
All VH11 genes expressed in peritoneal lymphocytes encode anti-bromelain-treated mouse red blood...
7
Eur. J. Irnmunol. 1990. 20: 1583-1589 VHI 1 expression and anti-BrMRBC specificity 1583 Pascal Poncet, Franqois Huetz, Miguel-Angel Marcos and Luiz Andrade Laboratory of Immunobiology, Pasteur Institute, Paris All VHl1 genes expressed in peritoneal lymphocytes encode anti-bromelain-treated mouse red blood cell autoantibodies but other VH gene families contribute to this specificity* We have studied the relationship between B cell reactivity to bromelain-treated autologous mouse erythrocytes (BrMRBC) and expression of the VHll gene family in splenic, peritoneal and pleuropericardial cell populations from normal C57BL/6 mice. B lymphocytes producing antibodies to BrMRBC were selectively enriched or depleted from normal populations by rosette formation with BrMRBC, followed by centrifugation over density gradients. This selection method, based on the presence of functional receptors (membrane IgM), is harmless for the cells and allowed subsequent cloning in agar (colony-forming unit-B). The utilization of the 10 VH gene families was then scored in mRNA colony blot assays. The analysis of >650 anti-BrMRBC clones and >350 VH11-expressing colonies indicates that about half of those antibody reactivities are encoded by VHll genes. Furthermore, it appears that all VH11-expressing B cells in the peritoneal cavity produce anti-BrMRBC antibodies. 1 Introduction Complex mechanisms of selection exerted on a potential repertoire of immune receptors (TcR and Ig) lead to the expression of the available repertoire and of certain activated clones constituting the actual repertoire at a given moment. Autoreactive clones are active members in the establishment and the maintainance of this homeostasis, and are themselves submitted to selection [l]. Natural autoantibodies which lyse bromelain-treated syn- geneic mouse RBC (BrMRBC) constitute a prime example of selected autoreactivity. They are present in all mouse strains and have been described to be isotypically and idiotypically restricted. Their selection concerns their spe- cificity and also the localization of producing B cells (pleural cavity > peritoneal cavity >spleen). Other charac- teristics of these antibodies, such as late ontogenic devel- opment and incomplete overlap with Ly-1 B cells, have been reported (reviewed in [2]). Studies of hybridoma collections have correlated the anti-BrMRBC specificity with the expression of the VHll gene family [3-91. Uncertainty as to the representativity of hybridoma collections with respect to normal, unselected populations, as well as the relatively low numbers of clones analyzed by this approach, led us to study VH gene family usage with techniques which allowed the analysis of a high number of cells. Thus, mRNA colony blot assay was [I 82301 * This research was supported by grants from CEE, DRETand ARC. Correspondence: Pascal Poncet, Unit6 d'hmunobiologie, Institut Pasteur, 25-28, rue du Dr. Roux, F-75724 Paris Cedex 15, France Abbreviation used: BCP: B cell clonal precursor BrMRBC: Brornelain-treated mouse red blood cell HRBC: Horse red blood cell LDL: Low density lipoprotein R: Rosetting fraction NR: Nonrosetting fraction RFC: Rosette-forming cell performed in normal cell populations depleted or enriched in B cells able to specifically form rosettes with BrMRBC (rosette-forming cell; RFC). The RFC technique is a convenient method selecting cells on the basis of the functionality of the expressed receptor. This results presented here show that the selection also apply to the genetic determinism of anti-BrMRBC specif- icity since we observed a strong positive correlation between BrMRBC reactivity of lymphocytes existing in spleen, pleural and peritoneal cavities and the expression of VHll genes. More importantly, our data suggest that all VHll genes expressed in peritoneal B cells encode anti- BrMRBC antibodies. 2 Materials and methods 2.1 Animals and cells Inbred C57BL/6, raised in our facilities at the Pasteur Institute, were used between 8-10 weeks of age. Spleen cells were prepared by gentle teasing of the organs in cold BSS. In certain experiments, they were fractionated on a discontinuous gradient of Percoll(40%0 -100% ; Pharmacia, Uppsala, Sweden). Low-density (50% -60% interface) large lymphocytes were considered as activated cells, while high-density (60% -70% and 70% -80% interfaces) lym- phocytes were taken as resting cells [lo]. Peritoneal and pleuropericardial cells were obtained by injecting 5-10 ml of chilled BSS supplemented with 3% FCS in the respective cavities. All cell suspensions were washed twice before utilization. SRBC and horse RBC (HRBC) were used after three washes in saline.TNP was coupled to SRBC according to Weyer and Bussard [ll]. Bromelinization of MRBC was performed as described previously [12]. 2.2 RFC technique The different cells suspensions were adjusted to lo7 celldm1 with cold BSS supplemented with 3% FCS and 1110th 0 VCH Verlagsgcsellschaft rnbH, D-6940 Weinheim, 1990 0014-2980/90/0707-1583$03.50 + ,2510
Transcript of All VH11 genes expressed in peritoneal lymphocytes encode anti-bromelain-treated mouse red blood...

