
ข้อมูลสรุปสาหรับผู้บริหาร (Executive Summary)...
156
7 ข้อมูลสรุปสาหรับผู้บริหาร (Executive Summary) โครงการ Molecular characterization of genotypes, virulence factors, gene transfer and copper transport system of Acinetobacter baumannii โดย ผศ.ดร. สุทธิรัตน์ สิทธิศักดิ์ ธันวาคม พ.ศ. 2560
-
Upload
khangminh22 -
Category
Documents
-
view
3 -
download
0
Transcript of ข้อมูลสรุปสาหรับผู้บริหาร (Executive Summary)...

7
ขอมลสรปส าหรบผบรหาร (Executive Summary)
โครงการ Molecular characterization of genotypes, virulence factors, gene transfer
and copper transport system of Acinetobacter baumannii
โดย ผศ.ดร. สทธรตน สทธศกด
ธนวาคม พ.ศ. 2560

8
1. ความส าคญและทมาของปญหา
A. baumannii เปนแบคทเรยทมความรนแรงในการกอโรคและพบเปนสาเหตของการตดเชอในผปวยทพก
รกษาตวในหออภบาลผปวยวกฤต (ICU) เปนเวลานาน (nosocomial infection) เมอผปวยมการตดเชอจากแบคทเรย
A. baumannii สายพนธทมการดอตอยาหลายชนด ท าใหอตราการเสยชวตเพมสงขน ในป 2557 จากการศกษาของ
Inchai และคณะ พบการระบาดของเชอ A. baumannii ดอยาในประเทศไทยอยท รอยละ 65.3 (Inchai 2015) การ
รายงานจากการตดตามเฝาระวงสถานการณการดอยาของเชอแบคทเรยของกรมวทยาศาสตรการแพทย ไทยพบวาในป
2516 พบ carbapenem-resistance A. baumannii (CR-AB) ในอตรา 67.2 % เพมจากป 2006 ทพบเพยง 38.5%
โดยเฉพาะในผ ป วยหอวกฤต (ICU) พบว าแยกเช อ A. calcoaceticus-baumannii complex ท ด อยากล ม
carbapenem ในอตราสงถง 80 % A. baumannii มความสามารถในการสรางปจจยกอโรคไดหลากหลายแบบเชน
การสรางไบโอฟลม ซงมผลท าใหยาปฏชวนะไมสามารถท าลายเชอและรกษาโรคตดเชอท เกดจากตวเชอนได การ
สรางฮโมไลซน เปนสารพษทท าใหเซลลเมดเลอดแดงของเซลลโฮสตแตกและเกดพยาธสภาพภายในรางกายของผปวย
การสรางอะซนโตแบคตน เปนโปรตนทจ าเพาะกบธาตเหลก ตวเชอสรางขนเพอแขงขนในการจบกบธาตเหลกในรป
Fe3+ ในเซลลโฮสต มผลท าใหเซลลโฮสตแตกและท างานผดปกต เปนตน นอกจากนยงพบวามมกลไกดอยาหลาย
แบบทส าคญคอการสรางเอนไซมในกลมบตาแลคทาเมส (beta-lactamase) เพอมาท าลายยาปฏชวนะในกลมบตาแล
คแตม ซงเปนยาปฏชวนะทนยมใชอยางแพรหลายในการรกษาโรคตดเชอ เอนไซมในกลมบตาแลคทาเมสทเชอสรางขน
สามารถแบงไดเปน 4 class คอ class A, B, C และ D ยนดอยาท พบใน CR-AB สวนใหญเปนยนดอยาในกลม class
D β-lactamase ในประเทศไทยมรายงานการแพรกระจายของยนดอยา class D β-lactamase อยเกอบทกภมภาค
การแพรกระจายของยนดอยา มกเกดจากการถายทอดยนดอยาในแนวราบ (horizontal gene transfer) โดยเปนการ
ถายทอดยนระหวางแบคทเรยชนดหนงไปสแบคทเรยอกชนดหนง โดยอาศยกระบวนการทส าคญ คอ ทรานสฟอรเมชน
ทรานสดกชน และคอนจเกชน มรายงานการศกษาพบวาในเชอ Acinetobacter spp. กระบวนการทรานสฟอรเมชน
สามารถถายทอดยนดอยา blaOXA-23, blaOXA-24, blaOXA-58 และ blaNDM-1ไปสแบคทเรยผรบได ท าใหเกดการ
แพรกระจายของยนดอยาภายในโรงพยาบาลเพมสงขน นอกจากนยงพบการถายทอดยนดอยา blaOXA-23, blaOXA-58,
blaGES-11, blaIMP-1 และ blaNDM-1 จาก A. baumannii ไปสแบคทเรยผรบ โดยอาศยกระบวนการคอนจเกชน

9
คอปเปอรเปนสารอาหารทจ าเปนตอการด ารงชวตส าหรบเซลลแบคทเรย แต ต อ งกา ร ในปรมาณนอย
(Trace element) รปแบบของคอปเปอรทพบม 2 รปแบบคอ Cu(II) ซงเปนรปแบบทใชรบอเลกตรอนจากสารตว
อน (Oxidize form) และ Cu(I) ทเปนตวใหอเลกตรอนแกสารตวอน (Reduce form) โดยคอปเปอรทพบในธรรมชาต
สวนใหญจะอยในรปแบบ Cu(II) แตเซลลแบคทเรยจะน าคอปเปอรเขาสเซลลในรปแบบ Cu(I) อยางไรกตามคอปเปอร
จะเปนพษตอเซลลแบคทเรยเมอเซลลสมผสคอปเปอรในปรมาณสง ดงนนแบคทเรยจงตองมกลไกในการควบคม
สมดลคอปเปอรในเซลล มการศกษาทแสดงใหเหนวาหากมการน ายนความตานทานตอคอปเปอรออกไปจากเซลล
แบคทเรยกอโรค จะท าใหความรนแรงของการกอโรคลดลงเชนสญเสยการสรางไบโอฟลม stress tolerance และ
genetic transformation (Singh et al., 2015) สงเหลานชใหเหนวาความตานทานคอปเปอรตอมความส าคญอยาง
ยงส าหรบการรอดชวตของแบคทเรยทงในสภาพแวดลอมทปนเปอนคอปเปอร และเกยวของกบความรนแรงและการ
กอโรคของแบคทเรย แบคทเรยจะมระบบควบคมสมดลคอปเปอรเพอชวยปองกนพษจากคอปเปอรท าใหแบคทเรย
รอดชวตในสภาวะทมคอปเปอรสง ดวยกลไกทแตกตางกนหลายกลไกเชน efflux pump หรอ detoxifying enzyme
(Marie et al., 2012; Erik & Michael, 2015) ปจจปนกลไกการทนตอคอปเปอรไดรบการศกษาโดยละเอยดใน
แบคทเรยแกรมลบ เชน Escherichia coli และแบคทเรยแกรมบวกเชน Enterococcus hirae สวนการศกษาระบบ
ควบคมสมดลคอปเปอรเปนหนงในกลไกทจ าเปนในเชอ A. baumannii ในการเจรญ พบวายงมการศกษานอย
ดงนนในการศกษาครงนจงมวตถประสงคเพอศกษารปแบบการดอตอยาตานจลชพ ตรวจหายนดอยาและ
virulence gene ศกษาการถายทอดยนดอยาในแนวราบ ไดแกกระบวนการทรานสฟอรเมชน ทรานสดกชน และ คอน
จเกชน ศกษารปแบบการระบาดของเชอ A. baumannii ทดอยาแบบรนแรง (XDR-AB) โดยวธ Repetitive-PCR
(REP-PCR) วธ blaOXA-51 sequence base typing (blaOXA-51 SBT) และวธ Multilocus sequence typing
(MLST) รวมทงศกษาระบบควบคมสมดลคอปเปอรของเชอ A. baumannii ทแยกไดจากผปวยและทแยกไดจาก
ธรรมชาต ซงการศกษาครงนจะน ามาซงขอมลเบองตนเพอทน าไปใชในการปองกนการแพรกระจายของเชอดอยาและ
พฒนาวธการรกษาการตดเชอ A. baumannii ตอไป

10
2. วตถประสงค
1) เพอศกษา virulence factors และ virulence gene ของเชอ A. baumannii clinical isolates
2) เพอศกษารปแบบการดอยาและยนดอยาของเชอ A. baumannii clinical isolates
3) เพอแยกเชอ A. baumannii จากสงแวดลอมรวมทงศกษารปแบบการดอยาและยนดอยาของเชอทแยกได
4) เพอศกษารปแบบการระบาดของเชอ Extensively-drug resistance Acinetobacter baumannii ดวยเทคนค
ทางอณชววทยา
5) เพอศกษาการถายทอดยนดอยาในแนบราบ โดยวธทรานสฟอรเมชน, ทรานสดกชน และคอนจเกชนจาก
Extensively-drug resistance Acinetobacter baumannii ไปยง A. baumannii ทแยกไดจากสงแวดลอม
6) เพอศกษาระบบควบคมสมดลคอปเปอรของเชอ A. baumannii
3. ระเบยบวธวจย
1) ศกษา virulence factors และ virulence ของเชอ A. baumannii clinical isolates
- ตรวจหา virulence genes ไดแกยน จ านวน 5 genes ไดแก bap, ompA, epsA, bauD-bauC (Acinetobactin )
และ hly (hemolysin) โดยวธ PCR
-ศกษาการสรางไบโอฟลมโดยวธ microtiter plate assay การศกษาการสรางฮโมไลซน โดยดการยอยสลายเมดเลอด
แดงบน Blood agar plate ศกษาการเจรญของเชอ A. baumannii บนอาหารทม iron จ ากดและการสราง
siderophore (อาหาร Chrome Azurol S agar (CAS agar))
2) เพอศกษารปแบบการดอยาและยนดอยาของเชอ A. baumannii clinical isolates
-ทดสอบความไวตอยาตานจลชพของเชอ A. baumannii clinical isolates โดยวธ disc diffusion
-ตรวจหายนดอยา OXA-type carbapenamase (blaOXA-23, blaOXA-24, blaOXA-58 and intrinsic blaOXA-
51) ดวยเทคนค Multiplex PCR ในเชอ A. baumannii clinical isolates
-ตรวจหายนดอยาในกลมบตาแลคทาเมส (Beta-lactamase) ทง 4 class (Class A, B, C, D) จ านวน 17 genesใน
เชอ Extensively-drug resistance Acinetobacter baumannii
3) เพอแยกเชอ A. baumannii จากสงแวดลอมรวมทงศกษารปแบบการดอยาและยนดอยาของเชอทแยกได

