
Arrested development in Fucus spiralis (Phaeophyceae) germlings exposed to copper
10
This article was downloaded by: [94.228.204.10] On: 21 March 2014, At: 12:36 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK European Journal of Phycology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tejp20 Arrested development in Fucus spiralis (Phaeophyceae) germlings exposed to copper Peter Bond a , Murray Brown a , Roy Moate a , Martha Gledhill b , Stephen Hill b & Malcolm Nimmo b a Department of Biological Sciences, University of Plymouth, Plymouth PL4 8AA, UK b Department of Environmental Sciences, University of Plymouth, Plymouth PL4 8AA, UK Published online: 03 Jun 2010. To cite this article: Peter Bond , Murray Brown , Roy Moate , Martha Gledhill , Stephen Hill & Malcolm Nimmo (1999) Arrested development in Fucus spiralis (Phaeophyceae) germlings exposed to copper, European Journal of Phycology, 34:5, 513-521 To link to this article: http://dx.doi.org/10.1080/09541449910001718871 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Arrested development in Fucus spiralis (Phaeophyceae) germlings exposed to copper

This article was downloaded by: [94.228.204.10]On: 21 March 2014, At: 12:36Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
European Journal of PhycologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tejp20
Arrested development in Fucus spiralis (Phaeophyceae)germlings exposed to copperPeter Bond a , Murray Brown a , Roy Moate a , Martha Gledhill b , Stephen Hill b & MalcolmNimmo ba Department of Biological Sciences, University of Plymouth, Plymouth PL4 8AA, UKb Department of Environmental Sciences, University of Plymouth, Plymouth PL4 8AA, UKPublished online: 03 Jun 2010.
To cite this article: Peter Bond , Murray Brown , Roy Moate , Martha Gledhill , Stephen Hill & Malcolm Nimmo (1999) Arresteddevelopment in Fucus spiralis (Phaeophyceae) germlings exposed to copper, European Journal of Phycology, 34:5, 513-521
To link to this article: http://dx.doi.org/10.1080/09541449910001718871
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Eur. J. Phycol. (1999), 34: 513–521. Printed in the United Kingdom 513
Arrested development in Fucus spiralis (Phaeophyceae)
germlings exposed to copper
PETER R. BOND1, MURRAY T. BROWN1, ROY M. MOATE1, MARTHA GLEDHILL2,STEPHEN J. HILL2 AND MALCOLM NIMMO2
"Department of Biological Sciences, University of Plymouth, Plymouth PL4 8AA, UK
#Department of Environmental Sciences, University of Plymouth, Plymouth PL4 8AA, UK
(Received 2 November 1998 ; accepted 16 June 1999)
Exposure of Fucus spiralis germlings to precise copper concentrations (0 to 844 nM Cu#+) in chemically defined medium demonstrated a
relationship between ultrastructural changes and growth retardation with increasing copper concentration. Electron-translucent vesicles,
present in ova, which normally disappear after fertilization, accumulated in germlings exposed to Cu#+ above 10±6 nM, suggesting that
copper may inhibit a metabolic pathway involved in cell wall formation which is initiated by fertilization. No membrane damage was
observed during the exposure period. During a post-exposure period in copper-free medium, recovery occurred (rhizoid extension, apical
hair formation) in germlings previously exposed to concentrations below 106 nM Cu#+ and electron-translucent vesicles became granular
and disappeared. It is proposed that the electron-translucent vesicles contain a cell wall precursor and that copper inhibits its incorporation
into the cell wall, preventing growth and development of the zygote.
Key words : copper, electron-translucent vesicles, Fucus spiralis, germlings, ultrastructure.
Introduction
Copper is an essential element for all organisms includingalgae, required in trace amounts for the functioning ofenzymes involved in oxygen chemistry and redoxreactions (Linder, 1991). However, at elevated concen-trations it becomes toxic, adversely affecting growth (e.g.Stro$ mgren, 1980 ; Munda & Hudnik, 1986), photosyn-thesis (e.g. Gupta & Arora, 1978), pigment synthesis (e.g.Pellegrini et al., 1993) and reproduction (Chung &Brinkhuis, 1986 ; Andersson & Kautsky, 1996). Increasedconcentrations of copper in the marine environment haveboth natural and anthropogenic origins, with mining,industrial discharges and pesticide applications beingmajor contamination sources (Nor, 1987).
