
A NEW SPECIES OF SPHAEROMYXA (MYXOSPOREA: SPHAEROMYXINA: SPHAEROMYXIDAE) IN DEVIL FIREFISH, PTEROIS...
9
1434 J. Parasitol., 90(6), 2004, pp. 1434–1442 q American Society of Parasitologists 2004 A NEW SPECIES OF SPHAEROMYXA (MYXOSPOREA: SPHAEROMYXINA: SPHAEROMYXIDAE) IN DEVIL FIREFISH, PTEROIS MILES (SCORPAENIDAE), FROM THE NORTHERN RED SEA: MORPHOLOGY, ULTRASTRUCTURE, AND PHYLOGENY A. Diamant, C. M. Whipps*, and M. L. Kent* Israel Oceanographic and Limnological Research Ltd., National Center for Mariculture, Eilat 88112, Israel. e-mail: [email protected] ABSTRACT: Sphaeromyxa zaharoni n. sp. (Myxosporea) is described from the gallbladder of devil firefish, Pterois miles (Scor- paenidae), from coral reefs of the Gulf of Eilat, Israel, northern Red Sea. The parasite was found also in bearded scorpionfish, Scorpaenopsis barbata, from the same area. This is the first report on Sphaeromyxa sp. from this zoogeographical region. The plasmodia are amoeboid when young, becoming disc-shaped and elongated when mature. In paraffin sections, the plasmodium periphery appears as a finely granulated, strongly eosinophilic layer with an intricate surface membrane network. Sphaeromyxa zaharoni n. sp. is polysporous, disporoblastic, and has asynchronous sporogenesis. The mature spore is elongated and fusiform, has delicately ridged valves, and contains a single, binucleated sporoplasm. In valvular view, the tips are truncated. The mean spore size is length 14.5 mm, width 4.8 mm, and polar capsule 4.8 3 3.4 mm. The 2 equally sized ovoid polar capsules are positioned at opposite ends of the spore, each containing a filament loosely folded in 2 loops. The fine structure of the sporoblast and spore corresponded with previously studied Sphaeromyxa species. According to small-subunit ribosomal DNA gene sequence analysis, S. zaharoni n. sp. is most closely related to 2 Myxidium spp. The close phylogenetic relatedness of Sphaeromyxa and Myxidium and similar spore morphology raises the question whether these 2 genera should be maintained in separate families and suborders. Thirty-eight species of the myxosporean Sphaeromyxa, all coelozoic from marine fish hosts, have so far been described and named (Lom, 2004). Sphaeromyxa spp., like other gall- bladder-inhabiting myxozoan species (Myxidium, Ceratomyxa, Chloromyxum, and Leptotheca), have long been regarded as having little or no significance as fish pathogens (Fantham and Porter, 1912; Lom and Dykova ´, 1992). The available informa- tion on Sphaeromyxa spp. is incomplete, dealing mainly with morphology and ultrastructure (Lom and de Puytorac, 1965; Lom, 1969, 2004; Grasse ´ and Lavette, 1978; Gracia et al., 1997). To date, no molecular study has been carried out on species in this genus. In this article, we describe a new Sphae- romyxa species, the first from the Red Sea region, and discuss the phylogeny of both family and genus within the Myxozoa, as reflected by its small-subunit ribosomal DNA (SSU rDNA) sequence. MATERIALS AND METHODS Sample collection Twenty-four devil firefish, Pterois miles (Bennett, 1828), and 4 beard- ed scorpionfish, Scorpaenopsis barbata (Ru ¨ ppell, 1838), were examined between September 2002 and March 2003 from 3 coastal locations of the Gulf of Eilat, Israel (Red Sea). The fish were measured, weighed, and dissected within several hours of capture. The external surfaces and internal organs of the fish were thoroughly examined for the presence of parasites and lesions. Myxosporean plasmodia and spores found in the gallbladder were examined in fresh bile mounts, measured, photographed, and drawn from observations using phase contrast and Nomarski interference mi- croscopy. For additional observations, air-dried bile smears were stained with the Giemsa method. Parasites were also fixed in 10% buffered neutral formalin and processed for histological examination. For trans- mission electron microscopy (TEM), plasmodia were fixed in 2.5% glu- taraldehyde in cacodylate buffer and postfixed with osmium tetroxide, stained en bloc with uranyl acetate, and embedded in resin blocks. Lead citrate–stained thin sections were mounted on copper grids and exam- ined with a JEOL 100CXII transmission electron microscope. Received 13 February 2004; revised 25 March 2004; accepted 8 April 2004. * Center for Fish Disease Research, Department of Microbiology, 220 Nash Hall, Oregon State University, Corvallis, Oregon 97331-3404. DNA analysis Plasmodia for molecular analysis were fixed in 90% ethanol. Nucleic acid was extracted using the DNeasy Tissue Kit (Qiagen Inc., Valencia, California). Overlapping regions of SSU rDNA were polymerase chain reaction (PCR) amplified using primers 