11
-ทดสอบความไวตอยาตานจลชพของเชอ A. baumannii โดยวธ disc diffusion
-ตรวจหายนดอยา OXA-type carbapenamase (blaOXA-23, blaOXA-24, blaOXA-58 and intrinsic blaOXA-
51) ดวยเทคนค Multiplex PCR
4) เพอศกษารปแบบการระบาดของเชอ Extensively-drug resistance Acinetobacter baumannii ดวยเทคนค
ทางอณชววทยา
-ศกษารปแบบการระบาดของเชอ XDR-AB 27 ไอโซเลทดวยวธ Repetitive-PCR (REP-PCR) วธ blaOXA-51
sequence base typing (blaOXA-51 SBT) และวธ Multilocus sequence typing (MLST)
5) เพอศกษาการถายทอดยนดอยาในแนบราบ โดยวธทรานสฟอรเมชน, ทรานสดกชน และคอนจเกชนจาก
Extensively-drug resistance Acinetobacter baumannii ไปยง A. baumannii ทแยกไดจากสงแวดลอม
6) เพอศกษาระบบควบคมสมดลคอปเปอรของเชอ A. baumannii
- ศกษาคา Minimum inhibitory concentration (MIC) ของคอปเปอรของเชอ A. baumannii clinical isolates
และ A. baumannii ทแยกไดจากสงแวดลอม ดวยวธ agar dilution method
-ตรวจหา copper resistance determinants ของ A. baumannii โดยวธ PCR
-ศกษาการแสดงออกของ copper associated genes เมอถกกระตนดวยโลหะหนกคอปเปอร ท าโดยใชวธ Real-
time qPCR assays
-ศกษาหนาทของยน 2 ยนทเกยวของกบการควบคมสมดลคอปเปอรโดยการท า gene deletion mutant
4. ผลงานวจย
ผลการศกษาในเชอ A. baumannii จากโรงพยาบาลในประเทศไทย 4 แหง และโรงพยาบาลในประเทศเนปาล
1 แหงพบรปแบบการดอตอยาตานจลชพในกลมคารบาพเนม (CR-AB) รอยละ 65-98 กลมดอยาหลายขนาน รอยละ 75-
98 กลมดอยาแบบรนแรง (XDR-AB) รอยละ 2-21 การตรวจหายนดอยาในกลม OXA-type carbapenemase ตรวจ
พบยนดอยา blaOXA-23 มากทสด รองลงมาไดแก ยนดอยา blaOXA-58 ตามดวยยน blaOXA-24 การตรวจหายนดอยาใน
กลม metallo beta-lactamase ตรวจพบ blaNDM-1 รอยละ 8-18 การตรวจหา virulence gene ในเชอ A. baumannii
พบยน hly (90-98%) ทความชกสงสดรองลงมาคอยน ompA (74-90%) และยน BauD-bauC (2.8-5.5%) พบต าสด
โดยการมยน ompA มความสมพนธกบเชอทมรปแบบการดอยาแบบ XDR-AB การศกษาการถายทอดยนดอยาใน

12
แนวราบ โดยอาศยกระบวนการทรานสฟอรเมชน ทรานสดกชน และ คอนจเกชน โดยใชเชอ XDR-AB เปนแบคทเรยผให
Acinetobacter spp. ซงแยกไดจากแหลงน าธรรมชาตทมคณสมบตไวตอยาตานจลชพทกชนดและไมมยนดอยาเปน
แบคทเรยผรบพบวาเชอ XDR-AB สามารถสงผานการดอยา kanamycin, tetracycline และ ticarcillin ไปสแบคทเรย
ผรบได สามารถตรวจพบยนดอยา tet(B) ซงท าใหเชอดอยา tetracycline ในแบคทเรยผรบของกระบวนการ ทรานส
ดกชน สามารถตรวจพบยนดอยา blaPER-1 ซงท าใหเชอดอยา ticarcillin ในแบคทเรยผรบของกระบวนการคอนจเกชน
และตรวจพบยนดอยา aphA6 ซงท าใหเชอดอยา kanamycin ในแบคทเรยผรบของกระบวนการคอนจเกชน การศกษา
ในเชอ XDR-AB ทแยกไดจากประเทศไทยทงหมด 27 ไอโซเลท เพอตรวจหายนดอยากลมเบตา-แลคตาเมสทง 4 classes
พบยนดอยาใน class A เฉพาะยน blaPER-1 และ class B ตรวจพบเพยงยน blaNDM-1 ยนดอยาใน class C ตรวจพบ
เฉพาะยน blaADC และยนดอยาใน class D ตรวจพบยน blaOXA-23 (100%) และ blaOXA-51 (100%) การศกษารปแบบ
การระบาดทางอณชววทยาโดยวธ REP-PCR ในการศกษานสามารถจดรปแบบการระบาดไดทงหมด 4 รปแบบ คอ
รปแบบ 1, 4, 15 และ 16 การศกษารปแบบการระบาดโดยวธ blaOXA-51 SBT ในการศกษาครงนพบรปแบบการระบาด
ได 4 กลมแบบ ไดแก OXA-64, OXA-83, OXA-104 และ OXA-508 และจากการจดกลมการระบาดโดยวธ Multilocus
sequence typing (MLST) พบเชอ XDR-AB ทงหมด 27 ไอโซเลท สามารถจดกลมเปน STs ได 23 ไอโซเลท และอก 4
ไอโซเลททไมถกจดอยใน STs ใดเลย (non-type) เชอ 23 ไอโซเลท ถกจดอยใน STs ทมความหลากหลาย ไดแก ST195,
ST208, ST368, ST451, ST457 และ ST1166 ผลการศกษาระบบควบคมสมดลคอปเปอรของเชอ A. baumannii จาก
whole genome sequence ของเชอ A. baumannii ทมอยใน GeneBank พบวามระบบควบคมสมดลของคอปเปอร
(cop operon) ทงหมด 3 ระบบคอระบบ cueR system ระบบ copRS system และ pCo-homolog system ซงแต
ละระบบจะควบคมการแสดงออกของยนทแตกตางกน เชอ A. baumannii ทแยกจากผปวยและแยกไดจากธรรมชาต
ตรวจพบระบบ cueR ทกสายพนธ (100%) พบระบบ copRS ในเชอทแยกไดผปวย 18.6% และ ในเชอทแยกจาก
ธรรมชาต 16.6 % โดยวธ PCR โดยเชอสวนใหญทมระบบควบคมสมดลของคอปเปอร (cop operon) ทง 2 ระบบจะม
คาความตานทานตอคอปเปอรสง (MIC เทากบ 10 mM) การแสดงออกของ copper resistance determinants ในทง
สามระบบถกชกน าโดยโลหะคอปเปอรและเมอท าการ mutate ยน copA ซง encoded multicopper oxidase จาก
CopRS operon และยน actP1 ซง encoded copper ATPase จากCue operon ในเชอ A. baumannii สายพนธ
AB 1521 จะพบวาแบคทเรยจะม copper sensitivity ลดลง

13
ผลงาน/หวขอเรองทได
1. เขารวมน าเสนอผลงานในทประชมวชาการระดบนานาชาตและระดบชาต ทมการตพมพเฉพาะ Abstract ใน
งานประชมวชาการ ISAAR (International Symposium on Antimicrobial Agents and Resistance) & ICIC
(International Interscience Conference on Infection and Chemotherapy) ในวนท 14-16 กนยายน 2560 ณ
เมองปซาน ประเทศเกาหล และงานสมนาระบาดวทยาแหงชาต ครงท 23 ภายใตการประชมวชาการเครอขายระดบ
โลกครงท 9 ระหวางวนท วนท 7-11 สงหาคม 2560 ณ โรงแรมดเอมเพลส จงหวดเชยงใหม
2. ตพมพในวารสารระดบนานาชาต 3 เรอง
2.1 Rapee Thummeepak, Phattaraporn Kongthai, Udomluk Leungtongkam, Sutthirat Sitthisak.
(2016). Distribution of virulence genes involved in biofilm formation in multi-drug resistant
Acinetobacter baumannii clinical isolates. International Microbiology, 19(2):121-129.
doi:10.2436/20.1501.01.270
2.2 Prabhu Raj Joshi, Mahesh Acharya, Trishna Kakshapati, Udomluk Leungtongkam, Rapee
Thummeepak and Sutthirat Sitthisak. (2017). Co-existence of bla OXA-23 and bla NDM-1 genes of
Acinetobacter baumannii isolated from Nepal: antimicrobial resistance and clinical significance.
Antimicrobial Resistance and Infection Control, 6(21):1-7. doi: 10.1186/s13756-017-0180-5.
2.3 Leungtongkam U, Thummeepak R, Wongprachan S, Thongsuk P, Kitti T, Ketwong K,
Runcharoen C, Chantratita N, Sitthisak S. (2017). Dissemination of blaOXA-23, blaOXA-24, blaOXA-58,
and blaNDM-1 Genes of Acinetobacter baumannii Isolates from Four Tertiary Hospitals in Thailand.
Microbial Drug Resistance, Jun 8. doi: 10.1089/mdr.2016.0248.
4. เตรยม manuscript เพอตพมพในวารสารระดบนานาชาต 3 เรอง
4.1 Udomluk Leungtongkam, Rapee Thummeepak, Kannipa Tasanapak and Sutthirat Sitthisak.
Acquisition and transfer of antibiotic resistance genes and integrons of Acinetobacter baumannii by
conjugation.

