
Assessment of Suitability of Two Serotype A Candidate Vaccine Strains for Inclusion In FMD Vaccine...
8
Assessment of suitability of two serotype A candidate vaccine strains for inclusion in FMD vaccine in India Jajati Keshari Mohapatra a, * , Divakar Hemadri a , T.V.S. Rao b , B.P. Sreenivasa b , Saravanan Subramaniam a , Aniket Sanyal a , Tamilselvan Ramasamy Periyasamy a , Nawal Kishor Singh a , Bramhadev Pattnaik a , Ramamurthy Venkataramanan b a Project Directorate on Foot-and-Mouth Disease, Indian Veterinary Research Institute Campus, Mukteswar-Kumaon, Nainital 263138, Uttarakhand, India b Indian Veterinary Research Institute, Bangalore Campus, Hebbal, Bangalore 560024, India Received 26 November 2007; received in revised form 25 February 2008; accepted 28 February 2008 Abstract The recent type A foot and mouth disease virus field isolates recovered in India are shown to be antigenically quite divergent from the in-use vaccine strain (IND 17/82), warranting the selection of a suitable vaccine strain which can cover this diversity in antigenic spectrum. In earlier studies employing neutralization test with anti-146S rabbit sera raised against eight candidate vaccine strains, IND 81/00 and IND 40/00 belonging to genotype VII were found to offer the best antigenic coverage. In order to assess the credibility of IND 81/00 and IND 40/00 as vaccine strains, 17 recent isolates received during 2005–2006 and representative isolates from older genotypes were subjected to two-dimensional micro-neutralization assay using bovine convalescent serum (against IND 81/00 and IND 40/00) and bovine vaccinate serum (against IND 40/00). From the results it is evident that both the isolates IND 81/00 (antigenic relationship ‘r-value’ >0.40 with 86% of isolates) and IND 40/00 (‘r-value’ >0.40 with 78% of isolates) show nearly equal antigenic relatedness with the recent field viruses and hence both of these are effective vaccine candidates in present context. Though very limited in its extent, these useful data obtained with antisera raised in homologous host system are logical extension of the on going quest for the appropriate vaccine strain and circumvents species disparities in the immune recognition of epitopes. # 2008 Elsevier B.V. All rights reserved. Keywords: FMD serotype A; Vaccine strain selection 1. Introduction India is endemic for foot and mouth disease (FMD) and vaccination has been the sole means to save the susceptible livestock from this scourge. Of the three serotypes (O, A and Asia 1) prevalent in India, type A www.elsevier.com/locate/vetmic Available online at www.sciencedirect.com Veterinary Microbiology 131 (2008) 65–72 * Corresponding author. Tel.: +91 5942 286004; fax: +91 5942 286307. E-mail address: [email protected] (J.K. Mohapatra). 0378-1135/$ – see front matter # 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.vetmic.2008.02.020
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Assessment of Suitability of Two Serotype A Candidate Vaccine Strains for Inclusion In FMD Vaccine...

Assessment of suitability of two serotype A candidate vaccine
strains for inclusion in FMD vaccine in India
Jajati Keshari Mohapatra a,*, Divakar Hemadri a, T.V.S. Rao b, B.P. Sreenivasa b,Saravanan Subramaniam a, Aniket Sanyal a, Tamilselvan Ramasamy Periyasamy a,
Nawal Kishor Singh a, Bramhadev Pattnaik a, Ramamurthy Venkataramanan b
a Project Directorate on Foot-and-Mouth Disease, Indian Veterinary Research Institute Campus,
Mukteswar-Kumaon, Nainital 263138, Uttarakhand, Indiab Indian Veterinary Research Institute, Bangalore Campus, Hebbal, Bangalore 560024, India
Received 26 November 2007; received in revised form 25 February 2008; accepted 28 February 2008
Abstract
The recent type A foot and mouth disease virus field isolates recovered in India are shown to be antigenically quite divergent
from the in-use vaccine strain (IND 17/82), warranting the selection of a suitable vaccine strain which can cover this diversity in
antigenic spectrum. In earlier studies employing neutralization test with anti-146S rabbit sera raised against eight candidate
vaccine strains, IND 81/00 and IND 40/00 belonging to genotype VII were found to offer the best antigenic coverage. In order to
assess the credibility of IND 81/00 and IND 40/00 as vaccine strains, 17 recent isolates received during 2005–2006 and
representative isolates from older genotypes were subjected to two-dimensional micro-neutralization assay using bovine
convalescent serum (against IND 81/00 and IND 40/00) and bovine vaccinate serum (against IND 40/00). From the results it is
evident that both the isolates IND 81/00 (antigenic relationship ‘r-value’ >0.40 with 86% of isolates) and IND 40/00 (‘r-value’
>0.40 with 78% of isolates) show nearly equal antigenic relatedness with the recent field viruses and hence both of these are
effective vaccine candidates in present context. Though very limited in its extent, these useful data obtained with antisera raised
in homologous host system are logical extension of the on going quest for the appropriate vaccine strain and circumvents species
disparities in the immune recognition of epitopes.