18E (59-CTG GTT GAT CCT GCC AGT) with Myxgen2r (59-CAR ATG CYT TCG CWY TTG TTA) and Myxgen4F (59-GTG CCT TGA ATA AAT CAG AG) with 18R (59-CTA CGG AAA CCT TGT TAC G). PCRs were carried out with the Taq PCR core kit (Qiagen Inc.), following the manufacturer’s pro- tocols. Amplified fragments were observed on a 1% agarose gel con- taining 100 ng/ml ethidium bromide, excised, and purified. Primers as used for PCR were used to sequence purified amplicons in both direc- tions using AP Biotecht DYEnamic ET Terminator cycle sequencing chemistry with Thermo Sequenase II (Amersham Biosciences, Pisca- taway, New Jersey) on an ABI PRISMt 377 DNA Sequencer (Applied Biosystems, Foster City, California). Resulting sequences were com- bined, and the 1,876–base pair sequence for Sphaeromyxa zaharoni n. sp. was deposited in GenBank (AY538662). Phylogenetic analyses Variisporinid SSU rDNA sequences of adequate length from pub- lished reports and representatives from the other major myxozoan groupings (Kent et al., 2001) were aligned with ClustalX (Thompson et al., 1997) and are as follows: Ceratomyxa labracis (AF411472), Cer- atomyxa sparusaurati (AF411471), Ceratomyxa shasta (AF001579), Zschokkella mugilis (AF411336), Enteromyxum scopthalmi (AF411335), Enteromyxum leei (AF411334), Myxidium truttae (AF201374), Myxidium lieberkuehni 1 mx18 (X76638), M. lieberkuehni 2 mx 19 (X76639), Myxidium sp. (U13829), Parvicapsula minibicornis (AF201375), Sphaerospora molnari (AF378345), Sphaerospora oncho- rynchi (AF201373), Kudoa thyrsites (AY152747), Kudoa crumena (AF378347), Myxobolus pendula (AF378340), Myxobolus cerebralis (U96492), Henneguya exilis (AF021881), and Tetracapsuloides bryos- almonae (U70623). All phylogenetic analyses were conducted in PAUP*4.0b1 (Swofford, 1998). Parsimony analyses used a heuristic search algorithm with 50 random additions of sequences and tree bi- section–reconnection (TBR) branch swapping. Bootstrap confidence values were calculated with 1,000 replicates using the same heuristic search as above. Neighbor-joining analyses were conducted using ge- netic distances calculated with maximum likelihood models described below. Bootstrap confidence values were calculated with 100 replicates. An optimal evolutionary model for the SSU data set was estimated using Modeltest 3.06 (Posada and Crandall, 1998). The GTR1I1G model was used. Maximum likelihood analysis used a heuristic search algorithm with 10 random sequence additions and TBR branch swap- ping. Bootstrap values (n 5 100) were calculated for maximum likeli-
-
Upload
chuaungthu -
Category
Documents
-
view
5 -
download
0
Transcript of A NEW SPECIES OF SPHAEROMYXA (MYXOSPOREA: SPHAEROMYXINA: SPHAEROMYXIDAE) IN DEVIL FIREFISH, PTEROIS...

1434
J. Parasitol., 90(6), 2004, pp. 1434–1442q American Society of Parasitologists 2004
A NEW SPECIES OF SPHAEROMYXA (MYXOSPOREA: SPHAEROMYXINA:SPHAEROMYXIDAE) IN DEVIL FIREFISH, PTEROIS MILES (SCORPAENIDAE), FROM THENORTHERN RED SEA: MORPHOLOGY, ULTRASTRUCTURE, AND PHYLOGENY
A. Diamant, C. M. Whipps*, and M. L. Kent*Israel Oceanographic and Limnological Research Ltd., National Center for Mariculture, Eilat 88112, Israel. e-mail: [email protected]
ABSTRACT: Sphaeromyxa zaharoni n. sp. (Myxosporea) is described from the gallbladder of devil firefish, Pterois miles (Scor-paenidae), from coral reefs of the Gulf of Eilat, Israel, northern Red Sea. The parasite was found also in bearded scorpionfish,Scorpaenopsis barbata, from the same area. This is the first report on Sphaeromyxa sp. from this zoogeographical region. Theplasmodia are amoeboid when young, becoming disc-shaped and elongated when mature. In paraffin sections, the plasmodiumperiphery appears as a finely granulated, strongly eosinophilic layer with an intricate surface membrane network. Sphaeromyxazaharoni n. sp. is polysporous, disporoblastic, and has asynchronous sporogenesis. The mature spore is elongated and fusiform,has delicately ridged valves, and contains a single, binucleated sporoplasm. In valvular view, the tips are truncated. The meanspore size is length 14.5 mm, width 4.8 mm, and polar capsule 4.8 3 3.4 mm. The 2 equally sized ovoid polar capsules arepositioned at opposite ends of the spore, each containing a filament loosely folded in 2 loops. The fine structure of the sporoblastand spore corresponded with previously studied Sphaeromyxa species. According to small-subunit ribosomal DNA gene sequenceanalysis, S. zaharoni n. sp. is most closely related to 2 Myxidium spp. The close phylogenetic relatedness of Sphaeromyxa andMyxidium and similar spore morphology raises the question whether these 2 genera should be maintained in separate familiesand suborders.