Eur. J. Irnmunol. 1990. 20: 1583-1589 VHI 1 expression and anti-BrMRBC specificity 1583
Pascal Poncet, Franqois Huetz, Miguel-Angel Marcos and Luiz Andrade
Laboratory of Immunobiology, Pasteur Institute, Paris
All V H l 1 genes expressed in peritoneal lymphocytes encode anti-bromelain-treated mouse red blood cell autoantibodies but other VH gene families contribute to this specificity*
We have studied the relationship between B cell reactivity to bromelain-treated autologous mouse erythrocytes (BrMRBC) and expression of the VHll gene family in splenic, peritoneal and pleuropericardial cell populations from normal C57BL/6 mice. B lymphocytes producing antibodies to BrMRBC were selectively enriched or depleted from normal populations by rosette formation with BrMRBC, followed by centrifugation over density gradients. This selection method, based on the presence of functional receptors (membrane IgM), is harmless for the cells and allowed subsequent cloning in agar (colony-forming unit-B). The utilization of the 10 VH gene families was then scored in mRNA colony blot assays. The analysis of >650 anti-BrMRBC clones and >350 VH11-expressing colonies indicates that about half of those antibody reactivities are encoded by VHll genes. Furthermore, it appears that all VH11-expressing B cells in the peritoneal cavity produce anti-BrMRBC antibodies.
1 Introduction
Complex mechanisms of selection exerted on a potential repertoire of immune receptors (TcR and Ig) lead to the expression of the available repertoire and of certain activated clones constituting the actual repertoire at a given moment. Autoreactive clones are active members in the establishment and the maintainance of this homeostasis, and are themselves submitted to selection [l].
Natural autoantibodies which lyse bromelain-treated syn- geneic mouse RBC (BrMRBC) constitute a prime example of selected autoreactivity. They are present in all mouse strains and have been described to be isotypically and idiotypically restricted. Their selection concerns their spe- cificity and also the localization of producing B cells (pleural cavity > peritoneal cavity >spleen). Other charac- teristics of these antibodies, such as late ontogenic devel- opment and incomplete overlap with Ly-1 B cells, have been reported (reviewed in [2]).
Studies of hybridoma collections have correlated the anti-BrMRBC specificity with the expression of the VHll gene family [3-91. Uncertainty as to the representativity of hybridoma collections with respect to normal, unselected populations, as well as the relatively low numbers of clones analyzed by this approach, led us to study VH gene family usage with techniques which allowed the analysis of a high number of cells. Thus, mRNA colony blot assay was
[I 82301
* This research was supported by grants from CEE, DRETand ARC.
Correspondence: Pascal Poncet, Unit6 d'hmunobiologie, Institut Pasteur, 25-28, rue du Dr. Roux, F-75724 Paris Cedex 15, France
Abbreviation used: BCP: B cell clonal precursor BrMRBC: Brornelain-treated mouse red blood cell HRBC: Horse red blood cell LDL: Low density lipoprotein R: Rosetting fraction NR: Nonrosetting fraction RFC: Rosette-forming cell
performed in normal cell populations depleted or enriched in B cells able to specifically form rosettes with BrMRBC (rosette-forming cell; RFC). The RFC technique is a convenient method selecting cells on the basis of the functionality of the expressed receptor.
This results presented here show that the selection also apply to the genetic determinism of anti-BrMRBC specif- icity since we observed a strong positive correlation between BrMRBC reactivity of lymphocytes existing in spleen, pleural and peritoneal cavities and the expression of VHll genes. More importantly, our data suggest that all VHll genes expressed in peritoneal B cells encode anti- BrMRBC antibodies.
2 Materials and methods
2.1 Animals and cells
Inbred C57BL/6, raised in our facilities at the Pasteur Institute, were used between 8-10 weeks of age. Spleen cells were prepared by gentle teasing of the organs in cold BSS. In certain experiments, they were fractionated on a discontinuous gradient of Percoll(40%0 -100% ; Pharmacia, Uppsala, Sweden). Low-density (50% -60% interface) large lymphocytes were considered as activated cells, while high-density (60% -70% and 70% -80% interfaces) lym- phocytes were taken as resting cells [lo]. Peritoneal and pleuropericardial cells were obtained by injecting 5-10 ml of chilled BSS supplemented with 3% FCS in the respective cavities. All cell suspensions were washed twice before utilization. SRBC and horse RBC (HRBC) were used after three washes in saline.TNP was coupled to SRBC according to Weyer and Bussard [ l l ] . Bromelinization of MRBC was performed as described previously [12].
2.2 RFC technique
The different cells suspensions were adjusted to lo7 celldm1 with cold BSS supplemented with 3% FCS and 1110th
0 VCH Verlagsgcsellschaft rnbH, D-6940 Weinheim, 1990 0014-2980/90/0707-1583$03.50 + ,2510

1584 l? Poncet, F. Huetz, M.-A. Marcos and L. Andrade Eur. J. Immunol. 1990. 20: 1583-1589
volume of a 10% BrMRBC suspension was then added. Cells were centrifuged 5 min (500 x g) and resuspended on a roller at 4°C for 15 min. Microscopic observation of a mixture allowed the determination of the percentages of FWC. Only rosettes showing > 4 erythrocytes per B cell were considered as positive and their number was expressed as percent of total lymphoid cells. Enrichment of RFC was performed on a discontinuous Percoll gradient composed of 2 ml of 70% solution for the lower layer and 2 ml of the 30% solution for the upper layer. Mixture of cells were gently overlaid on the top of the gradient (a maximum of 5 x lo6 cells). The tubes were then centrifuged 15 min (1500 x g) and the pellet containing free RBC and rosettes were washed before counting. For the subsequent experiment the RBC were removed by NH&l (0.83%) treatment followed by washings in RPMI 1640 medium before cul- ture. The 30%-70% interface corresponding to nonroset- ting fraction was also recovered and washed before utiliza- tion.