14
4.2 Phattaraporn Kongthai, Udomluk Leungtongkam, Rapee Thummeepak, Thawatchai kitti ,
Aunchalee Thanwisai, Narisara Chantratita, Sutthirat Sitthisak. Molecular epidemiology and virulence
gene profiles of extensively drug-resistant Acinetobacter baumannii from Thailand.
4.3 Rapee Thummeepak, Aunchalee Thanwisai. Narissara Chantratita and Sutthirat Sitthisak. Molecular
characterization of copper resistance determinants of Acinetobacter baumannii.
5. นสตปรญญาโท 2 ราย
5.1 นางสาวอดมลกษณ เหลองทองค า หวขอวทยานพนธเรอง การศกษาคณสมบตทางอณชววทยาของคลาสด
บตา-แลคทาเมส และการถายทอดยน ในเชอAcinetobacter baumannii
5.2 นางสาว ภทรพร คงไทย หวขอวทยานพนธเรอง ลกษณะทางอณชววทยาของ extensively drug-resistant
Acinetobacter baumannii ทแยกไดจากโรงพยาบาลในประเทศไทย
6. นสตปรญญาเอก 1 ราย หวขอวทยานพนธเรอง
6.1 นายระพ ธรรมมภกด หวขอวทยานพนธเรอง Molecular characterization of copper homeostasis
systems in Acinetobacter baumannii

15
บทท 1
การศกษา virulence factors ของ A. baumannii
clinical isolates
สวนหนงของการศกษาไดรบการตพมพในวารสาร International Microbiology.
Rapee Thummeepak, Phattaraporn Kongthai, Udomluk Leungtongkam, Sutthirat Sitthisak. 2016.
Distribution of virulence genes involved in biofilm formation in multi-drug resistant Acinetobacter
baumannii clinical isolates. International Microbiology. 19(2):121-129.

16
บทน า (Introduction)
Acinetobacter baumannii เปนแบคทเรยอยใน family Moraxellaceae genus Acinetobacter ม
ลกษณะเปนแกรมลบ ไมเคลอนท ไมสรางแคปซล ไมสรางสปอร ไมหมกน าตาลกลโคส และเจรญในสภาวะมออกซเจน ซง
สามารถพบไดในดน น า และยงเปน normal flora ทสามารถเตบโตไดในรางกายทงผวหนง คอหอย เสมหะ ปสสาวะ
และอจจาระ อาจพบไดในเยอเมอกสารคดหลงตางๆ และอาจพบไดทวไปในสงแวดลอมของโรงพยาบาลและผปวยทนอน
พกรกษาตวในโรงพยาบาลเปนเวลานานซงเปนสาเหตทส าคญในการตดเชอในโรงพยาบาล (nosocomial infection)
เชอ A.baumannii สามารถกอโรคไดหลายระบบ ทงปอดอกเสบ เยอหมสมองอกเสบ การตดเชอในกระแสเลอด แผล
ผาตด เยอบชองทองอกเสบ เยอหมหวใจอกเสบ ตดเชอในสายสวนหลอดเลอดและตดเชอทางเดนปสสาวะซงลวนเปนโรค
ทเกดไดในผปวยวกฤต และอตราการตายดวยปอดอกเสบจากเชอ A. baumannii นมคอนขางสง การตดเชอ A.
baumannii ในโรงพยาบาลยงเปนปญหาทส าคญของผปวยทพกรกษาตวในโรงพยาบาลทวโลกรวมทงในประเทศไทย
โดยสาเหตอนเนองมาจากเชอชนดนมการดอตอยาตานจลชพหลายชนด จงพบผปวยทมอตราการตายสงจากการตดเชอ
A. baumannii โดยมรายงานการดอยาตานจลชพของ เชอ A. baumannii ในประเทศไทยทแยกไดจากสงสงตรวจ
ของผปวยในโรงพยาบาลมหาราชนครเชยงใหม พบวาการดอยาตานจลชพของเชอนเพมจากรอยละ 46 ใน ป พ.ศ. 2546
เปนรอยละ 65.3 ในป พ.ศ. 2558 (Chaiwarith et al, 2005; Inchai et al, 2015) ผปวยทมการตดเชอเพมสงขนในแต
ละป ท าใหเกดการเสยชวตและเสยคาใชจายจ านวนมากในการรกษาผปวยทตดเชอ
การกอโรคของเชอ A. baumannii และความรนแรงของโรค ขนอยกบปจจยตางๆ ทงตวมนษยและตว
เชอ โดยเฉพาะผปวยทไดรบการรกษาในโรงพยาบาลเปนเวลานานและผปวยทมภาวะภมคมกนบกพรอง และA.
baumannii สายพนธทกอโรครนแรงมกพบเกยวของกบการมปจจยกอโรค (virulence factors) จากตวเชอ โดย
virulence factors ทส าคญๆ แสดงดงในภาพ 1

17
ภาพ 1 แสดงปจจยกอโรคของเชอ A. baumannii
เชอ A. baumannii จะมการสราง virulence factor ตางๆ เชน Outer membrane (OmpA), Capsular
polysaccharide (PTK และ EpsA), Acinetobactin, การสรางไบโอฟลม (Biofilm), การสรางฮโมไลซน(Hemolysin)
ซง virulence factor เหลานเปนปจจยส าคญทท าใหเชอ A. baumannii มความรนแรงในการกอโรคมากขน ดงน
1. Outer membrane vesicles (OMVs) เปนถงทเชอสรางจากโครงสรางหอหมเซลลสวน outer membrane
โดยขางในถงจะมการบรรจสารมหโมเลกล เชน OmpA ซงเปนโปรตนทถกควบคมการสรางโดยยน ompA ท าหนาทเปน
protease และ hemolysin เปนกลไกทเชอใชปองกนการตอบสนองทางภมคมกนของมนษย นอกจากนนในถงดงกลาว
ยงมการบรรจสารพนธกรรม ท าใหเกดการสงตอยนตางๆ ไปยงเซลลแบคทเรย (horizontal gene transfer) การศกษา
ของ Choi และคณะพบวา OmpA ท าใหเกดการตายแบบ apoptosis ใน laryngeal epithelial cell ซงเปนเซลลไลน
ของมนษยโดยท OmpA สามารถเคลอนเขาไปในไมโตคอนเดรย กระตนใหมการหลง cytochrome c และ apoptosis
inducing factor แลวเหนยวน าใหเกด apoptosis ของเซลลไลน จงสรปไดวา OmpA สามารถท าใหเกดการบาดเจบของ
เซลลบรเวณทางเดนหายใจในระหวางการตดเชอ (Choi et al., 2005; 2008)

18
2. capsular polysaccharide เปนสารจ าพวก polysaccharide มลกษณะเปนชนและมเมอกลอมรอบผนงเซลล
ชวยปองกนตวเชอจากการท าลายโดยระบบภมคมกน โดยตวเชอจะไมถกจบกนโดย phagocytes นอกจากนแคปซลท
เชอสรางขนยงท าใหการรกษาผปวยเปนไปไดยาก เนองจากยาปฏชวนะทใชในการรกษาไมสามารถซมผานแคปซล และ
ท าลายเชอได ยนทควบคมการสรางแคปซล ไดแก ptk (protein tyrosine kinase) และ epsA (McConnell, Actis &
Pachon, 2012; Russo, et al., 2010)
3. Biofilm เปนการอยรวมกนของแบคทเรยทมการสรางสารโพลเมอรจ าพวก polysaccharide ไดแก แคปซล
(capsule) หรอสารเมอก (slime) และปลอยออกมานอกเซลล เพอยดเกาะทผวของวตถ ไบโอฟลมชวยปองกนแบคทเรย
ทอาศยอยภายในได การสรางไบโอฟลมของเชอพบบนสายสวนปสสาวะของผปวย ท าใหผปวยตดเชอในระบบทางเดน
ปสสาวะ ท าใหยาปฏชวนะไมสามารถซมผานไบโอฟลมและออกฤทธท าลายเชอทอยในไบโอฟลมได การสรางไบโอฟลม
ของ A. baumannii ถกควบคมโดยยน bap, bfmR และ bfmS นอกจากนการสราง pilus ยงถกควบคมโดย
CsuA/BABCDE operon มการศกษาพบวา การท าใหยน bfmR ไมแสดงออกมผลตอ CsuA/BABCDE operon ท าใหไม
มการสราง pilus และไมมการสรางไบโอฟลม (McConnell et al., 2012) นอกจากนการสรางไบโอฟลมยงถกควบคม
โดยยนอน ไดแก ยน epsA และยน bap โดยยน epsA เปนยนทเกยวของกบการสราง exopolysaccharide capsule
ซงเปนองคประกอบหลกในการสรางไบโอฟลม (Russo, et al., 2010) สวนยน bap (Biofilm-Associated Protein) เปน
ยนทชวยในการสราง biofilm และชวยใหเชอยดเกาะตอพนผวในแบบตางๆ ได ทงพนผวทมชวตและไมมชวต เชน วสดท
ท าจาก polypropylene และ polystylene เปนตน (Goh et al., 2013)
4. Acinetobactin เปนโปรตนชนดหนงทมหนาทในการจบกบ Fe3+ เพอใหตวเชอใชในการเจรญและการ
ด ารงชวตในสภาวะทมปรมาณออกซเจนต า ซง Fe3+ มความจ าเปนในขนตอนการ replication ของเซลลมผลท าใหการ
สรางสารตางๆในเซลลโฮสตมความผดปกต ยนทเกยวของในการสงเคราะห acinetobactin คอ basA, basB, basC,
basD, basF, basG, basH, basI และ basJ สวนยนทควบคมการขนสง ferric-acinetobactin complexes ไดแก
bauB, bauC, bauD และ bauE จากการศกษาของ Gaddy และคณะ พบวาการกลายพนธของยน bauA และ bauD
มผลท าใหความสามารถในการกอโรคและความสามารถในการท าใหเซลลแตกของเชอลดลงใน epithelial cell ของเซลล
โฮสต (Gaddy, et al., 2012; McConnell et al., 2012; Hasan, Choi & Oh, 2015)