Thirty-eight species of the myxosporean Sphaeromyxa, allcoelozoic from marine fish hosts, have so far been describedand named (Lom, 2004). Sphaeromyxa spp., like other gall-bladder-inhabiting myxozoan species (Myxidium, Ceratomyxa,Chloromyxum, and Leptotheca), have long been regarded ashaving little or no significance as fish pathogens (Fantham andPorter, 1912; Lom and Dykova, 1992). The available informa-tion on Sphaeromyxa spp. is incomplete, dealing mainly withmorphology and ultrastructure (Lom and de Puytorac, 1965;Lom, 1969, 2004; Grasse and Lavette, 1978; Gracia et al.,1997). To date, no molecular study has been carried out onspecies in this genus. In this article, we describe a new Sphae-romyxa species, the first from the Red Sea region, and discussthe phylogeny of both family and genus within the Myxozoa,as reflected by its small-subunit ribosomal DNA (SSU rDNA)sequence.
MATERIALS AND METHODS
Sample collection
Twenty-four devil firefish, Pterois miles (Bennett, 1828), and 4 beard-ed scorpionfish, Scorpaenopsis barbata (Ruppell, 1838), were examinedbetween September 2002 and March 2003 from 3 coastal locations ofthe Gulf of Eilat, Israel (Red Sea). The fish were measured, weighed,and dissected within several hours of capture. The external surfaces andinternal organs of the fish were thoroughly examined for the presenceof parasites and lesions.
Myxosporean plasmodia and spores found in the gallbladder wereexamined in fresh bile mounts, measured, photographed, and drawnfrom observations using phase contrast and Nomarski interference mi-croscopy. For additional observations, air-dried bile smears were stainedwith the Giemsa method. Parasites were also fixed in 10% bufferedneutral formalin and processed for histological examination. For trans-mission electron microscopy (TEM), plasmodia were fixed in 2.5% glu-taraldehyde in cacodylate buffer and postfixed with osmium tetroxide,stained en bloc with uranyl acetate, and embedded in resin blocks. Leadcitrate–stained thin sections were mounted on copper grids and exam-ined with a JEOL 100CXII transmission electron microscope.
Received 13 February 2004; revised 25 March 2004; accepted 8 April2004.
* Center for Fish Disease Research, Department of Microbiology, 220Nash Hall, Oregon State University, Corvallis, Oregon 97331-3404.
DNA analysis
Plasmodia for molecular analysis were fixed in 90% ethanol. Nucleicacid was extracted using the DNeasy Tissue Kit (Qiagen Inc., Valencia,California). Overlapping regions of SSU rDNA were polymerase chainreaction (PCR) amplified using primers 18E (59-CTG GTT GAT CCTGCC AGT) with Myxgen2r (59-CAR ATG CYT TCG CWY TTG TTA)and Myxgen4F (59-GTG CCT TGA ATA AAT CAG AG) with 18R(59-CTA CGG AAA CCT TGT TAC G). PCRs were carried out withthe Taq PCR core kit (Qiagen Inc.), following the manufacturer’s pro-tocols. Amplified fragments were observed on a 1% agarose gel con-taining 100 ng/ml ethidium bromide, excised, and purified. Primers asused for PCR were used to sequence purified amplicons in both direc-tions using AP Biotecht DYEnamic ET Terminator cycle sequencingchemistry with Thermo Sequenase II (Amersham Biosciences, Pisca-taway, New Jersey) on an ABI PRISMt 377 DNA Sequencer (AppliedBiosystems, Foster City, California). Resulting sequences were com-bined, and the 1,876–base pair sequence for Sphaeromyxa zaharoni n.sp. was deposited in GenBank (AY538662).
Phylogenetic analyses
Variisporinid SSU rDNA sequences of adequate length from pub-lished reports and representatives from the other major myxozoangroupings (Kent et al., 2001) were aligned with ClustalX (Thompsonet al., 1997) and are as follows: Ceratomyxa labracis (AF411472), Cer-atomyxa sparusaurati (AF411471), Ceratomyxa shasta (AF001579),Zschokkella mugilis (AF411336), Enteromyxum scopthalmi(AF411335), Enteromyxum leei (AF411334), Myxidium truttae(AF201374), Myxidium lieberkuehni 1 mx18 (X76638), M. lieberkuehni2 mx 19 (X76639), Myxidium sp. (U13829), Parvicapsula minibicornis(AF201375), Sphaerospora molnari (AF378345), Sphaerospora oncho-rynchi (AF201373), Kudoa thyrsites (AY152747), Kudoa crumena(AF378347), Myxobolus pendula (AF378340), Myxobolus cerebralis(U96492), Henneguya exilis (AF021881), and Tetracapsuloides bryos-almonae (U70623). All phylogenetic analyses were conducted inPAUP*4.0b1 (Swofford, 1998). Parsimony analyses used a heuristicsearch algorithm with 50 random additions of sequences and tree bi-section–reconnection (TBR) branch swapping. Bootstrap confidencevalues were calculated with 1,000 replicates using the same heuristicsearch as above. Neighbor-joining analyses were conducted using ge-netic distances calculated with maximum likelihood models describedbelow. Bootstrap confidence values were calculated with 100 replicates.An optimal evolutionary model for the SSU data set was estimatedusing Modeltest 3.06 (Posada and Crandall, 1998). The GTR1I1Gmodel was used. Maximum likelihood analysis used a heuristic searchalgorithm with 10 random sequence additions and TBR branch swap-ping. Bootstrap values (n 5 100) were calculated for maximum likeli-

DIAMANT ET AL.—S. ZAHARONI N. SP. FROM RED SEA DEVIL FIREFISH 1435
FIGURES 1–6. 1. Line drawing of Sphaeromyxa zaharoni n. sp. Bar 5 5 mm. 2. Spores, some paired, in fresh bile mount (Nomarski interference).Bar 5 10 mm. 3, 4. Air-dried, Giemsa-stained spores. Bar 5 10 mm. 5. Spores showing contours of the polar filament (Nomarski interference).Bar 5 10 mm. 6. Aberrant dumbbell-shaped spore of S. zaharoni n. sp. (Nomarski interference). Bar 5 10 mm.