2.3 LD analysis (LDA)
Rat thymus cells were prepared as previously described [ 121 and used as feeders at 3 x lo6 cells/ml. For each frequency determination, at least three different cell concentrations were set up with 32 or 48 replicates per point. Cultures were performed for 7 days in 96-well flat-bottom plates in RPMI supplemented with 10% FCS and 30 yglml LPS.
2.4 Determination of AG-specific and IgM-secreting clone frequencies
The frequency of LPS-responsive B cells was determined using an ELISA as described before [12]. Anti-single- stranded (ss)DNA antibodies (Ab) were detected in an ELISA using ssDNA-coated plates as described [13]. For determination of anti-BrMRBC reactivity we used a com- plement-dependent lysis test as previously reported [ 121. Mean absorbance+3SD obtained with SN of the 48 cultures which did not receive murine cells were used as the threshold of positivity. Results from LDA were statistically treated by the x2 minimization method as described by Taswell[14] and results are expressed as absolute frequen- cies, i. e. specific frequencies obtained by the Ag-specific ELISA or by hemolysis divided by total IgM frequencies obtained by the IgM-specific ELISA.
2.5 B cell colony assay
The different cell suspensions were cultured in double-layer colony technique, as described by Wu and Paige [15]. Briefly, the bottom layer corresponded to 1 ml Iscove's modified Dulbecco's medium, supplemented with 5 mg/ml delipidated BSA, 10% FCS, 0.3% melted Bacto Agar (Difco, Detroit, MI), 50 pg/ml LPS and 1% washed SRBC. Cells were resuspended in the same medium without LPS and SRBC, and plated as top layer. The cultures were incubated at 37"C, 5% C02, for 5-6 days and transferred by suction to Hybond N filters (Amersham Int., Amer- sham, GB).
The RNA blots were prehybridized overnight at 42°C in 50% formamide, 5 x SSPE, 4 x Denhardt's, 0.2% SDS and
100 mg/ml each, polyadenylic acid and denaturated E . coli DNA and hybridized with C,- and the 10 VH family-specific cDNA fragments for 48 h at 42 "C. After hybridization, the filters were washed once with 2xSSPE, 0.2% SDS for 30 min at 60°C and once with 1 x SSPE, 0.1% SDS for 30 min at 60°C, dried and autoradiographed for 3-14 days. The dots of colony-forming unit (CFU-B) were enumerated and the relative representation of each gene family plotted as the percentage of total VH gene expression.
2.6 Probes
The C, and VH gene family probes were gifts of Drs. F. Alt, S. Riley, 5. Kearney, U. Krawinkel and L. Reininger.The 10 VH probes used were vH81x (7183), V~300-19 (Q52),
(36-60), VHPNP B4 (5558) ,VHVgam. 3.8 (Vgam.3 A), VH31 (V31), vHcP12 (VHll) , and have been described else- where [4, 16-21]. The C, probe is the genomic fragment PGGN-1 described in Chih-Ping et al. [22]. Probes were labeled by the random priming method [23], using 32P- dCTP (Amersham).
V~S107 (s107),V~x24 (X24) , V H P B V ~ ~ (5606) ,V#6-3R1
3 Results
3.1 Abundance of BrMRBC-RFC in peritoneal and pleural cells
Normal splenic and peritoneal B lymphocytes are both able to form specific stable rosettes with BrMRBC (Fig. 1). Peritoneal cells, however, exhibited a relatively high fre- quency (9.5 iz 3.2%) of rosettes mainly composed of 10-18 RBC per B cell, while spleen cells showed a relatively low frequency (< YO) with a majority of rosettes containing fewer red blood cells: 5-7 per B cell (Fig. 1).
Frequency determinations of RFC in different cell popula- tions before and after enrichment on Percoll gradients are summarized in Table 1. In control experiments, the percen- tage of peritoneal and splenic cells forming rosettes with TNP-SRBC. SRBC alone or HRBC was found to be within
Figure I . Phase-contrast photograph of lymphoid cells from adult C57BL16 mice rosetted together with BrMRBC ( X 160): (a) typical peritoneal BrMRBC-RFC, (b) "spleen-type'' BrMRBC-RFC (< 10 erythrocytes per B cell) and (c) NR cells.