The effects of copper on marine organisms have beenthe subject of intense research, much of which has centredon invertebrates. While algae, and in particular certainspecies of macroalgae, have been widely used as bio-monitors of metal pollution including copper, manyquestions concerning the processes of metal accumulation,toxicity and tolerance remain to be resolved. It is nowrecognized that total copper concentrations in seawaterdo not accurately represent the fraction bioavailable toalgae, since both inorganic and organic materials areknown to form complexes with the metal, and the
Correspondence to : Peter Bond, Plymouth Electron Microscopy Centre,
University of Plymouth, Drake Circus, Plymouth, Devon PL4 8AA, UK.
Tel}fax : 44 (0) 1752 233092. e-mail : pbond!plymouth.ac.uk
toxicities of the various chemical forms (species) differ(Gledhill et al., 1997). Copper toxicity can be directlyrelated to the free ion (Cu#+) concentrations and, by usingchemically defined growth media, it is possible to calculatethe available copper in solution thus allowing toxico-logical responses to be related directly to the bioavailablefraction. Moreover, there is a need to investigate growthresponses and physiological and cellular changes inducedby toxicants at concentrations representative of environ-mental conditions.
Typically, adult stages of macroalgae are used inbiomonitoring programmes, yet it is apparent from severalstudies (Stro$ mgren, 1980 ; Brinkhuis & Chung, 1986 ;Andersson & Kautsky, 1996) that the early developmentstages in a seaweed’s life history are more sensitive topollutants and thus can be used to investigate the effectsof short-term, low-level exposure. Fucoid germlings areprime examples : their ease of culture, rapid growth rateand quick response to toxicants make them convenientexperimental material for investigations into the mech-anisms of toxicity and tolerance.
The present study uses the chemically defined mediumAquil developed by Morel et al. (1979) to study the effectsof trace metal species on phytoplankton. Germlings of thebrown seaweed Fucus spiralis were exposed to precisecopper concentrations similar to those known to occur atpolluted sites in the UK (e.g. the Fal Estuary, Cornwall) toinvestigate the relationship between physiological(growth reduction), morphological and ultrastructuralchanges and increasing copper concentrations.
Dow
nloa
ded
by [
94.2
28.2
04.1
0] a
t 12:
36 2
1 M
arch
201
4

514P. R. Bond et al.
Materials and methods
Gamete}zygote release
Fucus spiralis L. is monoecious and fertilization normallytakes place within, or immediately after release of gametesfrom, the conceptacle and zygotes are released singly or,more often, in clumps associated with oogonia (Mu$ ller &Gassmann, 1985). To stimulate their release in thelaboratory, excised receptacles from fertile plants collectedfrom an unpolluted site at the mouth of the Erme estuaryin S. Devon, England were washed in distilled water for15 min, dried and chilled at 4 °C for several hours beforebeing incubated in growth medium in petri dishes at 15 °Cunder a light}dark regime of 16 h light}8 h dark and aphoton irradiance of 40–50 µmol m−# s−". Releasedzygotes were transferred into sterile polycarbonate petridishes containing 25 cm$ growth medium with totalcopper concentrations of 0, 50, 100, 200, 500, 1000 and2000 nM. Germlings were grown for a total of 10 daysunder the same conditions as described above. Mediumwas changed every 2 days. At the end of the exposureperiod, recovery of the germlings from copper exposurewas investigated over a 7 day period using copper-freemedium under the same culture conditions.
Growth media were based on Aquil (Morel et al., 1979)but without the addition of a chelating agent, as this hasbeen found to result in a small reduction in growth whencomparedwith equivalent unchelated total copper concen-trations (M. Gledhill et al., personal observation). The freecopper concentrations initially present in the growthmedia (Table 1) were calculated at pH 7±6, a pCO
#of
21±66 and a temperature of 15 °C using the computerprogram MINEQL (Environmental Research Software) ;however, it is likely that, as has been seen for germlings ofF. vesiculosus (M. Gledhill, personal observation), coppercomplexing material is released as the germlings grow,thereby decreasing the free copper concentration. Thisrelease of ligands was compensated for by changing thegrowth medium every 2 days. Concentrations of Cu#+ arereferred to throughout the paper, unless stated otherwise.