hood trees using default parameter in PAUP* because more complexmodels made these calculations computationally intractable.
DESCRIPTION
Eleven of 24 (45.8%) devil firefish measuring 210–355 mm(total length) and 2 of 4 (50%) bearded scorpionfish measuring185–255 mm harbored myxosporean plasmodia in the gallblad-der. On the basis of spore morphology and plasmodium shape,size, and site of infection, this myxosporean was placed in thegenus Sphaeromyxa Thelohan, 1892.
Sphaeromyxa zaharoni n. sp.(Figs. 1–11)
Spores: Mature spore fusiform, straight, tips truncated infrontal (valvular) view and slightly rounded in sutural view(Figs. 1–6). Valves equal in size, with longitudinal striations;difficult to discern in fresh material, but evident in Giemsa-stained spores (Fig. 4), and most clearly seen in TEM sections(see Fig. 21). Polar capsules ovoid, similarly sized, and posi-
tioned at opposite ends of spore. Polar capsule 1.3–1.4 timesas long as wide and contains a polar filament folded in 2 looseloops. Extruded filaments short and stubby. Sporoplasm binu-cleated and centrally located in spore cavity between the 2 polarcapsules (see Fig. 3). Aberrant dumbbell-shaped spores some-times occurred in bile (Fig. 6).
Spore measurements (in microns): Pterois miles (n 5 25),length 14.5 3 4.8 (range 13.7–15.1 3 4.2–5.5), polar capsule4.8 3 3.4 (range 3.3–5.6 3 2.5–4.0); S. barbata (n 5 25),length 14.2 3 4.6 (range 13.25–15.75 3 3.25–5.7), polar cap-sule 4.7 3 3.3 (3.5–5.3 3 2.8–4.4).
Plasmodia: Parasites float freely in bile, large plasmodia eas-ily seen through gallbladder wall (Fig. 7). Each large plasmo-dium partly wrapped around itself as well as around neighbor-ing plasmodia, forming loose parasite mass oftentimes occu-pying significant part of gallbladder volume. When removedand flattened in physiological saline, plasmodia slowly crinkleand fold. Small plasmodia (,50 mm) often amoeboid with lo-bose pseudopodia (Figs. 8, 9); display some mobility and ap-

1436 THE JOURNAL OF PARASITOLOGY, VOL. 90, NO. 6, DECEMBER 2004
FIGURES 7–11. Plasmodia of Sphaeromyxa zaharoni n. sp. 7. Pterois miles gallbladder packed with mature S. zaharoni n. sp. plasmodia. Bar5 250 mm. 8. Young, amoeboid S. zaharoni n. sp. plasmodium with lobose pseudopodia in a wet bile mount. Note spherical fragments presumablygenerated by plasmotomy (arrows). Bar 5 25 mm. 9. Low-power magnification of young, amoeboid and disc-shaped S. zaharoni n. sp. plasmodia.Bar 5 25 mm. 10. Low-power magnification of pancake-shaped S. zaharoni n. sp. plasmodia. Note clearly demarcated peripheral ectoplasm. Bar5 50 mm. 11. High-power magnification of peripheral region of S. zaharoni n. sp. plasmodium. Bar 5 10 mm.