# 2008 Elsevier B.V. All rights reserved.
Keywords: FMD serotype A; Vaccine strain selection
www.elsevier.com/locate/vetmic
Available online at www.sciencedirect.com
Veterinary Microbiology 131 (2008) 65–72
* Corresponding author. Tel.: +91 5942 286004;
fax: +91 5942 286307.
E-mail address: [email protected] (J.K. Mohapatra).
0378-1135/$ – see front matter # 2008 Elsevier B.V. All rights reserved
doi:10.1016/j.vetmic.2008.02.020
1. Introduction
India is endemic for foot and mouth disease (FMD)
and vaccination has been the sole means to save the
susceptible livestock from this scourge. Of the three
serotypes (O, A and Asia 1) prevalent in India, type A
.

J.K. Mohapatra et al. / Veterinary Microbiology 131 (2008) 65–7266
is antigenically and genetically most heterogeneous.
Four out of 10 global genotypes (Tosh et al., 2002) in
type A have been reported from India and since 2001,
genotype VII has out competed other genotypes and
has been behind all the outbreaks of recent years.
FMD virus is well known for its antigenic volatility
and infamous for giving rise to antigenically and
genetically divergent new strains with time leading to
poor antigenic coverage by in-use vaccine strains
(Brooksby, 1982). This notoriety on part of FMD
virus warrants a constant and elaborate antigenic and
genetic analysis of field viruses in relation to the
vaccine strain for confirming the appropriateness of
existing vaccine strain as well as for selection of new
vaccine strains, if situation demands. Preliminary
serological studies involving type A isolates collected
over a period of 9 years (1993–2001) using two-
dimensional micro-neutralization assay (2D-MNT)
indicated a poor intergenotypic antigenic coverage
among genotype VI and VII isolates (Mittal et al.,
2005; Tosh et al., 2003). To worsen the situation, the
Indian vaccine strain (IND 17/82) belongs to
genotype VI, whereas all the recent outbreaks are
attributed to genotype VII. In such an alarming
scenario, a need for a better vaccine strain that could
offer antigenic coverage to isolates from both the
genotypes was felt and subsequently a search for a
potential type A vaccine strain was initiated (Jangra
et al., 2005). In that study, 39 type A isolates collected
between 1999 and 2003 were subjected to one-way
antigenic analysis against two vaccine strains, viz.
IND 17/77 (genotype IV), IND 17/82 (genotype VI)
and a panel of eight candidate vaccine strains: four
field isolates each representing genotype VI (IND
233/99, IND 237/99, IND 258/99 and IND 68/01) and
genotype VII (IND 6/00, IND 40/00, IND 81/00 and
IND 173/00). Although intergenotypic antigenic
coverage offered by all the candidate vaccine strains
was poor, IND 81/00 and IND 40/00 emerged as better
candidates than all others. However, this extensive
antigenic analysis for selection of vaccine candidates
and their relative efficacy was carried out using anti-
146S rabbit sera. So a logical extension to this work
would be to assess the credibility of IND 81/00 and
IND 40/00 as candidate vaccine strains in homo-
logous target host system as obvious disparities in
epitope recognition pattern by the immune system
existing across species.
In this study, we report the antigenic analysis of
type A FMD field viruses isolated during 2005–2006
and a few selected strains of older genotypes (VI and
IV) in relation to the previously chosen better
candidate vaccine strains, IND 81/00 and IND 40/
00 (both genotype VII) using bovine convalescent sera
(BCS) and bovine vaccinal sera (BVS). Besides, the
study also provides performance of anti-146S rabbit
sera vis-a-vis bovine convalescent serum raised
against IND 81/00 with the field isolates. Finally,
results of the 2D-MNT using anti-146S rabbit sera
against the candidate vaccine strains, IND 258/99
(genotype VI) and the current vaccine strain, IND 17/
82 is also included for comparison purpose.