19
5. Hemolysin เปนสารพษ (toxin) ทสรางขนโดยแบคทเรยหลายชนด บทบาทของ hemolysin คอ การท าใหเกด
ร (pore-forming) บรเวณเยอหมเซลลของเซลลหลายชนด ไดแก เซลลเมดเลอดแดง (erythrocyte), monocytes,
neutrophil และ macrophage เมอเมดเลอกแดงแตกจะมการปลดปลอย heme ซงเปนแหลงของธาตเหลก และเปน
สารทจ าเปนตอแบคทเรยในขนตอนการ replication มการศกษาการยอยเมดเลอดแดงของ A. baumannii พบวา A.
baumannii เพยงบางไอโซเลทเทานนทสามารถยอยเมดเลอดแดงได และบางการศกษาพบวาไมเกดการยอยเมดเลอด
แดงแกะ แตสามารถยอยเมดเลอดแดงมาได ยนทเปนตวก าหนดการสราง hemolysin คอ ยน hly (Antunes, et al.,
2011)
การศกษาครงน เปนการศกษาคณสมบตในการสรางไบโอฟลม และการสรางฮโมไลซน และการสราง
siderophore ของเชอ A. baumannii รวมทงตรวจหายนทเกยวของกบการสรางไบโอฟลม การสรางฮโมไลซนและยน
ทเกยวของในการสงเคราะห acinetobactin ในเชอ A. baumannii ทแยกจากโรงพยาบาล 3 แหงในประเทศไทย
การศกษานสามารถท าใหทราบเกยวกบ virulence factor ของเชอ A. baumannii ทมการแพรระบาดในประเทศไทย
และยงน าขอมลไปประยกตใชเพอชวยพฒนาใชรกษาโรคทเกดจากเชอ A. baumannii ตอไปได
20
วธด าเนนการวจย (Material and Method)
1. แบคทเรยทใชในการศกษาวจย
เชอทใชศกษาคอ เชอ A. baumannii ทไดมาจากตวอยางผปวยในโรงพยาบาลพจตรจ านวน 55 ตวอยาง โรงพยาบาล
สวรรคประชารกษจ านวน 66 ตวอยาง โรงพยาบาลเชยงรายประชานเคราะหจ านวน 109 ตวอยาง รวมทงสน 230
ตวอยาง และเชอ Acinetobacter spp. จ านวน 19 ไอโซเลตทแยกจากแหลงน าธรรมชาตภายในมหาวทยาลยนเรศวร
และโรงพยาบาลพทธชนราช
2. การทดสอบความไวตอยาตานจลชพโดยวธ disc diffusion
เพาะเลยงเชอแบคทเรย A. baumannii บนจานอาหารเลยงเชอ TSA แลวน าไปบมท 37 องศาเซลเซยส เปนเวลา 24
ชวโมง จากนนน าโคโลนทเจรญบนจานอาหารเลยงเชอมาปรบความขนในสารละลาย 0.85%NaCl ใหไดความขน
เทยบเทากบ 0.5 Mcfarland โดยใชเครองวดความขน (McFarland densitometer: SiaBiosan, Latvia) น าไมพนส าล
ทปราศจากเชอจมลงในเชอทปรบความขนแลว ใหหมาดบรเวณขางหลอดทดลอง จากนนน าเชอมาปาย (swab) บนจาน
อาหารเลยงเชอ Mueller Hhinton agar (MHA) 3 ระนาบใหทวบรเวณผวหนาจานอาหาร รอใหเชอทปายบนอาหารเลยง
เชอแหง แลวจงน ายาตานจลชพทตองการทดสอบความไวตอยาตานจลชพมาวางบนจานอาหารเลยงเชอ MHA ทท าการ
ปายเชอทดสอบแลว โดยใชเทคนคปราศจากเชอ จากนนน าไปบมท 37 องศาเซลเซยส เปนเวลา 24 ชวโมง แลวบนทก
ผลการทดสอบความไวตอยาตานจลชพโดยการวด zone of inhibition (mm) และแปลผลการทดสอบความไวตอยาตาน
จลชพโดยใชเกณฑมาตรฐานจาก Clinical and Laboratory Standards Institute (CLSI) antimicrobial
susceptibility testing standards 2014 ดงแสดงในตาราง 1
ตาราง 1 แสดงยาตานจลชพและการแปลผลการทดสอบความไวตอยาตานจลชพ
ยาตานจลชพ Resistance Intermediated Susceptibility
1. Amikacin (AK) 30 µg 14 15-16 17
2. Cefotaxime (CTX)30 µg 14 15-22 23
3. Ceftazidime (CAZ) 30 µg 14 15-17 18
4. Ceftriazone (CRO) 30 µg 13 14-20 21

21
5. Cefepime (FEP) 30 µg 14 15-17 18
6. Ciprofloxacin (CIP) 5 µg 15 16-20 21
7. Gentamicin (CN) 10 µg 12 13-14 15
8. Imipenem (IPM) 10 µg 13 14-15 16
9. Meropenem (MEM) 10 µg 13 14-15 16
10. TMX/SXT 1.25/23.75 µg 10 11-15 16
11. Tetracycline (TE) 30 µg 11 12-14 15
12. Cefoperazone/sulbactam
(SCF)105 µg
14 15-17 18
13. Piperacillin/tazobactam
(TZP)100/10 µg
17 18-20 21
14. Colistin (CT) 10 µg 9 10-11 12
15. Tigecycline (TGC) 15 µg 12 13-16 17
ทมา: Clinical and Laboratory Standards Institute (CLSI) antimicrobial susceptibility testing Standards,
2014 และ Piewngam, & Kiratisin, 2014
3. การตรวจหา virulence gene โดยใชวธ PCR
เตรยม cell lysate เพอใชเปน DNA template ส าหรบการท า PCR โดยเพาะเชอ A. baumannii ลงบนอาหารเลยง
เชอ TSA จากนนน าไปบมท 37 องศาเซลเซยส เปนเวลา 18-24 ชวโมง เขยโคโลนทเจรญมา 1 loop ใสลงใน หลอด
microcentrifuge ทม TE buffer 20 ไมโครลตรท าการตม เปนเวลา 15 นาท จากนนปนเหวยงท 10,000 rpm เปน
เวลา 3 นาท การท า PCR เพอตรวจหา virulence gene จ านวน 5 genes ไดแก bap, ompA, epsA, bauD-bauC
(Acinetobactin ) และ hly (hemolysin) โดยใช primer ตามตารางท 2 ซง PCR mastermix แตละหลอด มปรมาตร
รวม 20 µl ประกอบดวยน ากลนปราศจากเชอ (sterile 18 ohms distilled water) 13.1 ไมโครลตร 10x buffer 2
ไมโครลตร dNTP ความเขมขน 2.5 ไมโครโมลาร 2 ไมโครลตร MgCl2 ความเขมขน 25 ไมโครโมลาร 0.5 ไมโครลตร

22
forward primer ความเขมขน 20 ไมโครโมลาร 0.15 ไมโครลตร reverse primer ความเขมขน 20 ไมโครโมลาร 0.15
ไมโครลตร 0.5U DNA taqpolymerase (0.5 ยนต) 0.1 ไมโครลตร และ DNA template 2 ไมโครลตร เมอท า PCR
เสรจแลว น า PCR product ทได มาท าการตรวจหา DNA ดวยวธอะกาโรสเจลอเลคโตรโฟรสส เพอหาขนาดของ PCR
product โดยเทยบกบดเอนเอมาตรฐาน 100 bp (solis bioline) โดยใช 1% เจลอะกาโรสทมเอทเดยมโบรไมด ใชความ
ตางศกย 100 โวลต เปนเวลา 30 นาท จากนนตรวจดผลภายใตแสงยว
ตารางท 2 ขอมล primer ของ virulence gene
gene Primer sequence Product size (bp)
Tm (˚C)
references
bap TACTTCCAATCCAATGCTAGGGAGGGTACCAATGCAG TTATCCACTTCCAATGATCAGCAACCAAACCGCTAC
1,225 65 Goh, et al., 2013
ompA CGCTTCTGCTGGTGCTGAAT CGTGCAGTAGCGTTAGGGTA
531 50 Tayabali et al., 2012
epsA AGCAAGTGGTTATCCAATCG ACCAGACTCACCCATTACAT
451 50 Tayabali, et al.,2012
Acinetobactin (bauD-bauC)
AAGCTTATTCTGCGTTCG ACTGGCTTAATCCATGCG
1,200 52 Dorsey, et al., 2004
bfmS TTGCTCGAACTTCCAATTTATTATAC 1428 53 Liou et al., 2017 TTATGCAGGTGCTTTTTTATTGGTC Hemolysin (hly)
ATGAAAAAAATTATCTTTCTTGGTTTAGC TTAACCACATGTTGCTTGAGTAATTT
435 56 A1S:1321(NCBI)
blaPER-1 ATGAATGTCATTATAAAAGC AATTTGGGCTTAGGGCAAGAAA
1428 52 Brown et al., 2005
4. การศกษาคณสมบตการสรางไบโอฟลม
การศกษาคณสมบตการสรางไบโอฟลมของเชอ A. baumannii สายพนธตางๆจะถกน ามาศกษาการสรางไบโอฟลมโดยวธ
microtiter plate assay โดยขนตอนการท าเรมจากน าเชอแบคทเรยทเกบเปน stock มาเพาะเลยงลงบนอาหาร Trypticase soy
agar (TSA) แลวน าไปบมทอณหภม 37 องศาเซลเซยส เปนเวลา 24 ชวโมง ตอมากน าเชอแบคทเรยทผานการบมมาใสลงใน
normal saline 0.85% 8 มลลลตร โดยปรบความขนใหไดคาเทากบ McFarland 0.5 โดยเครอง densitometer ตอมาน า
อาหาร LB broth ทเตม 1% glucose ปรมาณ 50 มลลลตร มาแบงใสใน 96-well micrititer plate (flat bottom) หลมละ 100