Eur. J. Immunol. 1990. 20: 1583-1589 VH1 I expression and anti-BrMRBC specificity 1585
Table 1. Percentages of RFC in different cell populations before and after enrichment on a 30%-70% Percoll gradient
Peritoneal cells Spleen cells Pleural Indicator before after before after cells RBC Percoll Percoll Percoll Percoll
BrMRBC 5.6 38a) 10") 60 6 60 9.5 66h) 8 65 5.5 6.3 76d)
13 63 11.6 57 15.7 54 10 32 6
11.7 13.9 10.3 67 8.3
9.5 k 3.2') 58 f 12.8c)
TNP-SRBC 3.3
SRBC 1 HRBC 0
2.7 '$la)
Total O d )
0.2 0 0.8 5
0.25 f 0 . 4 )
Larged) 5.4"' 7.7 1.6 1 I F ) 7.4 30h)
4.6 f 3.2c'
SmaW
Total 01,
2.3 1 0.5 1
the range previously described [24, 251. The relatively high frequency of BrMRBC-RFC in pleuropericardial and per- itoneal cavities (10%-12%) of adult mice, as compared to spleen (0.25%), correlates with the high anti-BrMRBC activity previously reported in these populations [26, 271. After separation of total spleen lymphocytes in large and small cell fractions on a Percoll gradient, all BrMRBC-RFC were recovered in the pool of large cells (4.6*3%) in agreement with previous observations that all such specif- icities are included in the activated lymphocyte pool [lo].
3.2 Enrichment in BrMRBC-RFC results in increased anti-BrMRBC B cell clonal precursor (BCP) frequency
The results of LDA carried out on BrMRBC rosetting (R) and nonrosetting (NR) fractions of splenic and peritoneal cells are summarized in Table2. They show that anti- BrMRBC BCP frequencies are 300 times higher in R than in NR fractions of peritoneal cells (1 : 14 vs. 1 : 4000). For splenic cells, no specific precursors were detected in NR fraction while they were present in the R fractions (1 : 296). Enrichment for anti-BrMRBC BCP in R fractions is specific, since anti-ssDNA BCP determined as a control in the same experiments are more frequent in the NR fraction.
3.3 BrMRBC-reactive B cell frequencies correlate positively with the frequency of B cells expressing V ~ l l genes
Since the percent of BrMRBC-RFC correlated with fre- quencies of anti-BrMRBC BCP, we have analyzed the VH
12")
a) Cell population submitted to VH typing in
b) Cell population submitted to LDA. c) MeankSD. d) Spleen cells are fractionated in large and small
RNA colony blot assay.
cells on a Percoll gradient.
gene family distribution of cell populations depleted or differentially enriched in RFC. Unfractionated populations of splenic, peritoneal and pleural cells showed a clear-cut difference in VHll expression which was higher in periton- eal and pleural cells (6%-12%) than in spleen cells (0.5%; Fig. 2). These results correlate with the percentages of BrMRBC-RFC observed in the above tissues, which were very low for the spleen cells (0.25%) but higher for peritoneal and pleural cells (9.5% and 12%, respectively; Table 1). There seems to be a gradation in the selection of V ~ l l expression in relation to the lymphoid compartment. As for specificity, the selection occurs in the following order: pleural cavity > peritoneal cavity >spleen.
Table 2. Absolute reciprocal frequencies of antigen-specific BCP in BrMRBC R and NR fractions of splenic and peritoneal cells of C57BL16 mice
Antigens Spleen Peritoneal cavity NR R NR R
(30%)a) (66%)
BrMRBC >lo0000 296 4Ooo 14
ssDNA 89 362 56 136 (190-660)b' (3000-6000) (10-24)
(69-125) (238-757) (42-83) (102-205)
a) % of RFC. b) 95% confidence limits.

1586 €? Poncet, F. Huetz, M.-A. Marcos and L. Andrade Eur. J. Immunol. 1990. 20: 1583-1589
When spleen cells were fractionated on the basis of their density on a Percoll gradient, all BrMRBC-RFC were recovered in the large cell pool (Table 1). The percent of v H 1 1 expression increased proportionally in this cell fraction as compared to the total spleen population (3% vs. 0.5%; Fig. 2). The major difference in VH distribution between pools of large and small cells was a tenfold lower value of VHll expression for small cells (0.1-0.3%) where no BrMRBC-RFC could be detected (Fig. 2).
These correlations are reinforced by the results obtained with peritoneal cells. Thus, when VH family distributions were studied in cell populations containing 38% or 76% of BrMRBC-RFC, 24% or 37%, respectively, of all VH genes
VH gene family
detected belonged to theVH11 family (Fig. 3). Fig. 4 shows typical autoradiographs obtained from R and NR fractions when filters were hybridized with CW-,V~J558- and vH11- specific probes. While V ~ l l genes were overrepresented in the R fraction, no V ~ l l gene expression was detected in NR fractions indicating an exclusive utilization of VHll genes for peritoneal anti-BrMRBC clones.
An increased representation of V ~ l l genes in the R fraction, concomitant to an enrichment of BrMRI3C-RFC, was also observed in splenic cells. Although the Percoll separation of splenic rosettes was more difficult due to their low frequency and characteristics, a 10-fold higher expres- sion of VHll genes was observed in the R fraction (5% in R
Figure 2. VH gene family distribution in clon- able B cells from splenic (Spl), peritoneal (PerC) and pleural (PL) cells of C57BL16 mice. (A) shows percentages of expression of V H l l gene family and (B) those of the nine other VH gene families. Each bar corresponds to the relative frequency of eachVH family in the total pool of VH genes expressed. A total of 3056 splenic, 1256 peritoneal and 260 pleural derived colonies expressing detecta- ble VH gene sequences were scored. The cloning efficiencies were roughly the same throughout the experiments and ranged from 1/4 to 116.