2. Materials and methods
2.1. Viruses
Seventeen type A FMDV field isolates recovered
during 2005–2006, current vaccine strain IND 17/82,
candidate vaccine strains IND 40/00, IND 81/00, IND
258/99 and a few representative isolates from
genotype IV and VI were used for antigenic analysis.
The history of FMDV field isolates is detailed in
Table 1. The virus isolates obtained from national
repository of FMDV maintained at Project Directorate
on FMD (PD on FMD) were passaged 5–6 times in
BHK-21 cell culture.
2.2. Animal sera
Hyperimmune anti-146S rabbit sera against binary
ethyleneimine (BEI) inactivated cell culture purified
virus particles (IND 258/99, IND 17/82 and IND 81/
00) was obtained from PD on FMD and were produced
as described previously (Tosh et al., 2003). For raising
BVS, IND 40/00 virus was grown in BKH-21 Razi cell
suspension culture and was subsequently purified, BEI
inactivated and oil adjuvanted. Bovines were inocu-
lated with this vaccine at a dose of 8 mg of antigen.
The serum from three vaccinated animals was
collected 21 days postvaccination and pooled. BCS
was raised by injecting intradermolingually 2 � 107
TCID50 dose of virus (IND 81/00 and IND 40/00) in
6–12 months old bovine calf. Serum was collected 21

J.K. Mohapatra et al. / Veterinary Microbiology 131 (2008) 65–72 67
Table 1
History of type A FMD virus field isolates used in this study
PD on FMD designation of virus isolate Date of collection Place of outbreak Host species
IND 17/77 1977 Tamilnadu Bovine
IND 490/97 (17/82)a 1982 West Bengal Bovine
IND 233/99 1999 Hisar, Haryana Bovine
IND 258/99b 12 February 1999 Bhavnagar, Gujarat Bovine
IND 40/00b 29 December 1999 Bangalore, Karnataka Bovine
IND 81/00b 29 January 2000 Bangalore, Karnataka Buffalo
IND 447/05 2005 Guwahati, Assam Not available
IND 22/06 11 November 2005 Kamrup, Assam Bovine
IND 23/06 12 November 2005 Kamrup, Assam Bovine
IND 26/00 2006 Kuchch, Gujarat Bovine
IND 27/06 2006 Kuchch, Gujarat Bovine
IND 28/06 2006 Kuchch, Gujarat Bovine
IND 43/06 15 February 2006 Jammu, J&K Bovine
IND 50/06 22 December 2005 Bangalore Bovine
IND 59/06 19 January 2006 Bangalore Bovine
IND 88/06 9 December 2005 Jabalpur, MP Bovine
IND 97/06 16 February 2006 Gwalior, MP Bovine
IND 99/06 6 March 2006 Madhya Pradesh Bovine
IND 100/06 6 March 2006 Gwalior, MP Bovine
IND 101/06 6 March 2006 Jabalpur, MP Bovine
IND 106/06 2006 Kuchcha, Gujarat Bovine
IND 109/06 26 November 2006 Madhya Pradesh Bovine
IND 152/06 17 March 2006 West Bengal Bovine
a Current vaccine strain.b Candidate vaccine strains.
days postinoculation and pooled from two calves to
avoid obvious individual bias in immune response.
2.3. Neutralization test
In order to estimate the antigenic relationship of the
field isolates, 2D-MNT was performed as per standard
method (Rweyemamu et al., 1978a). In brief, on the
same 96-well plate, seven 10-fold serial dilution of
virus (from 10�1 to 10�7) without serum in duplicate
wells horizontally was tested for determining virus
titre in terms of TCID50 and these dilutions were also
crosschecked by mixing and incubating with five 2-
fold serial dilution of serum (from 1:32 to 1:512) for
determining the dilution of serum that neutralizes 100
TCID50 virus. BHK 21 cells were used as indicator
system in neutralization test and the in vitro titres of
isolates varied from 106 to 108 TCID50/ml. Each test
was repeated thrice on 3 different days and the
averages were considered to avoid manual errors. On
every testing day, along with the isolates the
homologous virus was also tested to avoid any bias.
The end point titre of the serum was calculated as the
reciprocal of the last dilution of serum that neutralizes
100 TCID50 in 50% of the wells. The one-way
antigenic relationship (r-value) was calculated as the
ratio between heterologous and homologous serum
titre (Table 2). The results were interpreted based on
the criteria set by Samuel et al. (1990): an r-value
range of 0.40–1.00 indicates that existing vaccine
strain provide enough protection; while in the range of
0.20–0.39 indicates a need for more potent vaccine.