23
ไมโครลตร ทกหลม จากนนจะมการแบงเปนหลม control จะเตม normal saline 0.85% หลมละ 100 ไมโครลตร สวนหลมท
ใชทดสอบ จะเตม normal saline 0.85% และ normal saline 0.85% ทมการลงเชอ ทความขนเทากบ McFarland
0.5 หลมละ 50 ไมโครลตร โดยปรมาตรรวมของทกหลมจะเทากบ 200 ไมโครลตร น าไปบมท 37 องศาเซลเซยส เปน
เวลา 24 ชวโมง จากนนจะเปนขนตอนการยอมสไบโอฟลม โดยน า 96-well micrititer plate ทบมไวมาคว าทง ตอมาจง
ท าการลางเซลลออกดวยน ากลน ปรมาตร 250 ไมโครลตร 2 ครง คว าทงแลวปลอยใหแหง (air-dried) 10 นาท แลว
fixed เซลลแบคทเรยดวยการเตม Absolute methanol ทกหลม ปรมาตร 250 ไมโครลตร บมไว 10 นาท คว าทงแลว
ปลอยใหแหง (air-dried) 10 นาท จากนนยอมสดวยการเตม 0.4% crystal violet ทกหลม ปรมาตร 250 ไมโครลตร บม
ไว 15 นาท คว าทงแลวลางออกดวยน ากลนทนท ปรมาตร 250 ไมโครลตร 2 ครง คว าทงแลวปลอยใหแหง (air-dried)
10 นาท จากนนเตม 33% acetic acid 250 ไมโครลตร บมไว 15 นาท แลวน าไปวดคาดดกลนแสงท OD595 โดยใช
เครอง Micro-plate Reader ปรมาณการสรางไบโอฟลมจะก าหนดตามคาการดดกลนแสงออกเปน 4 กลมดงน: (I) OD
≤ ODc = non biofilm Producer; (II) ODc < OD ≤ 2X ODc = weak biofilm Producer; (III) 2X ODc < OD ≤ 4X
ODc = moderate biofilm Producer; and (IV) 4X ODc < OD = strong biofilm producer โดยคา Optical
density cut-off value (ODc) ค านวณจากคาเฉลยของ OD ของ negative control + 3X standard deviation (SD)
of negative control. The (Hassan et al., 2011)
5. การศกษาคณสมบตการสรางฮโมไลซน
การศกษาคณสมบตการสรางฮโมไลซน ของเชอ A. baumannii สายพนธตางๆจะถกน ามาศกษาการสรางฮโมไลซน โดย
วธ Blood agar plate ซงดจากการยอยสลายของเมดเลอดแดงแกะ โดยขนตอนการท าเรมจากน า เชอแบคทเรยทเกบ
เปน stock มาเพาะเลยงลงบนอาหาร Trypticase soy agar (TSA) แลวน าไปบมทอณหภม 37 องศาเซลเซยส เปนเวลา
24 ชวโมง ใหไดโคโลนเดยวๆ (single colony) จ านวนมาก จากนนท าการเขยโคโลนเดยว มา Streak plate ลงบนอาหาร
Blood agar (Columbia agar ผสมกบ 5% v/v ของเลอดแกะ) น าไปบมทอณหภม 37 องศาเซลเซยส บมเปนเวลานาน
5 วน (ท าการเกบบนทกผลทกวน) สงเกตลกษณะการเกด clear zone โดยใช Bacillus cereus เปน positive control
แลวบนทกผล

24
6. การศกษาการเจรญของเชอ A. baumannii บนอาหารทม iron จ ากดและการสรางsiderophore
การศกษาการเจรญของเชอ A. baumannii บนอาหารทม iron จ ากดและการสราง siderophore ท าโดยน าเชอ A.
baumannii สายพนธตางๆมาเพาะเลยงบนอาหาร Chrome Azurol S agar (CAS agar) (Schwyn and Neilands,
1987) โดยน า 5 ไมโครลตรของเชอ A. baumannii ทปรบความขนโดยใชเครอง densitometer ใหมปรมาณเชอเทากบ
108 CFU/ml มาหยดบนจานอาหาร CAS โดยใชอาหาร LBA เปนจานควบคม แลวน าจานอาหารไปบมทอณหภม 37
องศาเซลเซยส อานผลใน 5 และ 10 วน โดยดการเจรญบนอาหาร CAS เทยบกบ LBA และการสราง siderophore
สงเกตจากเปลยนสอาหารจากฟาเปนเหลองหรอสม
7. การศกษาความสมพนธของการสรางไบโอฟลมในเชอ A. baumannii กบการดอยาและการม
virulence genes
การศกษาความสมพนธของการสรางไบโอฟลมในเชอ A. baumannii กบการดอยาและการสรางไบโอฟลมกบการม
virulence genes ในเชอจ านวน 225 สายพนธ (เฉพาะสายพนธทใหผลบวกกบยน blaOXa51) โดยเปรยบเทยบความ
แตกตางของกลมทสรางและไมสรางไบโอฟลมกบการดอยาและการมvirulence genes โดยใช Mann–Whitney U
test และ Fisher's exact test คา P-values < 0.05 ถอวามนยส าคญทางสถต

25
ผลการวจย (Result)
1. การตรวจหา virulence gene โดยใชวธ PCR
ผลการตรวจหา virulence genes 5 ยนแสดงดงรปท 2 ในตวอยาง A. baumannii clinical isolates จากโรงพยาบาล
สวรรคประชารกษ พบยน epsA จ านวน 17 ตวอยาง (25.76%) ompA จ านวน 49 ตวอยาง (74.24%) bap จ านวน
34 ตวอยาง (51.52%) hly (hemolysin) จ านวน 60 ตวอยาง (90.91%) และ bauD-bauC (Acinetobactin) จ านวน
2 ตวอยาง (3.03%) ผลการตรวจหา virulence genes ในตวอยาง A. baumannii clinical isolates จาก
โรงพยาบาลพจตร พบยน epsA จ านวน 19 ตวอยาง (34.55%) ompA จ านวน 45 ตวอยาง (81.82%) bap จ านวน
39 ตวอยาง (70.91%) hly (hemolysin) จ านวน 54 ตวอยาง (98.18%) และ bauD-bauC (Acinetobactin) จ านวน
1 ตวอยาง (1.82%) ผลการตรวจหา virulence genes ในตวอยาง A. baumannii clinical isolates จากโรงพยาบาล
เชยงรายประชานเคราะห พบยน epsA จ านวน 16 ตวอยาง (14.68%) ompA จ านวน 99 ตวอยาง (90.83%) bap
จ านวน 42 ตวอยาง (38.53%) hly (hemolysin) จ านวน 107 ตวอยาง (98.17%) และ bauD-bauC (Acinetobactin)
จ านวน 6 ตวอยาง (5.5%) (ตารางท 3) ผลการตรวจหา virulence ยนของเชอ Acinetobacter spp. จากแหลงน า
ธรรมชาต พบยน epsA จ านวน 13 ตวอยาง (68.42%) ompA จ านวน 14 ตวอยาง (73.68%) bap จ านวน 4 ตวอยาง
(21.05%) hly (hemolysin gene จ านวน 14 ตวอยาง (73.68%) และ ตรวจไมพบ bauD-bauC (Acinetobactin)
(ตารางท 3)
รปท 2 แสดงผล PCR product ของ virulence genes โดยแถว M คอ molecular weight marker แถว 1 คอ
Hemolysin gene (435 bp) แถว 2 คอ epsA (451 bp) แถว 3 คอ OmpA (531bp) แถว 4 คอ bap (1,225 bp)
แถว 5 คอ bauD-bauC (1,200 bp)

26
ตารางท 3 แสดงผลการตรวจหา virulence genes ในตวอยาง A. baumannii
Virulence gene/ แหลงทมาของเชอ
จ านวน(%) ของเชอ A. baumannii โรงพยาบาลสวรรคประชารกษ (N=66)
โรงพยาบาลพจตร (N=55)
โรงพยาบาลเชยงรายประชานเคราะห(N=109)
แหลงน าธรรมชาต (N=19)
ompA 49 (74.24) 45 (81.82) 99 (90.83) 14 (73.68) epsA (capsular polysaccharide)
17 (25.76) 19 (34.55) 16 (14.68) 13 (68.42)
bap (Biofilm-associated Protein)
34 (51.52) 39 (70.91) 42 (38.53) 4 (21.05)
bfmS 78.26 (54) 87.27 (48) 80.73 (88) 13 (68.4) hly (Hemolysin) 60 (90.91) 54 (98.18) 107 (98.17) 14 (73.68) bauD-bauC (Acinetobactin)
2 (3.03) 1 (1.8) 6 (5.5) 0 (0)
2. การศกษาคณสมบตการสรางไบโอฟลม
การศกษาคณสมบตการสรางไบโอฟลมเชอ A. baumannii สายพนธตางๆจะถกน ามาศกษาการสรางไบโอฟลมโดยวธ
microtiter plate assay ผลการสรางไบโอฟลมพบเชอ A. baumannii clinical isolates ทงหมด 230 ตวอยางพบเชอ
ทสรางไบโอฟลม (biofilm former) จ านวน 99 ตวอยาง คดเปน 43 % และเชอทไมสราง (Non- biofilm former)
จ านวน 131 ตวอยาง คดเปน 57% ดงแสดงในตารางท 4 โดยเชอจากโรงพยาบาลสวรรคสวรรคประชารกษพบ
ความสามารถในการสรางไบโอฟลมไดมากทสดคดเปน 78%
3. การศกษาคณสมบตการสรางฮโมไลซน
ผลการทดสอบการยอยสลายเมดเลอดแดงของเชอ A. baumannii ทไดมาจากตวอยางผปวยในโรงพยาบาล
พจตรจ านวน 55 ตวอยาง โรงพยาบาลสวรรคประชารกษจ านวน 69 ตวอยาง โรงพยาบาลเชยงรายประชานเคราะห
จ านวน 109 ตวอยาง ไมพบการยอยสลายเมดเลอดแดงใน ทกๆตวอยาง เมอเทยบกบ positive control คอ Bacillus
cereus แสดงดงภาพท 3

27
ตารางท 4 แสดงการสรางไบโอฟลมของเชอ A. baumannii ทแยกจาก โรงพยาบาลพจตร โรงพยาบาลเชยงราย
ประชานเคราะห และโรงพยาบาลสวรรคประชารกษ
ภาพท 3 แสดงการศกษาคณสมบตการสรางฮโมไลซน ของเชอ A. baumannii โดยวธ Blood agar plate
(A) เชอ A. baumannii สายพนธทน ามาทดสอบ (B) เชอ B. cereus เปน positive control
4. การศกษาการเจรญของเชอ A. baumannii บนอาหารทม iron จ ากดและ siderophore production
assay
การศกษาการสราง siderophore ของเชอ A. baumannii ท าบนอาหาร CAS (Schwyn and Neilands,1987)
และใช LBA ส าหรบเปน control โดยเลยงเชอ A. baumannii บนอาหาร LBA แลวน าไปบมท 37 องศาเซลเซยส เปน
เวลา 24 ชวโมง จากนนน าโคโลนทเจรญบนจานอาหารเลยงเชอมาปรบ ความขนในสารละลาย 0.85%NaCl ใหไดความ
ขนเทยบเทากบ 0.5 Mcfarland โดยใชเครองวดความขน (McFarland densitometer: SiaBiosan, Latvia) หยด (spot)
โรงพยาบาล Biofilm formation จ านวน(%)
โรงพยาบาลพจตร (จ านวน 55 ตวอยาง)
สรางไบโอฟลม (biofilm former) 11 (20) ไมสราง (Non- biofilm former) 44 (80)
โรงพยาบาลเชยงรายประชานเคราะห (จ านวน 109 ตวอยาง)
สรางไบโอฟลม (biofilm former) 36 (33.0) ไมสราง (Non- biofilm former) 73 (67)
โรงพยาบาลสวรรคประชารกษ(จ านวน 66 ตวอยาง)
สรางไบโอฟลม (biofilm former) 52 (78) ไมสราง (Non- biofilm former) 14 22)
A.
B.