pear to shed small, spherical fragments by plasmotomy. Disc-shaped plasmodia vary greatly in size, several micrometers to3 cm, although in 1 devil firefish with 25 plasmodia, all ratheruniform in size (in mm), 200–270 3 210–550 (mean 230 3410, n 5 25). Parasites disc-shaped or elongated, bluish hyaline
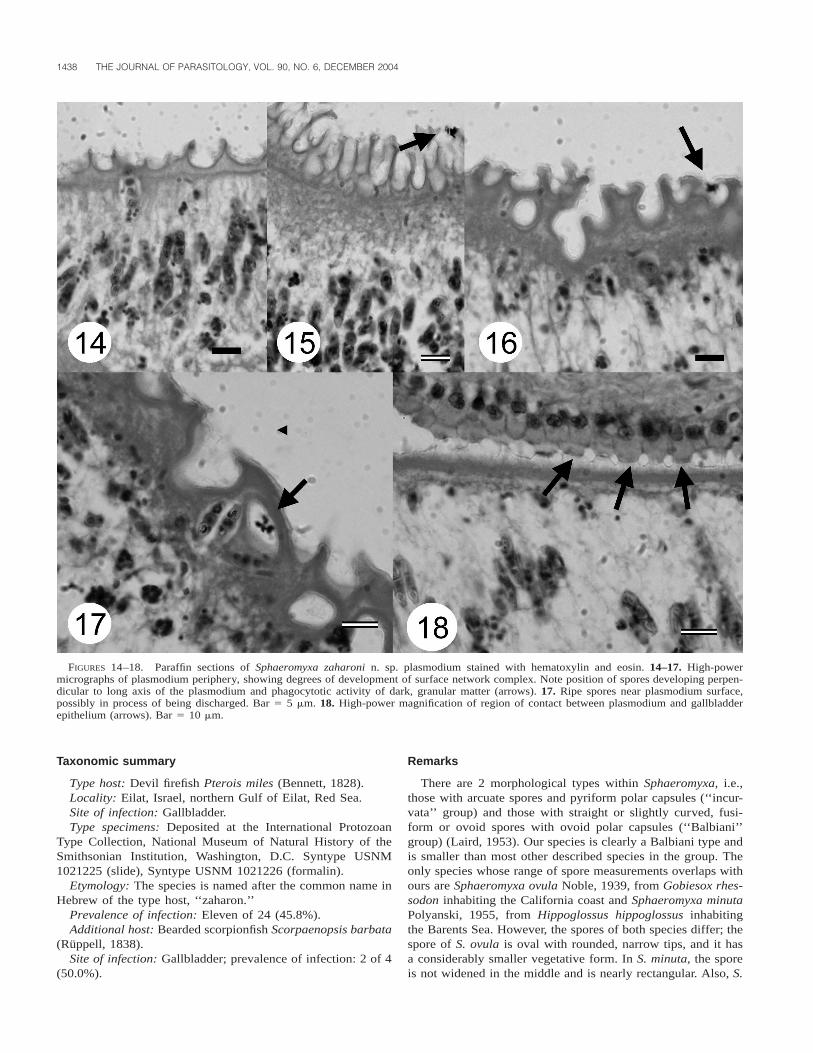
hue, clearly demarcated ectoplasm (Figs. 10, 11). Plasmodia.1,200 mm usually elongated, with brownish color. Refractivespherules and dark bodies visible in cytoplasm. Ectoplasm,;15–20 mm in width, strongly eosinophilic (Figs. 12–18). Spo-rogenesis asynchronous, earliest developmental stages located

DIAMANT ET AL.—S. ZAHARONI N. SP. FROM RED SEA DEVIL FIREFISH 1437
FIGURES 12–13. 12. Paraffin section through a parasitized Pterois miles gallbladder, showing packed plasmodia. No host tissue damage isevident despite the heavy infection. Bar 5 50 mm. 13. Periphery of plasmodium, showing the various layers. M, membrane surface network;ECT, eosinophilic ectoplasm; GC, generative cells; AE, alveolar endoplasm; SB, sporoblasts. Bar 5 15 mm.
near plasmodium periphery (Fig. 13). Outer surface with ele-vated crests and invaginations, producing irregular, intricatenetwork covered by single unit membrane with numerous mi-croscopic cytoplasmic extensions (Figs. 13–17). No damage togallbladder lining observed, although with evidence of plas-modium attachment to (but no invasion of) host epithelium(Fig. 18).
Plasmodium polysporous and disporoblastic; pairs of sporesin large numbers within endoplasm, positioned perpendicular toplasmodium long axis. In some cases, spores near plasmodiumouter membrane, possibly in process of being discharged fromplasmodium (Fig. 17).
Ultrastructure: Fine granular, fuzzy surface coat of amor-phous substance, approximately 5 mm thick, covered outer plas-modium membrane. Plasmodium surface undulated and delim-ited by continuous single cell membrane, with protrusions pos-sessing irregular microvilli. Beneath, 2 distinct ectoplasm and
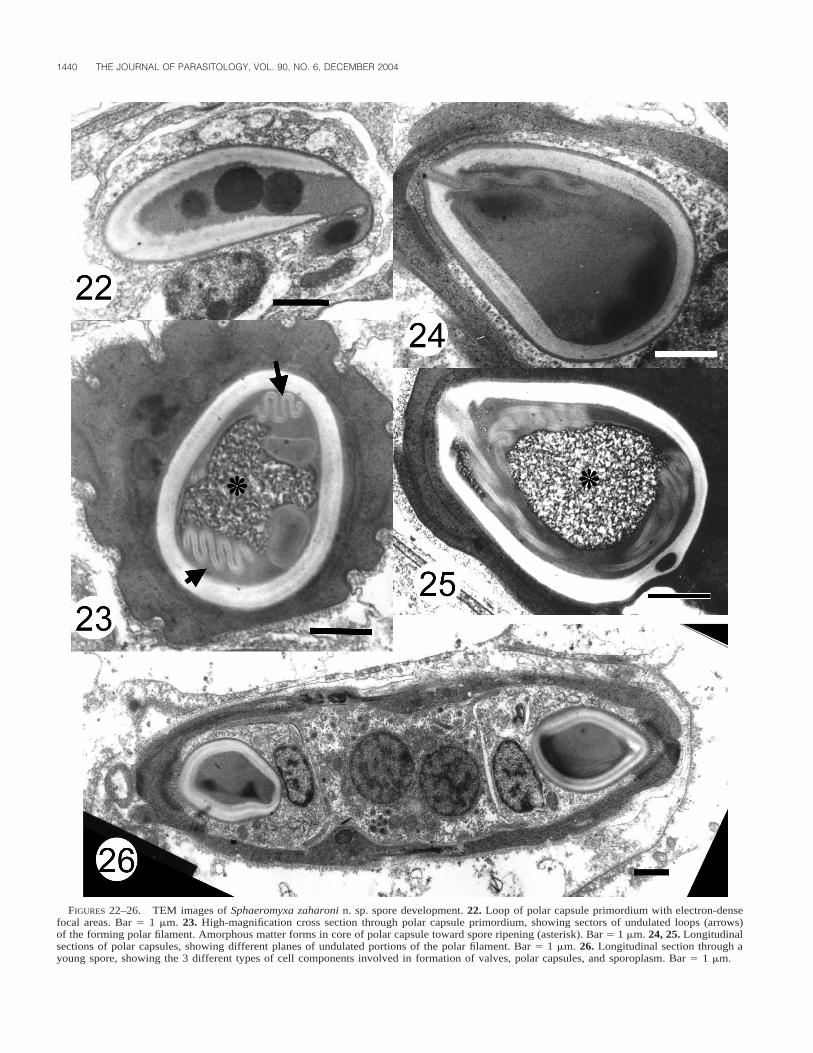
endoplasm zones observed with light microscope clearly evi-dent (Fig. 19). Endoplasm 10–12 mm in thickness, finely gran-ulated, and with no other notable feature. Beneath, endoplasmrich with small spherical or elongated mitochondria (dependingon the plane of sectioning), lipid inclusions, and generativecells. Adjoining underlying portions of endoplasm with alveolarappearance and comprising large compartments with small ar-eas of cytoplasm wedged in between. In some, aggregates ofvegetative nuclei, generative cells, pansporoblasts, or variousstages of developing