Figure 3. VH gene family distribu- tion in splenic (Spl) and peritoneal (Perc) BrMRBC-RFC R and NR clonable B cells. A total of 1209 splenic and 888 peritoneal colonies expressing detectable VH gene sequences were scored. The clon- ing efficiencies were roughly the same for all populations and ranged from 1/4 to 1/6.
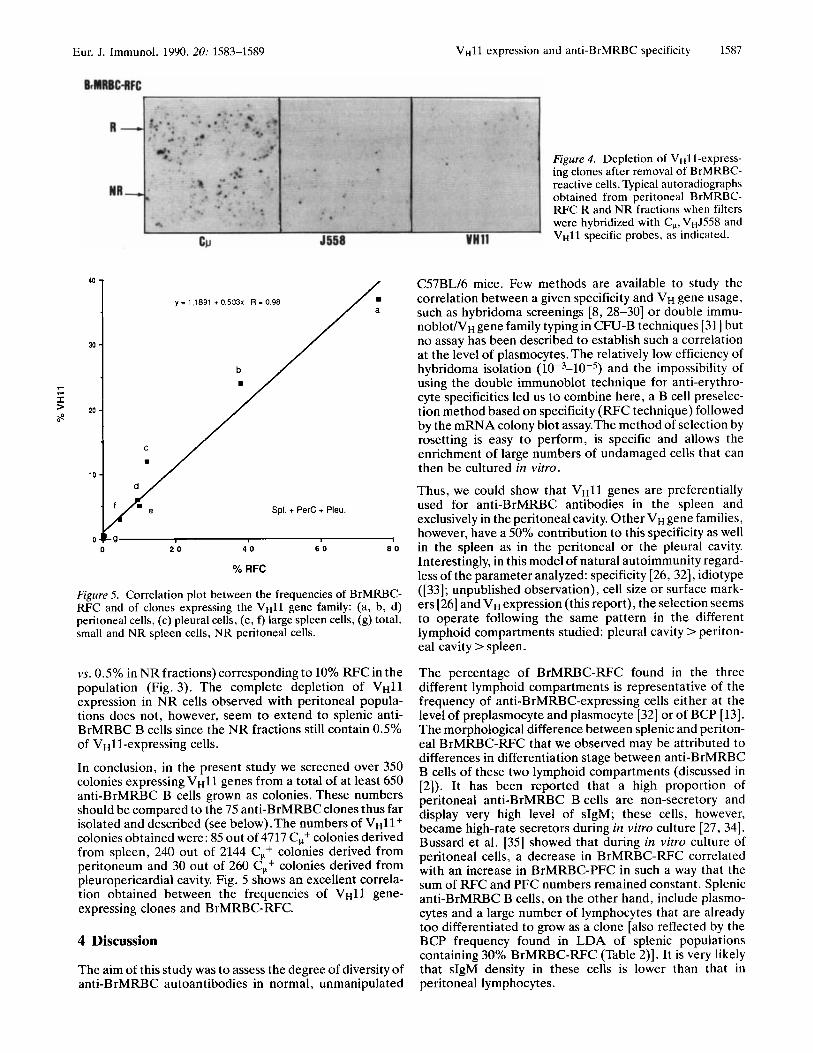
Eur. J. Immunol. 1990. 20: 1583-1589 VHll expression and anti-BrMRBC specificity 1587
r
f >
40 - y = 1.1891 + 0.503~ R = 0.98
30 -
20 -
Spl. + PerC + Pleu.
0 2 0 4 0 6 0 8 0
/
% RFC
Figure 5. Correlation plot between the frequencies of BrMRBC- RFC and of clones expressing the V H l l gene family: (a, b, d) peritoneal cells, (c) pleural cells, (e, f) large spleen cells, (8) total, small and NR spleen cells, NR peritoneal cells.
vs. 0.5% in NR fractions) corresponding to 10% RFC in the population (Fig. 3). The complete depletion of VHll expression in NR cells observed with peritoneal popula- tions does not, however, seem to extend to splenic anti- BrMRBC B cells since the NR fractions still contain 0.5% of V~11-expressing cells.
In conclusion, in the present study we screened over 350 colonies expressing VHll genes from a total of at least 650 anti-BrMRBC B cells grown as colonies. These numbers should be compared to the 75 anti-BrMRBC clones thus far isolated and described (see below).The numbers of vH11+ colonies obtained were: 85 out of 4717 C,+ colonies derived from spleen, 240 out of 2144 Cp+ colonies derived from peritoneum and 30 out of 260 Cp+ colonies derived from pleuropericardial cavity. Fig. 5 shows an excellent correla- tion obtained between the frequencies of VHll gene- expressing clones and BrMRBC-FWC.
4 Discussion
The aim of this study was to assess the degree of diversity of anti-BrMRBC autoantibodies in normal, unmanipulated
Figure 4 . Depletion of VHll-express- ing clones after removal of BrMRBC- reactive cells. Typical autoradiographs obtained from peritoneal BrMRBC- FWC R and NR fractions when filters were hybridized with C,, vHJ558 and V H l l specific probes, as indicated.
C57BL/6 mice. Few methods are available to study the correlation between a given specificity and VH gene usage, such as hybridoma screenings [8, 28-30] or double immu- nOblOtNH gene family typing in CFU-B techniques [31] but no assay has been described to establish such a correlation at the level of plasmocytes. The relatively low efficiency of hybridoma isolation ( 10-3-10-s) and the impossibility of using the double immunoblot technique for anti-erythro- cyte specificities led us to combine here, a B cell preselec- tion method based on specificity (RFC technique) followed by the mRNA colony blot assay.The method of selection by rosetting is easy to perform, is specific and allows the enrichment of large numbers of undamaged cells that can then be cultured in vitro.
Thus, we could show that VHll genes are preferentially used for anti-BrMRBC antibodies in the spleen and exclusively in the peritoneal cavity. Other VH gene families, however, have a 50% contribution to this specificity as well in the spleen as in the peritoneal or the pleural cavity. Interestingly, in this model of natural autoimmunity regard- less of the parameter analyzed: specificity [26, 321, idiotype ([33]; unpublished observation), cell size or surface mark- ers [26] and VH expression (this report), the selection seems to operate following the same pattern in the different lymphoid compartments studied: pleural cavity > periton- eal cavity > spleen.
The percentage of BrMRBC-RFC found in the three different lymphoid compartments is representative of the frequency of anti-BrMRBC-expressing cells either at the level of preplasmocyte and plasmocyte [32] or of BCP [ 131. The morphological difference between splenic and periton- eal BrMRBC-RFC that we observed may be attributed to differences in differentiation stage between anti-BrMRBC B cells of these two lymphoid compartments (discussed in [2]). It has been reported that a high proportion of peritoneal anti-BrMRBC B cells are non-secretory and display very high level of sIgM; these cells, however, became high-rate secretors during in vitro culture [27, 341. Bussard et al. [35] showed that during in vitro culture of peritoneal cells, a decrease in BrMRBC-RFC correlated with an increase in BrMRBC-PFC in such a way that the sum of RFC and PFC numbers remained constant. Splenic anti-BrMRBC B cells, on the other hand, include plasmo- cytes and a large number of lymphocytes that are already too differentiated to grow as a clone [also reflected by the BCP frequency found in LDA of splenic populations containing 30% BrMRBC-RFC (Table 2)]. It is very likely that sIgM density in these cells is lower than that in peritoneal lymphocytes.

1588 l? Poncet, F. Huetz, M.-A. Marcos and L. Andrade Eur. J. Immunol. 1990. 20: 1563-1589
The present report confirms with a large number of normal B cells selected on the basis of their ability to specifically form rosettes, that anti-BrMRBC antibodies preferentially utilize vH11 genes. TNP-SRBC-RFC enriched populations, used as control in the same experiments, do not show VHll gene family preference (data not shown). However, other VH genes must also encode anti-BrMRBC specificities. Thus, only about 50% of the RFC plated showed VH11 gene expression after growing as CFU-B (Fig. 5). This figure is somewhat lower than that obtained in the analysis of hybridomas and lymphomas. To date, out of 75 anti- BrMRBC hybridoma or lymphoma clones that have been described and typed for VH usage, 56 (75%) use VH1l genes, 12 (16%) use the recently described vH12 genes, 3 (4%) use V ~ J 5 5 8 genes and 3 were distributed in V~7183, VHs107 and V"Q52 VH families; 1 was not assigned in any known VH family [3, 4, 6-9, 361. The anti-BrMRBC specificity, therefore, seems to be preferentially but not exclusively associated with v H 1 1 expression.
Reciprocally, a small proportion (< YO) of V~11-express- ing clones was found in newborn spleen, BM and adult small spleen cells where no BrMRBC-reactive cells could be detected ([lo, 13, 371, Fig. 2). Furthermore, we and others have previously reported the existence of splenic VH11-expressing clones deprived of anti-BrMRBC specif- icity [7, 8, 381.
The incomplete inclusion of VH11-expressing clones in the pool of anti-BrMRBC-producing cells seems, however, specific to the spleen. Thus, our results demonstrated that no V ~ l l gene expression was detected in BrMRBC- RFC-depleted peritoneal cells, i.e. all VHl1 gene segments expressed in this population encode the anti-BrMRBC specificity. This strong association between utilization of a VH gene (vH11) and autoreactivity (anti-BrMRBC) is in contrast with what has been described for other autoim- mune models [30,39-411 and for preimmune repertoires [31,42-441 but is similar to what is observed in some responses to specific immunization [42, 43, 45, 461. This is likely the result of a high selective pressure acting on V regions (a combination of VH and V,) either via antigen andor idiotypic interactions.
The corresponding antigenic ligands, although not yet defined, may be akin to polar heads of phosphatidyl choline (33,47-491, an ubiquitous molecule in biological mem- brane. The involvement of external antigens, such as components of commensal bacteria, is unlikely since germ- free and specific pathogen-free mice show comparable frequencies of anti-BrMRBC activity and VH11 gene family expression ([50], Huetz et al., to be published). Concerning autoantigens, no evidence for an equivalent bromelain treatment of the RBC exist in vivo [51] and the soluble analogue of the epitope associated with low-density lipo- protein [52] is present in the mouse serum throughout development of the immune system; this does not fit with a delayed expression of anti-BrMRBC response assumed to take place [13, 53-55] if an autoantigen-driven selection operates.
The existence of natural autoanti-idiotypic antibodies has also been reported but their involvement in the regulation of anti-BrMRBC-producing cells has not been shown [56]. Preliminary data (not shown) suggest an autobinding
capacity of anti-BrMRBC antibodies (autobodies). This raises the possibility that in vivo selection and expansion of BrMRBC-reactive B cells might be mediated by a process based on autostimulation, the secreted pentameric form of the molecule binding to the monomeric membrane form or through a specific cell-to-cell interaction via sIgM.
It is currently postulated that most anti-BrMRBC precom- mitted B cells belong to the Ly-1 B cell subset [2]. In line with this assertion is the biased usage of the VHl1 gene family observed by sorted peritoneal Ly-1 B cells from adult mice [5, 6, 571. Likewise, resident pleuropericardial lym- phocytes, a phenotypically homogeneous Ly-1 B cell population [26], showed high frequencies of anti-BrMRBC clones and VHll gene family expression ([26], Fig. 2). However, anti-BrMRBC B cells cannot be taken as quan- titative markers of Ly-1 B cells since, for instance, in newborn spleen or in the motheaten mouse, the increased frequency of Ly-1 B cells [58,59] is accompanied by a lack of reactivity towards BrMRBC [13, 531.
We thank Pr. Antonio Coutinho for helpful comments and sugges- tions throughout this work.
Received January 13, 1990; in revised form April 17, 1990.
5 References
1 Coutinho, A. , Immunol. Rev. 1989. 110: 63. 2 Huetz, F. and Poncet, P., Res. Irnmunol. 1989. 140: 75. 3 Reininger, L., Ollier, l?, Poncet, P., Kaushik, A . and Jaton, J.
C., J. Immunol. 1987. 138: 316. 4 Reininger, L. , Kaushik, A., Izui, S. and Jaton, J. C., Eur. J.
Immunol. 1988. 209: 1348. 5 Pennell, C. A . , Mercolino,T. J. , Grdina,T. A. , Arnold, L. W.,
Haughton, G. and Clarke, S. H., Eur. J. Zmmunol. 1989. 19: 1289.
6 Hardy, R. R., Carmack, C. E., Shinton, S. A . , Riblet, R. J. and Hayakawa, K. , J. Immunol. 1989. 142: 3643.
7 Pennell, C. A . , Arnold, L.W., Haughton, G. and Clarke, S. H . , J. Immunol. 1988. 141: 2788.
8 Poncet, P , Reininger, L., Freitas, A . , Holmberg, D., Dighiero, G. and Coutinho, A . , Res. Immunol. 1989. 140: 255.
9 Conger, J. D., Sage, H. J. and Corley, R. B., J. lmrnunol. 1990. 143: 4044.
10 Portnoi, D., Freitas, A . , Holmberg, D., Bandeira, A. and Coutinho, A . , J. Exp. Med. 1986. 164: 25.
11 Weyer, J. and Bussard, A. E., Ann. Irnmunol. (Inst. Pasteur) 1974. 125C: 947.
12 Huetz, F., Larsson-Sciard, E. L., Pereira, l?, Portnoi, D. and Coutinho, A. , Eur. J. Immunol. 1988. 18: 1615.
13 Huetz, F., Poncet, l?, Coutinho, A. and Portnoi, D., Eur. J. lmmunol. 1989. 19: 1195.
14 Taswell, C., J. Irnmunol. 1981. 126: 1614. 15 Wu, G. E. and Paige, C. J., EMBO J. 1986. 5: 3476. 16 Winter, E . . Radbruch, A. and Krawinkel, U., EMBO J. 1985.
4: 2861. 17 Yancopoulos, G. D., Desiderio, S.V., Paskind, M., Kearney, J.
F., Baltimore, D. and Alt, F. W., Nature 1984. 311: 727. 18 Reth, M. G. , Jackson, E. C. and Alt, F.W., EMBO J. 1986. 5:
2131. 19 Near, R. G. , Juszcyak, E. C., Huang, S. Y , Sicar, S. A. ,
Margolies, M. N. and Gefter, M. L., Proc. Natl. Acad. Sci. USA 1984. 81: 2167.
20 Perlmutter, R . M., Klotz, J. L., Bond, M. W., Nahm, M., Davie, J. M. and Hood, L., J. Exp. Med. 1984. 159: 179.
21 Hartman, A . B. and Rudikoff, S., EMBO J. 1984. 3: 3023.

Eur. J. Immunol. 1990.20: 1583-1589 VH1 1 expression and anti-BrMRBC specificity 1589
41 Kofler, R., Noonan, D. J., Strohal, R., Balderas, R. S., Moller, N. P. H., Dixon, J. F. and Theofilopoulos, A. N., Eur. J. Immunol. 1987. 17: 91.