28
A. baumannii ทท าการศกษาปรมาตร 10 ไมโครลตร บนอาหาร CAS และ LBA ดการเจรญบนจานอาหาร CAS ทก
5 และ 10 วนเชอทเจรญบนอาหาร CAS ดงแสดงในภาพท 4 โดยผลการเจรญของเชอ A. baumannii บนอาหารทม
iron จ ากดพบเชอเจรญบนอาหาร CAS จ านวน134 ตวอยาง คดเปน 58.3 % และเชอทไมเจรญบนอาหาร CAS จ านวน
96 ตวอยาง คดเปน 41.7 % (ตารางท 5) โดยเชอทงหมดไมพบการสราง siderophore
ภาพ 4 การทดสอบการเจรญบนอาหาร CAS ภายใน 5 วน (A) 10 วน (B) และการเจรญบนอาหาร LBA (C)
ตารางท 5 แสดงการการเจรญของเชอ A. baumannii บนอาหารทม iron จ ากดของเชอ A. baumannii ท
แยกจาก โรงพยาบาลพจตร โรงพยาบาลเชยงรายประชานเคราะห และโรงพยาบาลสวรรคประชารกษ
โรงพยาบาล Biofilm formation จ านวน(%)
โรงพยาบาลพจตร (จ านวน 55
ตวอยาง)
เจรญบนอาหาร CAS 34 (61.8)
ไมเจรญบนอาหาร CAS 21 (38.2)
โรงพยาบาลเชยงรายประชาน
เคราะห(จ านวน 109 ตวอยาง)
เจรญบนอาหาร CAS 63 (57.8)
ไมเจรญบนอาหาร CAS 46 (42.2)
โรงพยาบาลสวรรคประชารกษ
(จ านวน 66 ตวอยาง)
เจรญบนอาหาร CAS 37 (56.1)
ไมเจรญบนอาหาร CAS 29 (43.9)

29
5. การศกษาความสมพนธของการสรางไบโอฟลมในเชอ A. baumannii และรปแบบการดอยา
ผลการทดสอบความไวของยาปฏชวนะตอยาปฏชวนะ 13 ชนดพบวาเชอสวนใหญดอตอยา ciprofloxacin (84.4%) ดอ
ตอยา amikacin (54.2%) ดอตอยา cefotaxime (76.9%) ดอตอยา ceftaidime (82.2%) ดอตอยา ceftriaxone
(81.3%) ดอตอยา cefepime (67.6%) ดอตอยา gentamicin (68%) ดอตอยา imipenem (79.6% ) ดอตอยา
meropenem (78.7%) ดอตอยา trimethoprim / sulfamethoxazole (54.2%) ดอตอยา tetracycline (62.7%) ดอ
ตอยา cefoperazone / sulbactam (20.4%) และดอตอยา piperacillin / tazobactam (79.1%) พบเชอทเปน ของ
multidrug resistant A. baumannii (MDR-AB) เทากบ 86.2% (194/225) โดยจะจดเปน MDR-AB เมอดอยาปฏชวนะ
มากกวา 3 กลม ผลการสรางไบโอฟลมในเชอ MDR-AB ทงหมด 194 ไอโซเลทพบเชอ MDRAB 150 สายพนธ (77.3%)
ทสรางไบโอฟลม ขณะทกลมทไมใช MDRAB พบสรางไบโอฟลม 74.2% (23/31) (P = 0.654; Fisher's exact test) เมอ
ท าการวเคราะหคา median OD595 ระหวางรปแบบการดอยาสามชนด (ภาพ 5) ดงแสดงในรปท 5B คา median (IQR)
ของเชอกลมไมดอยา (non-MDRAB) กลมดอยา (MDR-AB) และกลมดอยารนแรง (XDR-AB) เทากบ 0.500 (0.157,
0.990), 0.461 (0.245, 0.794) และ 0.605 (0.444, 0.770) ตามล าดบ โดยไมมความแตกตางกนในกลม (P = 0.536;
Kruskal - Wallis test)
ภาพ 5 การสรางไบโอฟลม (OD595) ในเชอ Acinetobacter baumannii 225 ไอโซเลทแบงกลมตามรปแบบ
การดอยา (A) การสรางไบโอฟลมของเชอกลมไมดอยา (non-MDRAB) (สฟา) กลมดอยา (MDR-AB) (สเขยว) และกลม
ดอยารนแรง (XDR-AB) (สสม) (B) จดวงกลมแสดงคาเฉลยของการสรางไบโอฟลม (OD595) ของแตละไอโซเลท แถบ (ส
ด า) ตรงกลางแสดงคาเฉลยของการสรางไบโอฟลม OD595 ของแตละกลมทมรปแบบความตานทานยาปฏชวนะแตกตาง
กน เสนประจะตรงกบคาตด (ODC)

30
6. การศกษาความสมพนธของการสรางไบโอฟลมในเชอ A. baumannii และการม virulence genes
ความสมพนธของการสรางไบโอฟลมในเชอ A. baumannii 225 ไอโซเลทบน microtiter plate และ virulence genes
ทมรายงานวาเกยวของกบการสรางไบโอฟลมจ านวน 5 ยนไดแก epsA, bap, ompA, bfmS และ blaPER-1 โดยใช
Mann–Whitney U test พบวายน epsA, bfmS และ blaPER-1 ไมเกยวของกบการสรางไบโอฟลมในเชอ A. baumannii
(P> 0.05; ภาพท 6 C-E) เชอสายพนธทไมมยน bap หรอ ompA สรางไบโอฟลมไดดกวาเชอทมยน bap หรอ ompA
(P <0.05; ภาพท 6 A, B) อยางไรกตามคาความถของการมยน epsA, bfmS, blaPER-1, bap และ ompA ไมแตกตางกน
อยางมนยส าคญทางสถตกบการสรางหรอไมสรางไบโอฟลมโดยใช Fisher's exact test ทคา P-value 0.05 เทากบ
0.253, 0.281, 0.393, 0.117 และ 0.199 ตามล าดบ นอกจากนพบวาเมอวเคราะหการสรางไบโอฟลม (OD595) ของแต
ละไอโซเลทกบจ านวนการม virulence gene พบวาการมจ านวน virulence gene เพมขนเชอมแนวโนมสรางไบโอฟลม
ลดลงแตไมแตกตางกนอยางมนยส าคญทางสถต (P > 0.05; Kruskal–Wallis test) ( ภาพ 6F).
ภาพ 6 ความสมพนธของการสรางไบโอฟลมในเชอ A. baumannii และการม virulence genes
(A–E) แสดงการสรางไบโอฟลม (OD595) บน microtiter plate ของเชอกลมทมและไมม virulence genes แตละชนด
เครองหมาย (*) แสดงคานยส าคญทางสถตโดยใช Mann–Whitney U-test (F) การสรางไบโอฟลม (OD595) กบจ านวน
การม virulence gene โดย P0 เชอไมพบ virulence gene และ P1–P5 แสดงการม virulence gene ตงแต 1-5
ยนตามล าดบ จดวงกลมแสดงคาเฉลยของการสรางไบโอฟลม (OD595) ของแตละไอโซเลท individual isolates แถบ
(สด า) ตรงกลางแสดงคาเฉลยของการสรางไบโอฟลม OD595 ของแตละกลม

31
ขอวจารณ (Discussion)
การตรวจหา virulence gene ในเชอ A. baumannii
เชอ A. baumannii มปจจยทกอใหเกดความรนแรงของโรคหลายปจจย ไดแก การสรางไบโอฟลม การสรางแคปซล
การสรางไซเดอรโรฟอร และการสรางฮโมไลซน เปนตน จากการศกษาครงนตรวจพบยนท เกยวของกบการสรางไบโอ
ฟลมหลากหลายยนในเชอทท าการศกษา ไดแก bap, ompA, epsA, bfmS และ blaPER-1 เปนตน โดยตรวจพบยน
ompA มากทสด การศกษาในครงนสอดคลองกบหลายงานวจยทตรวจพบยน bap, ompA, bfmS ช ง เปนยนมผล
ตอการสรางไบโอฟลมเชนการศกษาของ Liu และคณะในป ค.ศ.2016 ท าการศกษาการสรางไบโอฟลมในเชอ A.
baumannii ดอยา พบยน ompA รอยละ 100 จากการศกษาของ Smani และคณะ ในป ค.ศ.2014 พบวา ยน ompA
นอกจากเปนยนทควบคมการสรางไบโอฟลมแลว ยนนยงสงผลใหเชอปรบตวกลายเปนเชอด อยาไดอกดวย (Smani, et
al., 2014) ปจจยทกอความรนแรงของโรคอกปจจยหนง ไดแก การสรางอะซนโตแบคตน (acinetobactin) ในเชอ A.
baumannii โดย acinetobactin เปนสารประเภทโปรตนทมความจ าเพาะ (high affinity) ในการจบกบธาตเหลกใน
รปของ Fe3+ ในเซลลเจาบาน (host) โดยสามารถจบกบโมเลกลทมธาตเหลกเปนองคประกอบ ไดแก transferrin,
lactoferrin, ferritin และโมเลกลทม heme เปนองคประกอบ เปนตน ธาตเหลกมความส าคญตอเซลลเจาบาน
เนองจากเปน cofactor ในกระบวนการเมทาบอลซม หากปรมาณธาตเหลกในเซลลเจาบานมไมเพยงพอ อาจสงผลใหม
ความผดปกตในการท างานของระบบเมทาบอลซมตางๆ ได องคประกอบของ acinetobactin ประกอบดวยสาร 3 ชนด
คอ 2,3-dihydroxybenzoic acid (DHBA), L-threonine และ N-hydroxyhistamine (Hasan, et al., 2015)
ยนทควบคมการสราง acinetobactin แบงไดเปน 3 กลม ไดแก กลมยน A. baumannii acinetobactin biosynthesis
(Bas) ไดแก basABCDEFGHIJ กลมยน A. baumannii acinetobactin utilization (Bau) ไดแก bauABCDEF และ
กลมยน A. baumannii acinetobactin release (Bar) ไดแก barAB (Mihara et al., 2004) ในการศกษาครงนท าการ
ตรวจหายน BauD-bauC ซงอยในกลมยน A. baumannii acinetobactin utilization (Bau) พบความชกรอยละ 3.47
ในเชอ clinical isolates สวนการศกษาหายนทควบคมการสรางฮโมไลซน (hly) (Antunes, et al., 2011) พบรอยละ
ความชกเทากบ 100