Growth measurements
The areas of newly released zygotes and 10-day-oldgermlings were measured using a Kyowa binocularmicroscope attached to a Cambridge InstrumentsQuantimet 570 image analyser via a Hitachi VK}C150EDcolour video camera. The mean areas of 10 germlings fromeach treatment were analysed using a one-way ANOVA
Table 1. Total copper concentration and calculated freely
available copper in culture media
Total Cu (nM) 50 100 200 500 1000 2000
Free Cu (nM) 21±1 42±2 84±4 211 422 844
and differences between means of the seven exposuretreatments were tested with Duncan’s multiple range testat the 95% confidence levels using StatGraphics.
Light microscopy
Germlings were pipetted from culture vessels onto glassmicroscope slides, covered with coverslips and examinedand photographed on an Olympus Vanox microscopeunder brightfield illumination.
Preparation for transmission electron microscopy
Germlings were fixed for 2 h in 2±5% glutaraldehyde in0±1 M phosphate buffer, pH 7±2, with 3±0% sodiumchloride added. Caffeine (1±0%) was added to the fixativeto stabilize physodes (Clayton & Beakes, 1983). Post-fixation was in buffered 1±0% osmium tetroxide for 1 h.Dehydration through an ethanol series (30%, 50%, 70%,90%, 100%; 15 min each) was followed by infiltrationwith Spurr’s resin for 14 days (changed every 2 days) priorto polymerization overnight at 60 °C. Sections were cuton a Reichert Ultracut ultramicrotome with a diamondknife, stained with uranyl acetate and lead citrate andexamined in JEOL 1200s and JEOL 2000FX electronmicroscopes at 80–100 kV.
Results
Exposure period
Germling growth, estimated by a change in area after 10days of exposure, was unaffected by Cu#+ concentrations
Fig. 1. Effect of copper on the growth of Fucus spiralis germlings.
Growth was measured as change in germling area after 10 days.
Error bars represent standard deviation and range test categories
are indicated. Columns labelled with the same letter do not differ
significantly at the 95% confidence level.
Dow
nloa
ded
by [
94.2
28.2
04.1
0] a
t 12:
36 2
1 M
arch
201
4

Fucus spiralis arrested development 515
Figs 2–7. Light micrographs of F. spiralis germlings exposed to copper. Fig. 2. Day 1, zygotes prior to formation of rhizoid initial. Fig. 3.
Ten-day-old germling, no copper exposure, showing extensive rhizoid network (r) and apical hairs (h). Fig. 4. Ten-day-old germling
exposed to 84±4 nM Cu#+, showing extensive rhizoids (r) but no apical hairs (h). Fig. 5. Ten-day-old germling exposed to 211 nM Cu#+.
The rhizoid system (r) is less well developed and no apical hairs are present. Fig. 6. Ten-day-old germling exposed to 422 nM Cu#+,
showing short, undeveloped rhizoids (r). Fig. 7. Ten-day-old germling exposed to 844 nM Cu#+, showing development arrested at the
rhizoid initial stage. Scale bars represent 100 µm.
below 84±4 nM (200 nM total Cu), but at concentrationsabove this, significant growth retardation occurred (Fig.1).
Germlings exposed to 21±1 nM Cu#+ and42±2 nM Cu#+ appeared to be unaffected after 10 days.Multiple rhizoids had formed, attaching germlings tothe culture vessels, juvenile thalli were well developedand apical hairs were present. Exposure to 84±4 nM Cu#+
reduced or inhibited apical hair development while rhizoidgrowth appeared unaffected (cf. Figs 3 and 4). Germlinggrowth was retarded at 211 nM Cu#+ ; rhizoids wereshorter, no apical hairs had formed and thalli were less welldeveloped (Fig. 5). Germlings exposed to 422 nM Cu#+
had no apical hairs, rhizoids were very short and thalliwere less elongated (Fig. 6). At a concentration of844 nM Cu#+, germling development was severelyaffected ; while rhizoid initials had formed on the majorityof germlings, thallus elongation had only just begun (Fig7).