Conversely, r-value below 0.20 demands for a new
vaccine strain.
2.4. Genetic analysis
For phylogenetic analysis, nucleotide (nt) sequence
of ID region retrieved from local database of PD on
FMD and published previously (Tosh et al., 2002;
Jangra et al., 2005; Mittal et al., 2005; Mohapatra
et al., 2007) were used. Phylogenetic comparison was
performed using MEGA 3.1 software (Kumar et al.,
2004). The Tamura and Nei (1993) model of
J.K. Mohapatra et al. / Veterinary Microbiology 131 (2008) 65–7268
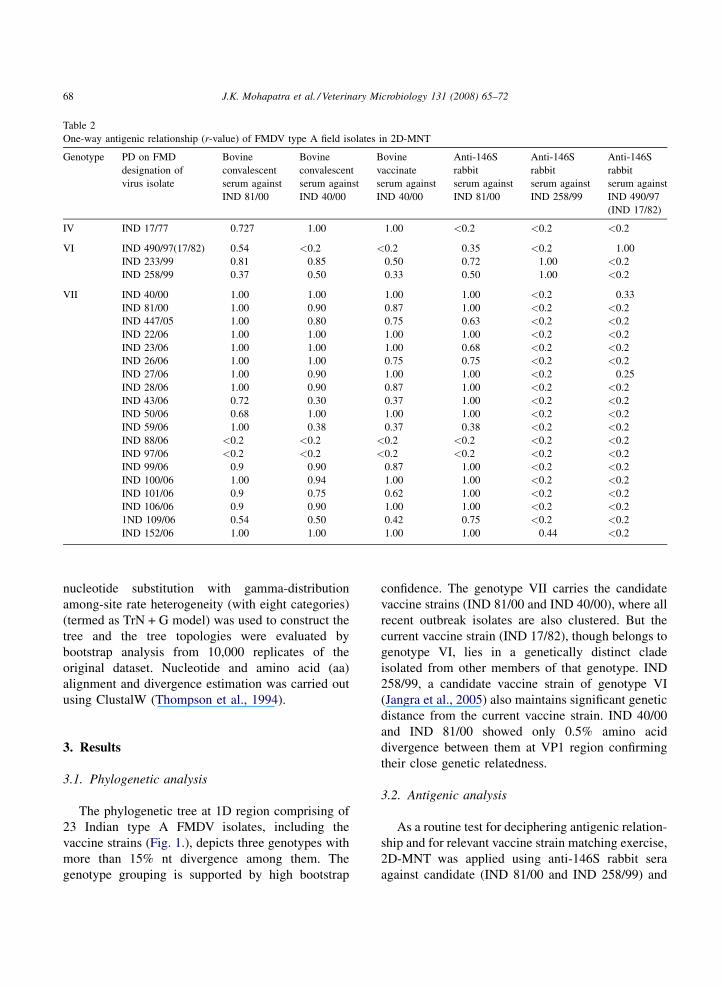
Table 2
One-way antigenic relationship (r-value) of FMDV type A field isolates in 2D-MNT
Genotype PD on FMD
designation of
virus isolate
Bovine
convalescent
serum against
IND 81/00
Bovine
convalescent
serum against
IND 40/00
Bovine
vaccinate
serum against
IND 40/00
Anti-146S
rabbit
serum against
IND 81/00
Anti-146S
rabbit
serum against
IND 258/99
Anti-146S
rabbit
serum against
IND 490/97
(IND 17/82)
IV IND 17/77 0.727 1.00 1.00 <0.2 <0.2 <0.2
VI IND 490/97(17/82) 0.54 <0.2 <0.2 0.35 <0.2 1.00
IND 233/99 0.81 0.85 0.50 0.72 1.00 <0.2
IND 258/99 0.37 0.50 0.33 0.50 1.00 <0.2
VII IND 40/00 1.00 1.00 1.00 1.00 <0.2 0.33
IND 81/00 1.00 0.90 0.87 1.00 <0.2 <0.2
IND 447/05 1.00 0.80 0.75 0.63 <0.2 <0.2
IND 22/06 1.00 1.00 1.00 1.00 <0.2 <0.2
IND 23/06 1.00 1.00 1.00 0.68 <0.2 <0.2
IND 26/06 1.00 1.00 0.75 0.75 <0.2 <0.2
IND 27/06 1.00 0.90 1.00 1.00 <0.2 0.25
IND 28/06 1.00 0.90 0.87 1.00 <0.2 <0.2
IND 43/06 0.72 0.30 0.37 1.00 <0.2 <0.2
IND 50/06 0.68 1.00 1.00 1.00 <0.2 <0.2
IND 59/06 1.00 0.38 0.37 0.38 <0.2 <0.2
IND 88/06 <0.2 <0.2 <0.2 <0.2 <0.2 <0.2
IND 97/06 <0.2 <0.2 <0.2 <0.2 <0.2 <0.2
IND 99/06 0.9 0.90 0.87 1.00 <0.2 <0.2
IND 100/06 1.00 0.94 1.00 1.00 <0.2 <0.2
IND 101/06 0.9 0.75 0.62 1.00 <0.2 <0.2
IND 106/06 0.9 0.90 1.00 1.00 <0.2 <0.2
1ND 109/06 0.54 0.50 0.42 0.75 <0.2 <0.2
IND 152/06 1.00 1.00 1.00 1.00 0.44 <0.2
nucleotide substitution with gamma-distribution
among-site rate heterogeneity (with eight categories)
(termed as TrN + G model) was used to construct the
tree and the tree topologies were evaluated by
bootstrap analysis from 10,000 replicates of the
original dataset. Nucleotide and amino acid (aa)
alignment and divergence estimation was carried out
using ClustalW (Thompson et al., 1994).
3. Results
3.1. Phylogenetic analysis
The phylogenetic tree at 1D region comprising of
23 Indian type A FMDV isolates, including the
vaccine strains (Fig. 1.), depicts three genotypes with
more than 15% nt divergence among them. The
genotype grouping is supported by high bootstrap
confidence. The genotype VII carries the candidate
vaccine strains (IND 81/00 and IND 40/00), where all
recent outbreak isolates are also clustered. But the
current vaccine strain (IND 17/82), though belongs to
genotype VI, lies in a genetically distinct clade
isolated from other members of that genotype. IND
258/99, a candidate vaccine strain of genotype VI
(Jangra et al., 2005) also maintains significant genetic
distance from the current vaccine strain. IND 40/00
and IND 81/00 showed only 0.5% amino acid
divergence between them at VP1 region confirming
their close genetic relatedness.
3.2. Antigenic analysis
As a routine test for deciphering antigenic relation-
ship and for relevant vaccine strain matching exercise,
2D-MNT was applied using anti-146S rabbit sera
against candidate (IND 81/00 and IND 258/99) and

J.K. Mohapatra et al. / Veterinary Microbiology 131 (2008) 65–72 69
Fig. 1. Neighbour joining tree at 1D region showing the phyloge-
netic relationship and genotype classification of FMDV type A
isolates. Bootstrap support value above 70% are shown. Current
vaccine strain is shaded in a box. Candidate vaccine strains and
genotype VI representative isolate are underlined.
in-use vaccine strain (IND 17/82). One-way antigenic
relationship (r-value) of type A isolates against these
strains representing two genotypes are shown in
Table 2. Out of 22 field isolates analyzed, only two
isolates showed an r-value in the range of 0.20–0.39
and rest of them had an r-value of <0.20 with the
current vaccine strain (IND 17/82) indicating its poor
antigenic relationship with recent field isolates
(Fig. 2). Similarly with IND 258/99 representing
genotype VI, only two isolates revealed an r-value of
>0.4. Whilst 77% (n = 17) and 9% (n = 2) of isolates
demonstrated an r-value in the range of 0.40–1.00 and
0.20–0.39, respectively with IND 81/00 anti-146S sera
confirming its wider antigenic relatedness with the
recent isolates.
In order to have a comprehensive antigenic
comparison of genotype VII candidates (IND 81/00
and IND 40/00) in homologous system, serum raised
in target host species for FMD, i.e. BCS (IND 81/00
and IND 40/00) and BVS (IND 40/00) were analyzed
for their neutralizing efficacy for recent isolates and
isolates from other genotypes. Here 20 out of the 22
tested isolates showed an r-value more than 0.2 with
IND 81/00 BCS while 19 of the 22 isolates showed an
r-value in the same range with IND 40/00 BCS and
also BVS (Fig. 2 and Table 2). Thus, from this result it
is evident that both IND 40/00 and IND 81/00 had
almost equal r-value relationships with the recent field
isolates tested.