32
การสรางฮโมไลซน
ฮโมไลซน (Hemolysin) เปนสารพษ (toxin) สามารถท าใหเกดความรนแรงของโรคไดมากขน โดยการท าใหเกด
ร (pore-forming) บรเวณเยอหมเซลลของเซลลหลายชนด ไดแก เซลลเมดเลอดแดง (erythrocyte) เซลล monocytes
เซลล neutrophil และ เซลล macrophage โดยยนทเกยวของกบการสรางฮโมไลซน คอยน hly ซงท าหนาทใหมการ
สราง exotoxin ออกฤทธท าลายเยอหมเซลลของเมดเลอดแดง โดยฮโมไลซนจะท าใหเยอหมเซลลเกดเปนร และเปน
สาเหตทท าใหเซลลเมดเลอดแดงแตก (Zhang et al, 2005) จากผลการตรวจหายน hly ดวยเทคนค PCR และตรวจหา
การสรางฮโมไลซนโดยวธ Blood agar plate พบวา การมยน hly ไมมผลตอการท าใหเมดเลอดแดงของแกะแตก โดย
เชอไมพบความสามารถในการยอยสลายเมดเลอดแดงในทกๆตวอยาง เมอน าเทยบกบ positive control (B. cereus)
แตมการตรวจพบยน โดยคดเปน 90-98% ของเชอทแยกไดในโรงพยาบาล สอดคลองกบการศกษาทางระบาดวทยาใน
การสรางฮโมไลซนโดย Bitrian และคณะพบวามการตรวจพบการสรางฮโมไลซนจ านวน 1 สายพนธ คอเชอ A.
haemolyticus จากทงหมด 9 สายพนธ ของ Acinetobacter genospecies clinical isolates และตรวจไมพบการ
สรางฮโมไลซนในเชอ A. baumannii ทน ามาทดสอบทงหมด (Bitrian et al, 2012) รวมทงการศกษากอนหนานมการ
รายงานของ Tayabali และคณะมการทดสอบ Haemolytic Activity ในเชอกลม Acinetobacter spp. พบวาม 2 ใน 7
สายพนธทมการยอยสลายเมดเลอดแดงแกะซงไดแกเชอ A. haemolyticus และ A. venetianus (Tayabali et al,
2012) จากผลการศกษาในครงนแสดงใหเหนวาการมยน hly ไมมความสมพนธกบการสรางฮโมไลซนทมผลท าใหเกดการ
แตกของเมดเลอดแดงแกะในเชอ A. baumannii ทน ามาทดสอบ ซงอาจเปนไปไดวาการสรางฮโมไลซนของเชอ A.
baumannii ไมมผลตอการยอยสลายเมดเลอดแดงของแกะ แตถาเปนเมดเลอดแดงของสงมชวตอนอาจจะสงผลใหมการ
แตกของเมดเลอดแดงได เนองจากในกระบวนการออกฤทธของฮโมไลซนขนตอนทฮโมไลซนจะไปจบกบเมดเลอดแดง
อาจขนอยกบความจ าเพาะบนพนผวของเมดเลอดแดงได และเมอน า sequence ของยน hly (A1S_1321) ของเชอ A.
baumannii ทใชในการเพมจ านวนของยนในครงนไปเทยบกบ sequence ของเชอ Escherichia coli str. K-12 substr.
MG1655 โดยท าการ BLAST ใชฐานขอมลใน NCBI พบวามยน hlyE อย โดยม % identity เทากบ 100% จงสามารถ
สรปไดวาเชอ A. baumannii มยนทก าหนดการสรางฮโมไลซน แตจะท าใหเมดเลอดแดงแตกในบางสงมชวตเทานน จง
ควรจะมการใชเมดเลอดแดงของสงมชวตอนๆน าทดสอบตอไป

33
การเจรญของเชอ A. baumannii บนอาหารทม iron จ ากด
การเจรญของเชอ A. baumannii บนอาหารทม iron จ ากดrพบวาเชอ A. baumannii บางสวนไมสามารถ
เจรญบนอาหารทม iron จ ากดไดและเชอทเจรญบนอาหารทม iron จ ากดไมพบการสราง siderophore หรอ
Acinetobactin โดย Acinetobactin เปน siderophore ทผลตในเชอ A. baumannii มคณสมบตเปน catechol-
hydroxamate siderophore จะสรางออกมาในสภาวะทม iron จ ากด A. baumannii ผลต acinetobactin และ
ferric-siderophore receptor ทผนงเซลล Acinetobactin จะจบกบ ferric เปน acinetobactin-ferric complex และ
ขนสงไปในเซลลผาน ferric-siderophore receptor ผานระบบ siderophore efflux system เพอใชน า iron ไปใชใน
เซลล (Kamjam & Pathom-aree., 2014) การทไมพบการสราง siderophore บนอาหาร CAS อาจเปนผลมาจาก
siderophore ในเชอ A. baumannii มหลายชนดและถกควบคมการสรางโดยกลมของยนหลายกลมเชนไดแก กลมยน
A. baumannii acinetobactin biosynthesis (Bas) ไดแก basABCDEFGHIJ กลมยน A. baumannii acinetobactin
utilization (Bau) ไดแก bauABCDEF และกลมยน A. baumannii acinetobactin release (Bar) (Mihara et al.,
2004; Shapiro & Wencewicz, 2016) นอกจากนการผลต siderophore ของเชอแตละชนดยงขนกบองคประกอบใน
อาหารโดยกรด succinic acid จะกระตนการสราง siderophore ไดดกวา glucose (Sayyed et al., 2005) และความ
เขมขนของ HDTMA ทสงเกนไปในอาหาร CAS อาจเปนพษตอการผลต siderophore ในแบคทเรยแกรมลบ (Schwyn
and Neilands, 1987)
การสรางไบโอฟลมในเชอ A. baumannii และรปแบบการดอยา
การศกษากอนหนานโดย Sanchez และคณะ (2013) พบความสมพนธของรปแบบการดอยาแบบ MDR-AB กบการ
สรางไบโอฟลมในเชอ A. baumannii (Sanchez et al., 2013) แตการศกษาครงนไมพบความสมพนธของการสรางไบ
โอฟลมในเชอ A. baumannii และรปแบบการดอยา แตความสามารถของเชอในการสรางไบโอฟลมอาจเกยวของกบ
การดอยาในเชอ A. baumannii ตวอยางเชน Naparstek และคณะ (2014) ศกษาความสามารถของเชอ Klebsiella
pneumoniae ในการสรางไบโอฟลมและไดขอสรปวาสายพนธททนตอ gentamicine ระดบสง จะสรางสรางไบโอฟลม
ไดดกวากลมไมดอยา (Naparstek et al., 2014) และในป 2016 Duarte และคณะพบวาเชอ A. baumannii ทดอตอ
ยา gentamicin และ tobramycin สามารถสรางไบโอฟลมไดดกวาสายพนธทไมดอยา (Duarte et al., 2016) โดยใน
การศกษาครงนผวจยพบวาความสามารถของเชอในการสรางไบโอฟลมของเชอสมพนธกบการดอตอยา gentamicin

34
การสรางไบโอฟลมในเชอ A. baumannii และการม virulence genes
การศกษาครงนพบวาเชอ A. baumannii ทมยนทท าใหเกดความรนแรงไมสงเสรมความสามารถในการสรางไบโอฟลม
ขณะทการมยน bap หรอ ompA กลบใหผลการสรางไบโอฟลมลดลง แมวาหลายรายงานไดแสดงใหเหนวายนท
เกยวของกบไบโอฟลม ไดแก bap, ompA, epsA, bfmS และ blaPER-1 มสวนท าใหมการสรางไบโอฟลมของ A.
baumannii บางสายพนธ (Gallant et al., 2005; Loehfelm et al., 2008; Russo et al., 2010; Tayabali et al.,
2012; Liou et al., 2014) รายงานเหลานศกษาความเกยวของของการสรางไบโอฟลมของ A. baumannii สายพนธท
จ าเพาะเพยงไมกสายพนธและศกษาบนพนผวทแตกตางกน อยางไรกตามการศกษาครงนพบความสมพนธของการมยน
ompA กบเชอทมรปแบบการดอยาแบบ XDR-AB