42 Berek, C. and Milstein, C., Immunol. Rev. 1988. 105: 5 . 43 Victor-Kobrin, C., Bonilla, F. A. and Bona, C., Immunol. Rev.
44 Manser, T., Huang, S. Y. and Gefter, M., Science 1984. 226:
45 Crews, S., Griffin, J., Huang, H., Calame, K. and Hood, L..
46 Kimura, K., Cook, R., Meek, K., Umeda, M., Ball, E. , Capra,
47 Kawaguchi, S., Immunology 1987. 62: 11. 48 Mercolino,T. J., Arnold, L.W. and Haugthon, G., J. Exp. Med.
49 Cox, K. 0. and Hardy, S., J. Immunol. 1985. 55: 263. 50 Cunningham, A. J., Transplant. Rev. 1976. 31: 23. 51 Cunliffe, D. A. and Cox, K. O., Mol. Immunol. 1979. 16:
427. 52 Linder, E. J. and Edgington, T. S., J. Immunol. 1973. I I O :
53. 53 Mercolino,T. J., Arnold, L.W., Hawskins, L. A. and Haugthon,
G., J. Exp. Med. 1988. 168: 687. 54 Fujiwara, M. and Akiyama, Y., Cell. Immunol. 1980. 55:
366. 55 Cunningham, A. J. and Steele, E., J. Clin. Exp. Immunol.
1981. 44: 38. 56 Wieties, K. M. and Moticka, E. J., Cell. Immunol. 1986. 98:
68. 57 Andrade, L., Freitas, A. A., Huetz, F., Poncet, P. and
Coutinho, A., Eur. J. Immunol. 1989. 19: 1117. 58 Herzenberg, L. A., Stall, A., Lalor, I?, Sidman, C., Moore,W.,
Park, D. and Herzenberg, L. A., Immunol. Rev. 1986. 93: 81.
59 Sidman, C. L., Shultz, L. D., Hardy, R. R., Hayakawa, K. and Herzenberg, L. A., Science 1986. 232: 1423.
1989. 110: 151.
1283.
Cell 1981. 25: 59.
J. D. and Marcus, D. M., J. Immunol. 1988. 140: 1212.
1986. 163: 155.
22 Chih-Ping, L.,Tucker, P.W., Mushinski, J. and Blattner, F. R., Science 1986. 209: 1348.
23 Feinberg, A. F! and Vogelstein, B., Anal. Biochem. 1983.132: 6.
24 Greaves, M. F., MBller, E. and Moller, G., Cell. Immunol. 1970. I : 386.
25 Moller, E., Bullock, W. W. and Makela, O., Eur. J. Immunol. 1973. 3: 172.
26 Marcos, M. A. R., Huetz, F., Pereira, F!, Andreu, J. L., Martinez-A, C. and Coutinho, A., Eur. J. Immunol. 1989.19: 2031.
27 Pages, J. M. and Bussard, A. E., Nature 1975. 257: 316. 28 Holmberg, D., Eur. J. Immunol. 1987. 17: 399. 29 Glotz, D., Sollazzo, M., Riley, S. and Zanetti, M., J. Immunol.
1988. 141: 383. 30 Duchosal, M. A., Kofler, R., Balderas, R. S., Aguado, M.T.,
Dixon, F. J. and Theofilopoulos, A. N., J. Immunol. 1989.142: 1737.
31 Kelsoe, G., Miceli, R., Cerny, J. and Schulze, D. H., Immu- nogenetics 1989. 29: 288.
32 Steele, E. J. and Cunningham, A. J., Nature 1978. 274: 483. 33 Poncet, P., Kocher, H. F!, Pages, J., Jaton, J. C. and Bussard, A.
E., Mol. Immunol. 1985. 22: 541. 34 Daenke, S. and Cox, K. O., Scand. J. Immunol. 1986. 23:
567. 35 Bussard, A, , Rattner, E., Vanko, L. and Pages, J., Ann.
Immunol. (Inst. Pasteur) 1977. 128C: 319. 36 Mercolino, T. J., Locke, A. L., Afshari, A. L., Sasser, D.,
Travis, W. W., Arnold, L. W. and Haugthon, G., J. Exp. Med. 1989. 169: 1869.
37 Garzelli, C., Forlani, A.,Lombardi, S., Colizzi,V., Carnpa, M. and Falcone, G., Int. J. Allergy Appl. Immunol. 1983. 72: 253.
38 Braun, J. and King, L., J. Mol. Cell. Immunol. 1989. 4: 121. 39 Painter, C. J., Monestier, M., Chew, A., Bona-Dimitriu, A.,
Kasturi, K., Bailey, C., Scott,V. E., Sidman, C. L. and Bona, C. A., J. Exp. Med. 1988. 167: 1137.
40 Shlomchik, M. J., Nemazee, D. A., Sato,V. L. ,Van Snick, J., Carsson, A. and Weigert, M. G., J. Exp. Med. 1986. 164: 407.




