spores (or all) observed (Figs. 20, 21, 26).In early sporoblasts, polar capsule primordium consisted ofmultiple cores of electron-dense matter and undulate sectionsof polar filament (Figs. 22–25). In ripe spores, polar capsuledeveloped core of amorphous matter (Figs. 23, 25) aroundwhich polar filament was wound. Longitudinal section throughyoung spore, showing details of the sporoplasm, polar capsules,and valves, shown in Figure 26.
1438 THE JOURNAL OF PARASITOLOGY, VOL. 90, NO. 6, DECEMBER 2004
FIGURES 14–18. Paraffin sections of Sphaeromyxa zaharoni n. sp. plasmodium stained with hematoxylin and eosin. 14–17. High-powermicrographs of plasmodium periphery, showing degrees of development of surface network complex. Note position of spores developing perpen-dicular to long axis of the plasmodium and phagocytotic activity of dark, granular matter (arrows). 17. Ripe spores near plasmodium surface,possibly in process of being discharged. Bar 5 5 mm. 18. High-power magnification of region of contact between plasmodium and gallbladderepithelium (arrows). Bar 5 10 mm.
Taxonomic summary
Type host: Devil firefish Pterois miles (Bennett, 1828).Locality: Eilat, Israel, northern Gulf of Eilat, Red Sea.Site of infection: Gallbladder.Type specimens: Deposited at the International Protozoan
Type Collection, National Museum of Natural History of theSmithsonian Institution, Washington, D.C. Syntype USNM1021225 (slide), Syntype USNM 1021226 (formalin).
Etymology: The species is named after the common name inHebrew of the type host, ‘‘zaharon.’’
Prevalence of infection: Eleven of 24 (45.8%).Additional host: Bearded scorpionfish Scorpaenopsis barbata
(Ruppell, 1838).Site of infection: Gallbladder; prevalence of infection: 2 of 4
(50.0%).
Remarks
There are 2 morphological types within Sphaeromyxa, i.e.,those with arcuate spores and pyriform polar capsules (‘‘incur-vata’’ group) and those with straight or slightly curved, fusi-form or ovoid spores with ovoid polar capsules (‘‘Balbiani’’group) (Laird, 1953). Our species is clearly a Balbiani type andis smaller than most other described species in the group. Theonly species whose range of spore measurements overlaps withours are Sphaeromyxa ovula Noble, 1939, from Gobiesox rhes-sodon inhabiting the California coast and Sphaeromyxa minutaPolyanski, 1955, from Hippoglossus hippoglossus inhabitingthe Barents Sea. However, the spores of both species differ; thespore of S. ovula is oval with rounded, narrow tips, and it hasa considerably smaller vegetative form. In S. minuta, the sporeis not widened in the middle and is nearly rectangular. Also, S.