Interestingly, isolates IND 88/06 and IND 97/06
revealed an r-value of <0.20 with all the sera tested
including IND 81/00 (both rabbit and bovine sera) and
IND 40/00 (both BCS and BVS) indicative of their
exceptional antigenic divergence. At the same time,
IND 109/06 and IND 152/06 which cluster with IND
88/06 and IND 97/06, respectively in the phylogenetic
tree have shown r-value of >0.4 with both IND 81/00
and IND 40/00 sera.
4. Discussion
In an FMD endemic country where vaccination is
the most important means of disease control, it is
imperative that a constant watch on the antigenic
makeup of circulating field viruses and their antigenic
and genetic relationship with the in-use vaccine strains
be maintained for updating vaccine strains. For this, in
vitro neutralization test is considered to be more
relevant to in vivo protection than other measures of
virus–antibody interactions, although non-neutraliz-
ing antibodies may also play critical role in terms of
protection (Dunn et al., 1998; McCullough et al.,
1992).
Here, we present antigenic analysis of 17 recent
field viruses isolated between 2005 and 2006 with
current vaccine strain (IND 17/82) and candidate
strains from two genotypes (IND 81/00 and IND 40/00
from genotype VII; IND 258/99 from genotype VI) in
2D-MNT as a continuation of the search for a better
relevant vaccine strain. As observed earlier (Jangra
et al., 2005), the circulating field viruses are also
antigenically unrelated with the current vaccine strain,
making it irrelevant for inclusion in the vaccine
formulation any more. This is evident from the ‘r-
value’ pattern observed for the recent isolates with
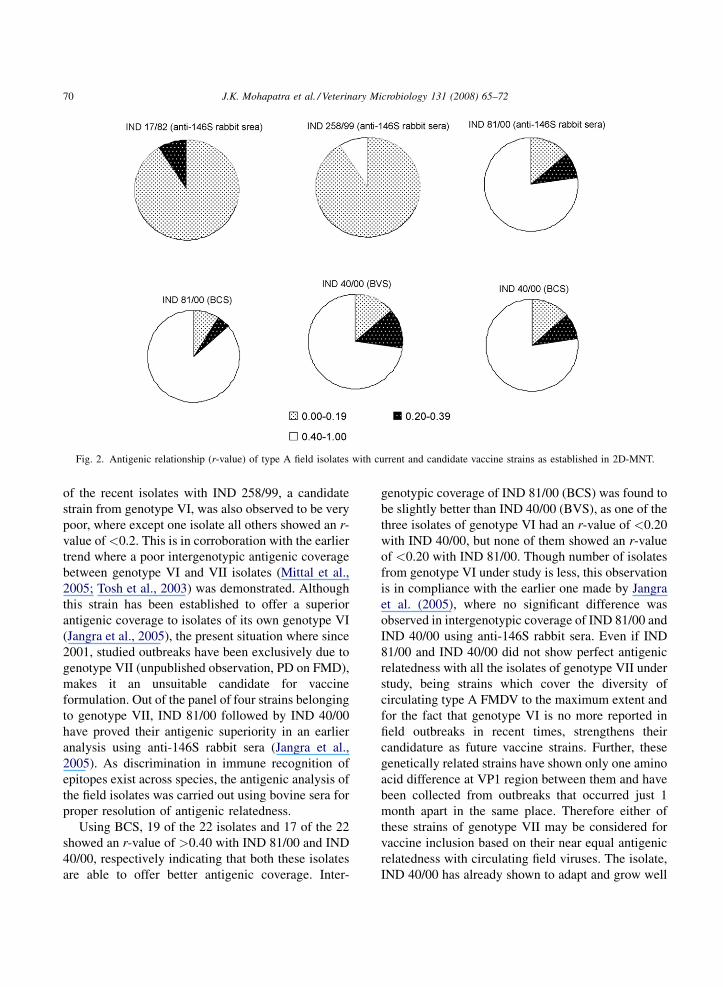
IND 17/82 (Fig. 2), where 94% of the isolates tested
showed an r-value of <0.2. The antigenic relatedness
J.K. Mohapatra et al. / Veterinary Microbiology 131 (2008) 65–7270
Fig. 2. Antigenic relationship (r-value) of type A field isolates with current and candidate vaccine strains as established in 2D-MNT.
of the recent isolates with IND 258/99, a candidate
strain from genotype VI, was also observed to be very
poor, where except one isolate all others showed an r-
value of <0.2. This is in corroboration with the earlier
trend where a poor intergenotypic antigenic coverage
between genotype VI and VII isolates (Mittal et al.,
2005; Tosh et al., 2003) was demonstrated. Although
this strain has been established to offer a superior
antigenic coverage to isolates of its own genotype VI
(Jangra et al., 2005), the present situation where since
2001, studied outbreaks have been exclusively due to
genotype VII (unpublished observation, PD on FMD),
makes it an unsuitable candidate for vaccine
formulation. Out of the panel of four strains belonging
to genotype VII, IND 81/00 followed by IND 40/00
have proved their antigenic superiority in an earlier
analysis using anti-146S rabbit sera (Jangra et al.,
2005). As discrimination in immune recognition of
epitopes exist across species, the antigenic analysis of
the field isolates was carried out using bovine sera for
proper resolution of antigenic relatedness.
Using BCS, 19 of the 22 isolates and 17 of the 22
showed an r-value of >0.40 with IND 81/00 and IND
40/00, respectively indicating that both these isolates
are able to offer better antigenic coverage. Inter-
genotypic coverage of IND 81/00 (BCS) was found to
be slightly better than IND 40/00 (BVS), as one of the
three isolates of genotype VI had an r-value of <0.20
with IND 40/00, but none of them showed an r-value
of <0.20 with IND 81/00. Though number of isolates
from genotype VI under study is less, this observation
is in compliance with the earlier one made by Jangra
et al. (2005), where no significant difference was
observed in intergenotypic coverage of IND 81/00 and
IND 40/00 using anti-146S rabbit sera. Even if IND
81/00 and IND 40/00 did not show perfect antigenic
relatedness with all the isolates of genotype VII under
study, being strains which cover the diversity of
circulating type A FMDV to the maximum extent and
for the fact that genotype VI is no more reported in
field outbreaks in recent times, strengthens their
candidature as future vaccine strains. Further, these
genetically related strains have shown only one amino
acid difference at VP1 region between them and have
been collected from outbreaks that occurred just 1
month apart in the same place. Therefore either of
these strains of genotype VII may be considered for
vaccine inclusion based on their near equal antigenic
relatedness with circulating field viruses. The isolate,
IND 40/00 has already shown to adapt and grow well

J.K. Mohapatra et al. / Veterinary Microbiology 131 (2008) 65–72 71
in suspension cells for vaccine production (results not
shown) but the growth characteristics of IND 81/00
remains to be assessed. Besides, anti-146S rabbit
serum against IND 81/00 revealed an r-value of <0.2
with genotype IV isolate (IND 17/77), whereas BCS
showed an r-value of >0.40. This again supports the
species-specific immune recognition pattern and the
rationale behind determining antigenic relationship
using serum raised in target host system for flawless
interpretation.
Usually vaccine matching tests employ BVS raised
against vaccine strains (Paton et al., 2005), partly
because its production does not require handling of
live virus outside the laboratory and partly because of
the variability in result in terms of virus specificity
reported for convalescent sera (Rweyemamu et al.,
1978b). But in this study, the serum against IND 81/00
is a convalescent serum whereas against IND 40/00
both convalescent and vaccinal sera were applied. As
it is established that BEI inactivation can compromise
the integrity of certain proteins of the virus
(Kasermann et al., 2001), which may lead to loss of
certain immunologically critical neutralizing epitopes,
comparison of results obtained with BVS and that with
BCS might be questionable. Notably, comparison of
‘r’ values obtained from convalescent and vaccinal
sera against IND 40/00 shows similar performances by
both these sera indicating use of convalescent sera did
not affect the final inference of the test results in our
study. Still to elucidate how IND 81/00 behaves after
BEI inactivation and for a better comparison with IND
40/00, BVS against IND 81/00 remains to be produced
and tested with all circulating field viruses.
To sum up, in compliance with the earlier findings,
both the isolates IND 40/00 and IND 81/00 are found
equally better candidate vaccine strains for replace-
ment of the existing vaccine strain. The current
vaccine strain, IND 17/82 and the candidate vaccine
strain from genotype VI (IND 258/99) continues to
offer poor intergenotypic coverage to recent isolates
belonging to genotype VII. Hence in present scenario,
where genotype VII has been exclusively associated
with all the field outbreaks in India since more than 6
years, IND 81/00 and IND 40/00 by virtue of their
superiority in their antigenic and genetic relatedness to
the circulating field isolates emerge to be the candidate
vaccine strains of choice. Not withstanding this fact,
complete genomic characterization of these two
strains and antigenic matching with more number of
circulating field viruses need to be undertaken.
Acknowledgements
We are thankful to Indian Council of Agricultural
Research for providing necessary facilities to carry out
this work. Technical assistance of Shri N.S. Singh and
Shri G. Singh, PD on FMD and Shri H.R. Narayana,
TO of FMD vaccine production at IVRI, Bangalore
campus is appreciated.
References
Brooksby, J.B., 1982. Portraits of viruses: foot-and-mouth disease
virus. Intervirology 18, 1–23.
Dunn, C.S., Samuel, A.R., Pullen, L.A., Anderson, J., 1998. The
biological relevance of virus neutralisation sites for virulence
and vaccine protection in the guinea pig model of foot-and-
mouth disease. Virology 247, 51–61.
Jangra, R.K., Tosh, C., Sanyal, A., Hemadri, D., Bandyopadhyay,
S.K., 2005. Antigenic and genetic analyses of foot and mouth
disease virus type A isolates for selection of candidate vaccine
strain reveals emergence of a variant virus that is responsible for
most recent outbreaks in India. Virus Res. 112, 52–59.
Kasermann, F., Wyss, K., Kempf, C., 2001. Virus inactivation and
protein modifications by ethyleneimines. Antivir. Res. 52, 33–
41.
Kumar, S., Tamura, K., Nei, M., 2004. MEGA3: integrated software
for molecular evolutionary genetics analysis and sequence
alignment. Briefings Bioinform. 5, 150–163.
McCullough, K.C., De Simone, F., Brocchi, E., Capucci, L.,
Crowther, J.R., Kihm, U., 1992. Protective immune response
against foot-and-mouth disease. J. Virol. 66, 1835–1840.
Mittal, M., Tosh, C., Hemadri, D., Sanyal, A., Bandyopadhyay, S.K.,
2005. Phylogeny, genome evolution and antigenic variability
among endemic foot and mouth disease virus type A isolates
from India. Arch. Virol. 150, 911–928.
Mohapatra, J.K., Saravanan, S., Tosh, C., Hemadri, D., Sanyal, A.,
Tamilselvan, R.P., Rasool, T.J., 2007. Genotype differentiating
RT-PCR and sandwich ELISA: handy tools in epidemiological
investigation of foot and mouth disease. J. Virol. Methods 143,
117–121.
Paton, D.J., Valarcher, J.-F., Bergmann, I., Matlho, O.G., Zakharov,
V.M., Palma, E.L., Thomson, G.R., 2005. Selection of foot and
mouth disease vaccine strains—a review. Rev. Sci. Techn. Off.
Int. Epiz. 24, 981–993.
Rweyemamu, M.M., Booth, J.C., Head, M., Pay, T.W.F., 1978a.
Microneutralization tests for serological typing and subtyping of
foot-and-mouth disease virus strains. J. Hyg. 81, 107–123.
Rweyemamu, M.M., Ouldridge, D.J., Head, M., Ferrari, R., 1978b.
The effect of antiserum quality on strain specificity assessment

J.K. Mohapatra et al. / Veterinary Microbiology 131 (2008) 65–7272
of foot and mouth disease virus by the neutralization reaction. J.
Biol. Stand. 12, 295–303.
Samuel, A.R., Ouldridge, E.J., Arrowsmith, A.E., Kitching, R.P.,
Knowles, N.J., 1990. Antigenic analysis of serotype O foot-and-
mouth disease virus isolates from the middle east, 1981–1988.
Vaccine 8, 390–396.
Tamura, K., Nei, M., 1993. Estimation of the number of nucleotide
substitutions in the control region of mitochondrial DNA
in humans and chimpanzees. Mol. Biol. Evol. 10, 512–
526.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTAL W:
improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, positions-specific gap
penalties and weight matrix choice. Nucleic Acids Res. 22,
4673–4680.
Tosh, C., Sanyal, A., Hemadri, D., Venkataramanan, R., 2002.
Phylogenetic analysis of serotype A foot-and-mouth disease
virus isolated in India between 1977 and 2000. Arch. Virol.
147, 493–513.
Tosh, C., Hemadri, D., Sanyal, A., Bandyopadhyay, S.K., 2003.
Genetic and antigenic analysis of two recently circulating
genotypes of type A foot and mouth disease virus in India:
evidence for positive selection in the capsid coding genes. Arch.
Virol. 148, 853–869.




