Prior to release from the oogonium the mature,unfertilized ovum has no cell wall (Fig. 2). Most of the cell
volume is occupied by electron-opaque (densely staining)physodes and vesicles with electron-translucent contents(lightly staining) (Fig. 8). After fertilization, and releasefrom the conceptacle, these pale vesicles, which arepredominant in the eggs, disappear as the zygote matures(cf. Figs 8 and 9).
Transmission electron microscopic (TEM) observationsof germling ultrastructure complement the light micro-scopy observations and the growth data. Althoughgrowth was retarded in germlings exposed to844 nM Cu#+ for 10 days, there was no detectableultrastructural change in the membranes of the chloro-plasts, Golgi bodies, and mitochondria, and the singlemembrane surrounding the electron-translucent vesiclesappeared to be intact (Figs 10, 11).
At concentrations of 844, 422 and 211 nM Cu#+, pale,electron-translucent, vesicles (v) were present in themajority of cells, and occupied a large proportion of anycell profile (Figs 12–14). At 84±4 nM Cu#+ pale vesicleswere observed in some cells but they never occupied themajority of any cell profile (Fig. 15), and at lower
Dow
nloa
ded
by [
94.2
28.2
04.1
0] a
t 12:
36 2
1 M
arch
201
4

516P. R. Bond et al.
Figs 8–11. Transmission electron microscopy of F. spiralis germlings exposed to copper. Fig. 8. Ovum prior to fertilization and release
from oogonium, showing distribution of vesicles including physodes (p) and electron-translucent vesicles (v) ; c, chloroplast. Scale bar
represents 2 µm. Fig. 9. Thallus cell of 10-day-old germling grown in copper-free medium, showing physodes (p), chloroplasts (c) and
absence of electron-translucent vesicles ; w, cell wall. Scale bar represents 2 µm. Fig. 10. Detail from thallus cell of a 10-day-old germling
exposed to 422 nM Cu#+, showing undamaged Golgi bodies (g) and mitochondria (m). Scale bar represents 200 nm. Fig. 11. Detail from a
thallus cell of a 10-day-old germling exposed to 844 nM Cu#+, showing intact chloroplast (c) and mitochondrial (m) membranes and
extensive accumulation of electron-translucent vesicles (v). Scale bar represents 200 nm.
concentrations (42±2 nM and 21±1 nM) they were absent(Figs 16, 17).
Post-exposure period
When returned to copper-free medium after 10 days ofexposure, germlings previously exposed to Cu#+ concen-trations of 422 nM or less began to recover (cf. Figs 18–21and Figs 3–7). After 7 days of recovery in copper-freemedium, germlings previously exposed to Cu#+ at84±4 nM appeared to be fully recovered as extensiverhizoids were present and apical hairs had fully developed(Fig. 18). Germlings exposed to 211 nM Cu#+ showedsome recovery but no apical hairs had formed and theirrhizoid networks were shorter and more branched (cf. Figs18 and 19). After exposure to 422 nM Cu#+, recovery wasslower ; after 7 days, rhizoids were short and unbranchedbut the thallus had not begun to elongate (Fig. 20).
Growth of germlings previously exposed to 844 nM Cu#+
showed no signs of recovery during the 10 day post-exposure period (Fig. 21).
Ultrastructural changes during the post-exposure re-covery period began within 2 days, except in germlingsexposed to 844 nM Cu#+, in which no change was notedduring the 7 day recovery period (cf. Figs 22–24 and Figs12–17). Electron-translucent vesicles, which occupied themajority of the cell volume in germlings exposed to422 nM Cu#+ during exposure, were present in some cellsafter 2 days of recovery, but by day 7 physodes were thepredominant vesicles within the cytoplasm (Fig. 22).During the recovery period the contents of the electron-translucent vesicles became more granular and electron-opaque (Figs 22, 23). Within 7 days of transfer to copper-freemedium, electron-translucent vesicles had disappearedcompletely from germlings previously exposed to concen-trations of 211 nM Cu#+ and 84±4 nM Cu#+ (Fig. 24).
Dow
nloa
ded
by [
94.2
28.2
04.1
0] a
t 12:
36 2
1 M
arch
201
4

Fucus spiralis arrested development 517
Figs 12–17. Transmission electron microscopy of Fucus spiralis germlings exposed to copper. Fig. 12. Thallus cell of 10-day-old
germling exposed to 844 nM Cu#+. Electron-translucent vesicles (v) occupy the majority of the cell volume, with chloroplasts (c) and
physodes (p) restricted to the cell periphery. Fig. 13. Thallus cell of 10-day-old germling exposed to 422 nM Cu#+, showing physodes (p)
and chloroplasts (c) distributed in an extensive accumulation of electron-translucent vesicles (v). Fig. 14. Thallus cell of 10-day-old
germling exposed to 211 nM Cu#+, showing distribution of electron-translucent vesicles (v), chloroplasts (c) and physodes (p). Fig. 15.
Thallus cell of 10-day-old germling exposed to 84±4 nM Cu#+, showing abundant physodes (p) with electron-translucent vesicles (v)
occupying less than half the cell volume. Figs 16, 17. Thallus cell of 10-day-old germling exposed to 42±2 nM Cu#+ (Fig. 16) and
21±1 nM Cu#+ (Fig. 17). No electron-translucent vesicles (v) are present ; physodes (p) and chloroplasts (c) make up the majority of the cell
contents. N, nucleus ; w, cell wall. Scale bars represent 2 µm, except Fig. 17 where the scale bar represents 1 µm.
Discussion
The pattern of germination and development of fucoidzygotes is well documented (Brawley et al., 1976 ;Quatrano & Stevens, 1976 ; Evans et al., 1982). Cell wallmaterial is deposited after fertilization of the egg, and the
zygote then undergoes cell division, the basal cell formingan attachment rhizoid while the apical cell forms thethallus. As the germling continues to grow, furtherdifferentiation occurs with the development of a rhizoidalnetwork, elongation and thickening of the thallus andformation of apical hairs after several days of growth. It
Dow
nloa
ded
by [
94.2
28.2
04.1
0] a
t 12:
36 2
1 M
arch
201
4

518P. R. Bond et al.
Figs 18–21. Light microscopy of Fucus spiralis germlings after 7 days post-exposure recovery in copper-free culture medium. Fig. 18.
Germling previously exposed to 84±4 nM Cu#+ with extensive rhizoid network (r) and apical hairs (h) developing. Fig. 19. Germling
previously exposed to 211 nM Cu#+. No apical hairs have developed and rhizoids (r) are shorter and more branched than controls (Fig. 3).
Fig. 20. Germling previously exposed to 422 nM Cu#+, showing arrested thallus development, restricted rhizoid (r) growth and absence of
apical hairs. Fig. 21. Germling previously exposed to 844 nM Cu#+. Development has been halted by copper exposure and no recovery
has begun. Scale bars represent 100 µm.
was found in the present study that this sequence ofdevelopment was disrupted by exposure to copper.
Ten days of exposure to copper concentrations greaterthan 211 nM resulted in a significant reduction in growthrates of F. spiralis germlings compared with those growingin media containing copper at lower concentrations. Theseeffects on growth were accompanied by disruption of thetypical mode of morphological development, includingreduction in apical hair formation, rhizoid inhibition andretardation of thallus development.
On return to a copper-free medium for 7 days, allgermlings previously exposed to copper concentrations ashigh as 422 nM Cu#+ resumed growth and the normalcourse of development resumed. However, exposure toconcentrations greater than this resulted in germlingdevelopment being irreversibly inhibited even aftertransfer to the copper-free solution. Despite the observedalterations in the physiological (growth) and morpho-logical (development) responses of the germlings tocopper exposure, there was no detectable damage to thecell membranes. Chloroplast, mitochondrial, nuclear andGolgi body membranes all appeared to be intact, even at
the highest copper concentration used (844 nM Cu#+).This contrasts with the findings of Brinkhuis & Chung(1986) who describe chloroplast membrane swelling anddisruption, and separation of cell wall fibrils, in copper-exposed Laminaria saccharina sporophytes. However,these ultrastructural changes occurred only when copperwas added at 7500 nM, a concentration 3 times higherthan any used in the current study. As the presentinvestigation was more concerned with sublethal effectsof copper exposure, further studies at higher concen-trations would be required to evaluate the tolerance limitsof membrane integrity of Fucus germlings.
The information obtained from light microscopy andTEM in our study indicates that short-term exposure tocopper at concentrations less than 1000 nM total copperis not lethal to Fucus germlings. This finding has importantecological significance since many pollution incidents areacute, and therefore an ability to withstand short-termexposure to toxicants followed by the resumption ofnormal growth and development will enhance the like-lihood of survival.
While no damage to the ultrastructure of cells was
Dow
nloa
ded
by [
94.2
28.2
04.1
0] a
t 12:
36 2
1 M
arch
201
4
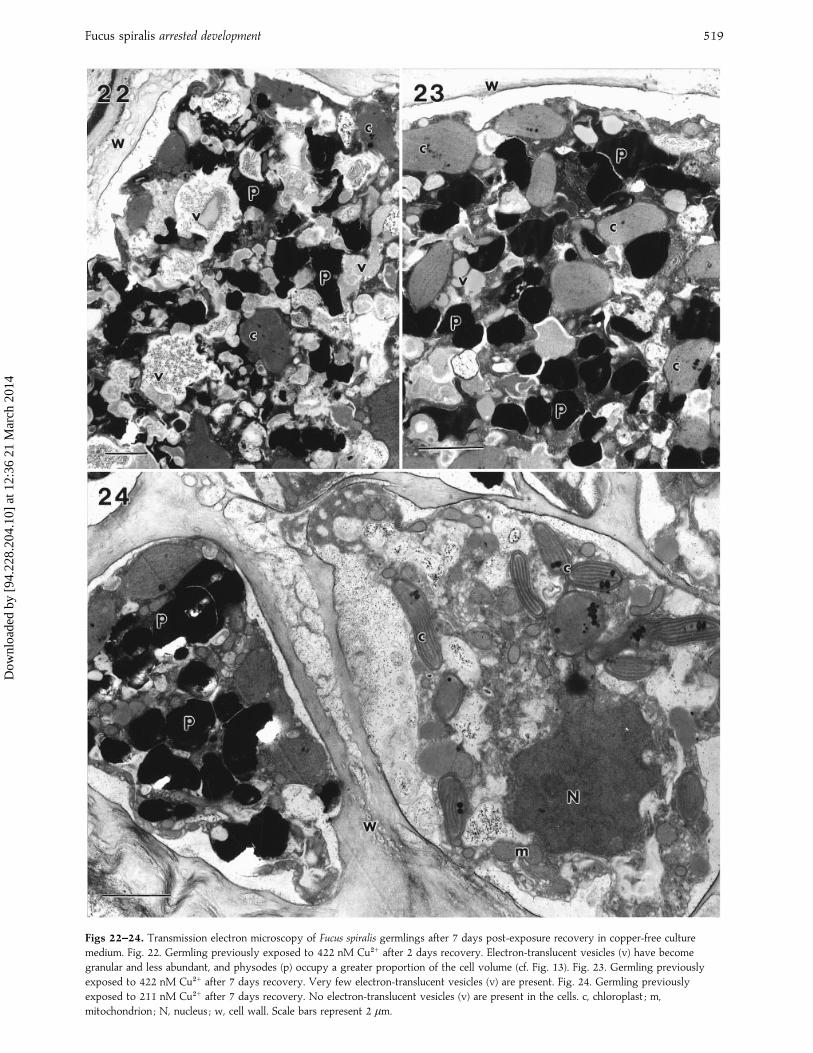
Fucus spiralis arrested development 519
Figs 22–24. Transmission electron microscopy of Fucus spiralis germlings after 7 days post-exposure recovery in copper-free culture
medium. Fig. 22. Germling previously exposed to 422 nM Cu#+ after 2 days recovery. Electron-translucent vesicles (v) have become
granular and less abundant, and physodes (p) occupy a greater proportion of the cell volume (cf. Fig. 13). Fig. 23. Germling previously
exposed to 422 nM Cu#+ after 7 days recovery. Very few electron-translucent vesicles (v) are present. Fig. 24. Germling previously
exposed to 211 nM Cu#+ after 7 days recovery. No electron-translucent vesicles (v) are present in the cells. c, chloroplast ; m,
mitochondrion ; N, nucleus ; w, cell wall. Scale bars represent 2 µm.
Dow
nloa
ded
by [
94.2
28.2
04.1
0] a
t 12:
36 2
1 M
arch
201
4

520P. R. Bond et al.
evident, even at the highest level of copper exposure,modifications to the normal course of ultrastructuraldevelopment were apparent when germlings were cul-tured in medium containing copper. The ultrastructure ofthe Fucus vesiculosus ovum and early development of thezygote have been described by Brawley et al. (1976). Priorto penetration of the sperm nucleus the egg is notsurrounded by a cell wall and the cytoplasm contains arange of membrane-bound bodies including electron-opaque physodes and pale, electron-translucent vesicles.They suggested that fertilization initiates transformationand utilization of the contents of these pale vesicles for cellwall deposition as the zygote begins to grow and divide,because during this phase of growth these vesicles becomegranular and eventually disappear.
Physodes and electron-translucent vesicles occupiedthe majority of the cytoplasm in F. spiralis ova anddisappeared after fertilization in algae cultured in copper-free medium. However, exposure to increasing copperconcentrations up to 844 nM Cu#+, whilst inhibitinggrowth and development, also resulted in the accumu-lation of electron-translucent vesicles in the cells of thegermlings.
During the recovery period, following removal ofcopper from the medium, these electron-translucentvesicles became granular and disappeared from germlingsexposed to copper below 106 nM. Exposure to copperthus appears to inhibit utilization of the material stored inthe vesicles but does not affect the metabolic processinvolved in their production. Germling growth dependson new cell wall formation, and if, as suggested byBrawley et al. (1976), the pale vesicles contain alginic acid,which is a major component of the cell walls of brownalgae, then inhibition of its mobilization and depositionwould restrict growth and impede normal development.During the recovery period the correlation between theresumption of growth of the germlings and disappearanceof the electron-translucent vesicles suggests that thesevesicles may contain a cell wall precursor and that copperions inhibit its mobilization or incorporation, thus pre-venting cell expansion and growth. The mechanism forthis may be inhibition by Cu#+ ions of enzyme activityinvolved in the mobilization and deposition of wallpolymers, but there is no immediate evidence for this andfurther study is needed.
However, it is known that Cu#+ binds to cell wallpolysaccharides and also to polyphenols, which are storedin physodes. Other ligand sites, for example cytoplasmicpolysaccharides and proteins, are occupied when thephysode binding sites are saturated (Smith et al., 1986 ;Karez & Pereira, 1995). This may explain the tolerance ofgermlings to low and medium concentrations of copperand the toxic effects at higher concentrations, and mightexplain the accumulation of carbohydrates in pale vesicles,because either enzyme hydrolysis is inactivated orpolysaccharide cross-linking (and thus cell wall deposition)is prevented because copper is bound to these reactionsites.
Zygotes and germlings are considered to be moresensitive than mature individuals to metal pollution(Brinkhuis & Chung, 1986 ; Andersson & Kautsky, 1996),and we have found that 3¬10$ nM total copper exposurewas needed to reduce the growth of apical segments ofmature thalli of F. spiralis, F. serratus and F. vesiculosus by50% with respect to controls (unpublished results). Incontrast, only 500 nM copper was required to reduce thegrowth of germlings by 50% in the present study. In thiscontext, Stro$ mgren (1980) found that growth of segmentsof mature individuals of F. spiralis, F. serratus, and F.vesiculosus was reduced to 50% of controls when exposedto total copper concentrations above 1200 nM for a 10day period.
The coincidence of growth inhibition with the ac-cumulation of electron-translucent vesicles in zygotes andgermlings of F. spiralis in response to copper exposure, andthe ability of germlings to recover from exposure, havenot been shown before in macroalgae, and further work toinvestigate copper localization within cells of exposed F.spiralis zygotes is presently being undertaken.
Acknowledgements
The authors would like to thank John Green for his helpfulcomments on the manuscript.
References
A, S. & K, L. (1996). Copper effects on reproductive stages
of Baltic Sea Fucus vesiculosus. Mar. Biol., 125 : 171–176.
B, S.H., W, R. & Q, R.S. (1976). Fine-structural
studies of the gametes and embryo of Fucus vesiculosus L. (Phaeophyta) II.
The cytoplasm of the egg and young zygote. J. Cell Sci., 29 : 255–271.
B, B.H. & C, I.K. (1986). The effects of copper on the fine
structure of the kelp Laminaria saccharina (L.) Lamour. Mar. Environ. Res.,
19 : 2205–223.
C, I.K. & B, B.H. (1986). Copper effects in early stages of the
kelp, Laminaria saccharina. Mar. Poll. Bull. 17 : 213–218.
C, M. & B, G.W. (1983). Effects of fixatives on the ultrastructure
of physodes in vegetative cells of Scytosiphon lomentaria
(Scytosiphonaceae, Phaeophyta). J. Phycol., 19 : 4–16.
E, L.V., C, J.A. & C, M.E. (1982). The biology and
biochemistry of reproduction and early development in Fucus. In Progress
in Phycological Research, vol. 2 (Round, F.E. and Chapman, D.J., Editors).
Elsevier Biomedical Press, Dordrecht.
G! , J.A., M, K.H. & C, A.R.O. (1982). Seasonal patterns of
growth and storage in Laminaria longicruris in relation to differing patterns
of availability of nitrogen in the water. Mar. Biol., 69 : 91–101.
G, M., N, M., H, S. & B, M. (1997). The toxicity of
copper species to marine algae, with particular reference to macroalgae. J.
Phycol., 33 : 2–11.
G, A.B. & A, A. (1978). Morphology and physiology of Lyngbya
nigra with reference to copper toxicity. Physiol. Plant., 44 : 215–220.
K, C.S. & P, R.C. (1995). Metal contents in polyphenolic fractions
extracted from the brown alga Padina gymnospora. Bot. Mar., 38 :
151–155.
L, O.M., P, A.M., V, M.T.S.D., G, C.A.R. &
S, R. (1996). Potential tolerance mechanisms of Procentrum micans
(Dinophyceae) to sublethal levels of copper. J. Phycol., 32 : 416–423.
L, M.C. (1991). Biochemistry of Copper. Plenum Press, New York.
Dow
nloa
ded
by [
94.2
28.2
04.1
0] a
t 12:
36 2
1 M
arch
201
4

Fucus spiralis arrested development 521
M, F.M.M., R, J.G., A, D.M. & G, R.R.L. (1979).
Aquil : a chemically defined phytoplankton culture medium for trace metal
studies. J. Phycol., 15 : 135–142.
M$ , D.G. & G, G. (1985). Sexual reproduction and the role of
sperm attractants in monoecious species of the brown algal order Fucales
(Fucus, Hesperophycus, Pelvetia and Pelvetiopsis). J. Plant Physiol., 118 :
401–408.
M, I.M. & H, V. (1986). Growth response of Fucus vesiculosus to
heavy metals, singly and dual combinations, as related to accumulation.
Bot. Mar., 29 : 401–412.
N, Y.M. (1987). Ecotoxicity of copper to aquatic biota : a review. Environ.
Res., 43 : 274–282.
P, M., L, A., S, M., P-T-L, R., V, R. &
P, L. (1993). Interactions between the toxicity of the heavy
metals cadmium, copper, zinc in combinations and the detoxifying role of
calcium in the brown alga Cytoseira barbata. J. Appl. Phycol., 5 : 351–361.
Q, R.S. & S, P.T. (1976). Cell wall assembly in Fucus zygotes.
I. Characterisation of the polysaccharide components. Plant Physiol., 58 :
224–231.
S, K.L., H, A.C. & H, J.L. (1986). The subcellular localisation
of absorbed copper in Fucus. Physiol. Plant., 66 : 692–698.
S$ , T. (1980). The effect of dissolved copper on the increase in
length of four species of intertidal fucoid algae. Mar. Environ. Res., 3 :
5–13.
W, R.S. (1991). Cadmium in seaweeds. PhD thesis. University of Otago,
Dunedin, New Zealand.
Dow
nloa
ded
by [
94.2
28.2
04.1
0] a
t 12:
36 2
1 M
arch
201
4




