35
สรปและขอเสนอแนะ (Conclusion and Recommendation)
สรปผลงานวจย
ในการศกษานเราสามารถตรวจพบ virulence gene ในเชอ A. baumannii โดยพบยน hly (90-98%) ทความ
ชกสงสดตามดวย ompA (74-90%) และตรวจพบยน BauD-bauC ซงอยในกลมยน A. baumannii acinetobactin
utilization (Bau) ต าสด (2.8-5.5 %) โดยการมยน ompA มความสมพนธกบเชอทมรปแบบการดอยาแบบ XDR-AB
และไมพบความสมพนธของการสรางไบโอฟลมในเชอ A. baumannii และรปแบบการดอยาและการม virulence gene
แตพบความสมพนธของการสรางสรางไบโอฟลมในเชอ A. baumannii กบการดอยาเปนรายชนดเชนการดอยา
gentamicin
ขอเสนอแนะ
การศกษาปจจยทเกยวของกบความรนแรงของโรคควรมการศกษาตอไปเกยวกบการแสดงออกของ virulence gene
กบการสรางไบโอฟลมและศกษากลไกการท างานและแสดงออกของยนทเกยวของกบการสราง acinetobactin ทง 3
กลมตอไปนอกจากนควรมการศกษาเพมเตมในทกระบบของกลมยนทเกยวของกบการสราง acinetobactin ไดแกกลม
ยน A. baumannii acinetobactin biosynthesis (Bas); basABCDEFGHIJ, กลมยน A. baumannii acinetobactin
utilization (Bau); bauABCDEF และกลมยน A. baumannii acinetobactin release (Bar) ตอไป

36
เอกสารอางอง (reference)
1. Antunes, L. C. S., Imperi, F., Carattoli, A., & Visca, P. (2011). Deciphering the Multifactorial Nature
of Acinetobacter baumannii Pathogenicity. PLoS ONE, 6(8), 1-10.
2. Bitrian, M., Solari, C.M., González, R.H. & Nudel, C.B. (2012). Identification of virulence markers
in clinically relevant strains of Acinetobacter genospecies. International Microbiology, 15(2), 79-
88.
3. Brown, S., Young, H.K., & Amyes, S.G. (2005.) Characterization of OXA-51, a novel class D
carbapenemase found in genetically unrelated clinical strains of Acinetobacter baumannii from
Argentina. Clinical Microbiology and Infection, 11(1), 15-23.
4. Chaiwarith, R., Mahatthanaphak, S., Boonchoo, M., Supparatpinyo, K. & Sirisanthana, T. (2005).
Pandrug-Resistant Acinetobacter baumannii at Maharaj Nakorn Chiang Mai Hospital. Journal of
Infectious Diseases and Antimicrobial Agents, 22(1), 347-352.
5. Choi, C.H., Lee, E.Y., Lee, Y.C., Park, T.I., Kim, H.J., Hyun, S.H., Kim, S.A., Lee, S.K. & Lee, J.C.
(2005). Outer membrane protein 38 of Acinetobacter baumannii localizes to the mitochondria
and induces apoptosis of epithelial cells. Cellular Microbiology, 7(8), 1127-1138.
6. Choi, C.H., Lee, J.S., Lee, Y.C., Park, T.I. & Lee, J.C. (2008). Acinetobacter baumannii invades
epithelial cells and outer membrane protein A mediates interactions with epithelial cells. BMC
Microbiology, 10(8), e216.
7. Duarte, A., Ferreira, S., Almeida, S., & Domingues, F.C. (2016) Clinical isolates of Acinetobacter
baumannii from a Portuguese hospital: PFGE characterization, antibiotic susceptibility and
biofilm-forming ability. Comparative Immunology, Microbiology & Infectious Diseases, 45, 29-
33.
8. Gaddy, J.A., Arivett, B.A., McConnell, M.J., Lopez-Rojas, R., Pachon, J. & Actis, L.A. (2012). Role

37
of acinetobactin-mediated iron acquisition functions in the interaction of Acinetobacter
baumannii ATCC 19606 with human lung epithelial cells, Galleria mellonella caterpillars and
mice. Infection and Immunity, 80(3), 1015-1024.
9. Gallant, C.V., Daniels, C., Leung, J.M., Ghosh, A.S., Young, K.D., Kotra, L.P., & Burrows, L.L.
(2005) Common beta-lactamases inhibit bacterial biofilm formation. Molecular Microbiology,
58(4), 1012–1024.
10. Goh, H.M.S., Beatson, S.A., Totsika, M., Moriel, D.G., Phan, M.D., Szubert, J., Runnegar, N., Sidjabat,
H.E., Paterson, D.L., Nimmo, G.R., Lipman, J., & Schembri, M.A. (2013). Molecular Analysis of the
Acinetobacter baumannii Biofilm-Associated Protein. Applied and Environmental Microbiology,
79(21), 6535-6543.
11. Hassan, A., Usman, J., Kaleem, F., Omair, M., Khalid, A., & Iqbal, M. (2011). Evaluation of different
detection methods of biofilm formation in the clinical isolates. Brazilian Journal of Infectious
Diseases, 15(4), 305-311.
12. Hasan, T., Choi, C. H., & Oh, M. H. (2015). Genes Involved in the Biosynthesis and Transport of
Acinetobactin in Acinetobacter baumannii. Genomics & Informatics, 13(1), 2-6.
13. Inchai, J., Liwsrisakun, C., Theerakittikul, T., Chaiwarith, R., Khositsakulchai, W. & Pothirat, C.
(2015). Risk factor of multidrug-resistant, extensively drug-resistant and pandrug-resistant
Acinetobacter baumannii ventilator-associated pneumonia in a Medical Intensive Care Unit of
University Hospital in Thailand. Journal of Infection and Chemotherapy, 21(8), 570-574.
14. Kamjam, M. & Pathom-aree, W. (2014). Siderophores from Microorganisms. Srinakharinwirot
Science Journal, 30(1), 230-247.
15. Liou, M.L., Soo, P.C., Ling, S.R., Kuo, H.Y., Tang, C.Y., & Chang, K.C. (2014). The sensor kinase
BfmS mediates virulence in Acinetobacter baumannii. Journal of Microbiology, Immunology
and Infection, 47, 275-281.

38
16. Liu, L., Cui, Y., Zheng, B., Jiang, S., Yu, W., Shen, P., Ji J., Li L., Qin N., & Xiao, Y. (2016). Analysis
of tigecycline resistance development in clinical Acinetobacter baumannii isolates through a
combined genomic and transcriptomic approach. Scientific Reports, 31(6), 1-12.
17. Loehfelm, T.W., Luke, N.R., & Campagnari, A.A. (2008). Identification and characterization of an
Acinetobacter baumannii biofilm-associated protein. Journal of Bacteriology, 190, 1036-44.
18. McConnell, M.J., Actis, L. & Pachón, J. (2012). Acinetobacter baumannii: Human infections,
factors contributing to pathogenesis and animal models. FEMS Microbiology Reviews, 37(2),
130-155.
19. Mihara, K., Tanabe, T., Yamakawa, Y., Funahashi, T., Nakao, H., Narimatsu, S., & Yamamoto, S.
(2004). Identification and transcriptional organization of a gene cluster involved in
biosynthesis and transport of acinetobactin, a siderophore produced by Acinetobacter
baumannii ATCC 19606T. Microbiology, 150(8), 2587-2597.
20. Naparstek, L., Carmeli, Y., Navon-Venezia, S., & Banin, E. (2014). Biofilm formation and
susceptibility to gentamicin and colistin of extremely drug-resistant KPC-producing Klebsiella
pneumoniae. Journal of Antimicrobial Chemotherapy, 69, 1027–1034.
21. Russo, T.A., Luke, N.R., Beanan, J.M., Olson, R., Sauberan, S.L., MacDonald, U., Schultz, L.W.,
Umland, T.C., & Campagnari, A.A. (2010). The K1 capsular polysaccharide of Acinetobacter
baumannii strain 307-0294 is a major virulence factor. Infection and Immunity, 78, 3993-4000.
22. Sanchez, C.J. Jr., Mende, K., Beckius, M.L., Akers, K.S., Romano, D.R., Wenke, J.C., & Murray,
C.K. (2013). Biofilm formation by clinical isolates and the implications in chronic infections.
BMC Infectious Diseases, 13(47), 1-12.
23. Sayyed, R.Z., Badgujar, M.D., Sonawane H.M., Mhaske, M.M., & Chincholkar, S.B. (2005).
Production of microbial iron chelators siderophores by fluorescent pseudomonads. Indian

39
Journal of Biotechnology, 4, 484-190.
24. Schwyn, B., & Neilands, J.B. (1987). Universal chemical assay for the detection and
determination of siderophores. Analytical Biochemistry, 160(1), 47-56.
25. Shapiro, J. A. & Wencewicz, T.A. (2016). Acinetobactin Isomerization Enables Adaptive Iron
acquisition in Acinetobacter baumannii through pH-Triggered Siderophore Swapping. ACS
Infectious Diseases, 2(2), 157-168.
26. Smani, Y., Fàbrega, A., Roca, I., Sánchez-Encinales, V., Vila, J. & Pachón, J. (2014). Role of OmpA
in the Multidrug Resistance Phenotype of Acinetobacter baumannii. Antimicrobial Agents and
Chemotherapy, 58(3), 1806–1808.
27. Tayabali, A.F., Nguyen, K.C., Shwed, P.S., Crosthwait, J., Coleman, G. & Seligy, V.L. (2012).
Comparison of the virulence potential of Acinetobacter strains from clinical and environmental
sources. PLoS One, 7(5), e37024.
28. Zhang, X.H. & Austin, B. (2005). Haemolysins in Vibrio species. Journal of Applied Microbiology,
98(5), 1011-1019.

40
บทท 2
การศกษาคณสมบตทางอณชววทยาของการถายทอดยน
ดอยาในเชอ Acinetobacter baumannii
สวนหนงของการศกษาไดรบการตพมพในวารสาร
1. Leungtongkam U, Thummeepak R, Wongprachan S, Thongsuk P, Kitti T, Ketwong K, Runcharoen C, Chantratita N, Sitthisak S. Dissemination of blaOXA-23, blaOXA-24, blaOXA-58, and blaNDM-1 Genes of Acinetobacter baumannii Isolates from Four Tertiary Hospitals in Thailand. Microbial Drug Resistance. 2017 Jun 8. doi: 10.1089/mdr.2016.0248.
2. Prabhu Raj Joshi, Mahesh Acharya, Trishna Kakshapati, Udomluk Leungtongkam, Rapee Thummeepak and Sutthirat Sitthisak. Co-existence of bla OXA-23 and bla NDM-1 genes of Acinetobacter baumannii isolated from Nepal: antimicrobial resistance and clinical significance. