&1258.
Vargas, C., Martinez, L.J., Megias, M. & Quinto, C. 1990 Identification and cloning of nodulation genes and host specificity determinants of the broad host range Rhizobium leguminosarum biovar phaseoli strain CIAT899. Molecular Microbiology 4, 1899-1910.
Vazquez, M., Santana, O., Villalobos, M.A. & Quinto, C. 1992 Similarity of the nod1 and nod] products with proteins from gram negative bacteria. Organization and function of nodulation genes related sequences in Rhizobium leguminosarum bv. phaseoh strain CE3. In 6fh Infernafional Symposium on the Molecular Genefics of Plant-Microbe Interactions, Seattle, Washington, USA, 11-16 July 1992, p. 140.
Rodriguez-Quiriones, F., Banfalvi, Z., Murphy, P. & Kondorosi, A. 1987 Interspecies homology of nodulation genes in Rhizobium. Plant Molecular Biology 8, 61-67.
Rodriguez-Quiiiones, F., Femandez-Burriel, M., Banfalvi, Z., Megias, M. & Kondorosi, A. 1989 Identification of a conserved, reiterated DNA region that influences the efficiency of nodulation in strain RS1051. Molecular Plunf-Microbe Inferucfions 2, 75-83.
Ronson, C.W., Nixon, B.T. & Ausubel, F.M. 1987 Conserved domains in bacterial regulatory proteins that respond to environmental stimuli. Cell 49, 579-581.
Rossen, L., Shearman, C.A., Johnston, A.W.B. & Downie, J.A. 1985 The nodD gene of Rhizobium leguminosarum is autoregulatory and in the presence of plant exudate induces the nodABC genes. EMBO ]ournaf 4, 3369-3373.
Ruvkun, G.B. & Ausubel, F.M. 1980 Interspecies homology of nitrogenase genes. Proceedings of the National Academy of Sciences of the Unifed States of America. 77, 191-195.
Sanjuan, J., Carlson, R.W., Spaink, H.P., Bhat, U.R., Barbour, W.M., Glushka, J. & Stacey, G. 1992 A 20methylfucose moiety is present in the lipo-oligosaccharide nodulation signal of Brudyrhizobium japonicum. Proceedings of the National Academy of Sciences of the United Stafes of America. 89, 8789-8793.
Schlaman, H.R.M., Okker, R.J.H. & Lugtenberg, B.J.J. 1992 Regulation of nodulation gene expression by NodD in rhizobia. ]ournal of Bacteriology 174, 5 177-5 182.
Schmidt, J., Wingender, R., John, M., Wieneke, U. & Schell, J. 1988 Rhizobium meliloti nodA and nodB genes are involved in generating compounds that stimulate mitosis of plant cells. Proceedings of the National Academy of Sciences of the United Sfafes of America. 85, 8578-8582.
Schofield, P.R. & Watson, J.M. 1986 DNA sequence of Rhizobium trifohi nodulation genes reveals a reiterated and potentially regulatory sequence proceeding nodAB and nodF. Nucleic Acids Research 14, 2891-2903.
Schultze, M., Quiclet-Sire, B., Kondorosi, E., Virelizier, H., Glushka, J.N., Endre, G., Gero, S.D. & Kondorosi, A. 1992 Rhizobium melilofi produces a family of sulphated lipo-oligosaccharides exhibiting different degrees of plant host specificity. Proceedings of the National Academy of Sciences of the United States of America, 89, 192-196.
Schwedock, J. & Long, S.R. 1990 ATP sulphurylase activity of the nodP and nodQ gene products of Rhizobium melilofi. Nature 348, 644-647.
Shearman, C.A., Rossen, L., Johnston, A.W.B. & Downie, J.A. 1986 The Rhizobium leguminosurum nodulation gene nodF encodes a polypeptide similar to acyl-carrier protein and is regulated by nodD plus a factor of pea root exudates. EMBO journal 5, 647-652.
Smit, G., Kijne, J.W. & Lugtenberg, B.J.J. 1987 Involvement of both cellulose fibrils and a Ca “+-de P endent adhesin in the attachment of Rhizobium leguminosarum to pea root hair tips. Journal of Bacteriology 169, 4294-4301.
Sheldon, P.S., Kelwick, R.G.O., Sidebottom, C., Smith, C.G. & Slabas, A.R. 1990 3-Oxoacyl-(acyl-carrier protein) reductase from avocado (Persea americana) fruit mesocarp. Biochemistry 272, 713-720.
Sousa, C., Folch, J., Boloix, P., Megias, M., Nava, N. & Quinto, C. 1993 A Rhizobium fropici DNA region carrying the amino terminal-half of a nodD gene and a nod-box like sequence is responsible for host range extension. Molecular Microbiology, in press.
Spaink, H.P., Aarts, A., Bloemberg, G.V., Folch, J., Geiger, O., Schalman, H.R.M., Thomas-Oates, J.E., Van Brussel, A.A.N., Van
World Jouml of Microbiology and Biotechnology, Vol 9. 1993 453

M. Megi’as, J.L. Folch and C. Sousa
Virts, E.L., Stanfield, S.W., Helinski, D.R. & Ditta, G.S. 1988 Common regulatory elements control symbiotic and micro- aerobic induction of nifA in Rhizobium meliloti. Proceedings of the National Academy of Sciences of the United States of America. 85, 3062-3065.
Yao, P.Y. & Vincent, J.M. 1969 Host specificity in the root hair “curling factor” of Rhizobium spp. Australian Journal of Biological Sciences 22, 413-423.
Zaat, S.A.J., Schripsema, J., Wijffelman, C.A., Van Brussel, A.A.N. & Lugtenberg, B.J.J. 1989 Analysis of the major inducers of the R&&urn nodA promoter from Viciu sutiva root exudates and their activity with different nodD genes. Plant Molecular Biology 13, 175-188.
Young, J.P.W. & Johnston, A.W.B. 1989 The evolution of specificity in the legume-R&&urn symbiosis. Tree 4,34X-349. (Accepted 5 April 1993)
454 World Journal of Microbiology ad Biotechnology, Vol 9. 1993
Copyright © 2022 FDOKUMEN




