DIAMANT ET AL.—S. ZAHARONI N. SP. FROM RED SEA DEVIL FIREFISH 1439
FIGURES 19–21. Transmission electron microphotographs of Sphaeromyxa zaharoni n. sp. (uranyl acetate and lead citrate). 19. Cross sectionthrough plasmodium periphery, ECT, ectoplasm; AE, alveolar endoplasm; MV, microvilli; Mt, mitochondria; L, lipid globules; GC, germinativecells. Bar 5 5 mm. 20. Central portion of the plasmodium, showing generative cells in an early stage of sporoblast formation. Bar 5 2 mm. 21.Cross section through 3 developing spores inside compartments within the alvelolar cytoplasm. In the sporoblast to the left, a loop of the polarcapsule lies adjacent to the polar capsule cell nucleus. The sporoblasts on right are developing in a single compartment; the sporoplasms arevisible in both. Note the forming ridges in the valve cells (small arrows). Bar 5 5 mm.
1440 THE JOURNAL OF PARASITOLOGY, VOL. 90, NO. 6, DECEMBER 2004
FIGURES 22–26. TEM images of Sphaeromyxa zaharoni n. sp. spore development. 22. Loop of polar capsule primordium with electron-densefocal areas. Bar 5 1 mm. 23. High-magnification cross section through polar capsule primordium, showing sectors of undulated loops (arrows)of the forming polar filament. Amorphous matter forms in core of polar capsule toward spore ripening (asterisk). Bar 5 1 mm. 24, 25. Longitudinalsections of polar capsules, showing different planes of undulated portions of the polar filament. Bar 5 1 mm. 26. Longitudinal section through ayoung spore, showing the 3 different types of cell components involved in formation of valves, polar capsules, and sporoplasm. Bar 5 1 mm.

DIAMANT ET AL.—S. ZAHARONI N. SP. FROM RED SEA DEVIL FIREFISH 1441
FIGURE 27. Single most parsimonious tree obtained from analysisof SSU rDNA sequence. Numbers at nodes indicate bootstrap confi-dence values.
minuta spores differ in that they lack ridges or striations. Bothparasitize fish from different host families and have differentzoogeographic distributions. Thus, we consider the currentlydescribed parasite as a new species.
Molecular analysis
In maximum parsimony (MP) analysis, S. zaharoni n. sp. wasbasal to clade containing a Myxidium species from mottled scul-pin (Cottus bairdii) and M. truttae with 100% bootstrap support(Fig. 28). On the basis of pairwise sequence comparisons (un-corrected p-distances), SSU rDNA sequence of S. zaharoni n.sp. was also most similar to that of M. truttae at 79.5% and tothat of a Myxidium sp. at 78.2%. Other phylogenetic methods(distance and maximum likelihood) largely supported MP anal-ysis, with minor differences in relationships that were not cen-tral to this study. For example, P. minibicornis was sister to Z.mugilis in MP analysis but sister to Kudoa species by distanceanalysis and sister to Ceratomyxa species by ML analysis. Re-gardless of these minor differences, there was strong supportfor 2 major groups, one containing species of Ceratomyxa, Ku-doa, Enteromyxum, Zschokkella, and Parvicapsula and the oth-er comprising Myxidium, Sphaerospora, Henneguya, Myxobol-us species, and S. zaharoni n. sp. (Fig. 27).
DISCUSSION
Of the 38 species of Sphaeromyxa so far described, aboutone-third parasitize families belonging to the Scorpaeniformes(including Cottidae, Liparidae, and Scorpaenidae) in zoogeo-graphically distant localities (see Lom, 2004). Our species isthe first to be described from the Red Sea. It is worth mention-ing that in a recent survey of myxosporeans of Red Sea coastalfish, no other case of Sphaeromyxa infection was found in anyof .160 host species examined, including several other scor-paenids (A. Diamant, pers. obs.).
The plasmodium of S. zaharoni n. sp. is typical of the veg-etative stages of the genus, aptly described as ‘‘large and leaf-like’’ by Laird (1953). Differentiation into ectoplasm and en-doplasm, as observed in this study, seems to be typical of coe-lozoic plasmodia, but not in all cases, e.g., pseudoplasmodia ofSphaerospora renicola Lom et al. (1982). The layer of amor-phous adhesive substance on the plasmodium outer membraneis probably produced and secreted by the parasite. Sphaeromyxa
zaharoni n. sp. has an asynchronous spore development, and itsfine structure is very similar to previously described species ofthe genus (see Lom, 1969, 2004; Grasse and Lavette, 1978;Gracia et al., 1997).
To the best of our knowledge, the budding process and plas-motomy observed in young, amoeboid plasmodial forms of thepresent species have not been previously reported in Sphaero-myxa, although this manner of division is known from othercoelozoic myxosporeans (Mitchell, 1978; Lom and Dykova,1995). In the species studied at present, there is an indicationthat ripe spores may be actively released by the plasmodiuminto the surrounding bile. To the best of our knowledge, activedischarge of spores from living plasmodia into the surroundingmedium is not known in Myxosporea. In K. thyrsites, a se-quential development occurs where intact plasmodia with ma-ture spores first appear, followed by spores released from themuscle fibers into the surrounding tissues and engulfed by mac-rophages (Moran et al., 1999). However, these authors were notable to determine whether the myxosporean plasmodium waslysed by the host response or whether the spores were releasedfrom the plasmodium by some other mechanism.
In our phylogenetic analyses, the SSU rDNA of S. zaharonin. sp. was most closely related to those of M. truttae and aMyxidium sp. Specifically, S. zaharoni n. sp. was basal to these2 Myxidium species with strong bootstrap support. With only asingle Sphaeromyxa representative in our analysis, it is difficultto come to any conclusions regarding the relationship of thisgenus to other myxozoan genera. Originally, this genus was amember of the Myxidiidae Thelohan 1892, which concurs withour finding of S. zaharoni n. sp. being sister to some Myxidiumspecies in our analysis. In the classification scheme proposedby Lom and Noble (1984), Sphaeromyxa was placed in a newsuborder (Sphaeromyxina) and family (Sphaeromyxidae) be-cause spores of this genus possess broad polar filaments, thusdistinguishing them from all other Myxozoa. Our analysis,therefore, raises the question whether the occurrence of ribbon-like polar filaments is a sufficiently distinct character to warranta unique suborder. At this time, however, we propose no changein the taxonomy of this group because our analysis includedonly 1 representative and we examined only 1 gene.
ACKNOWLEDGMENTS
This study was carried out within the framework of the Israel-JordanMiddle East Research Cooperation (MERC), Red Sea Marine PeacePark (RSMPP) project, funded by US-AID. We thank all our colleaguesat IUI, IOLR, and the Israel Nature and Parks Authority for their helpand support throughout the project.
LITERATURE CITED
FANTHAM, H. B., AND A. PORTER. 1912. Some effects of the occurrenceof myxosporidia in the gall bladder of fishes. Annals of Tropicaland Medical Parasitology 6: 467–481.
GRACIA, M. P., P. A. MAıLLO, J. M. AMIGO, AND H. SALVADO. 1997.Ultrastructural study of Sphaeromyxa balbiani Thelohan 1892 (My-xozoa, Myxosporea: Bivalvulida), a parasite of Cepola macro-phthalma Linnaeus, 1758. Acta Protozoologica 36: 171–179.
GRASSE, P. P., AND A. LAVETTE. 1978. La myxosporidie Sphaeromyxasabrazesi et le nouvel embranchement des myxozoaires (Myxozoa).Recherches sur l’etat pluricellulaire primitive et considerations phy-logenetiques. Annales des Sciences Naturelles, Zoologie, Paris 12:193–285.
KENT, M. L., K. B. ANDREE, J. L. BARTHOLOMEW, M. EL-MATBOULI, S.

1442 THE JOURNAL OF PARASITOLOGY, VOL. 90, NO. 6, DECEMBER 2004
S. DESSER, R. H. DEVLIN, S. W. FEIST, R. P. HEDRICK, R. W. HOFF-MANN, J. KHATTRA, S. L. HALLETT, R. J. G. LESTER, M. LONGSHAW,O. PALENZEULA, M. E. SIDDALL, AND C. X. XIAO. 2001. Recentadvances in our knowledge of the Myxozoa. Journal of EukaryoticMicrobiology 48: 395–413.
LAIRD, M. 1953. The protozoa of New Zealand intertidal zone fishes.Transactions of the Royal Society of New Zealand 81: 79–143.
LOM, J. 1969. Notes on the ultrastructure and sporoblast developmentin fish parasitizing myxosporidian of the genus Sphaeromyxa. Zeit-schrift Zellforschung 97: 416–437.
———. 2004. Morphology and ultrastructure of Sphaeromyxa nobleisp. n. (Myxozoa), parasite of Heteroclinus whiteleggii (Pisces) fromAustralian New South Wales coast. Folia Parasitologica 51: 19–26.
———, AND P. DE PUYTORAC. 1965. Studies on the myxosporidian ul-trastructure and polar capsule development. Protistologica 1: 53–65.
———, AND I. DYKOVA. 1992. Protozoan parasites of fishes. Elsevier,Amsterdam, The Netherlands, 315 p.
———, AND ———. 1995. Myxosporea (Phylum Myxozoa). Fish dis-
eases and disorders, vol. 1. Protozoan and metazoan infections.CAB International, Wallingford, U.K., p. 97–148.
———, AND E. R. NOBLE. 1984. Revised classifications of class My-xosporea Butschli, 1881. Folia Parasitologica (Prague) 31: 193–205.
MITCHELL, L. J. 1978. Myxosporidian infections in some fishes of Iowa.Journal of Protozoology 25: 100–105.
MORAN, J. D. W., L. MARGOLIS, J. M. WEBSTER, AND M. L. KENT. 1999.Sequential development of Kudoa thyrsites (Myxozoa: Myxospo-rea) in netpen-reared Atlantic salmon determined by light micros-copy and a polymerase chain reaction test. Diseases of AquaticOrganisms 37: 185–193.
POSADA, D., AND K. A. CRANDALL. 1998. Modeltest: Testing the modelof DNA substitution. Bioinformatics 14: 817–818.
SWOFFORD, D. L. 1998. PAUP*: Phylogenetic analysis using parsimony(*and other methods), version 4. Sinauer Associates, Sunderland,Massachusetts.
THOMPSON, J. D., T. J. GIBSON, F. PLEWNIAK, F. JEANMOUGIN, AND D. G.HIGGINS. 1997. The Clustal X windows interface: Flexible strate-gies for multiple sequence alignment aided by quality analysistools. Nucleic Acids Research 24: 4876–4882.










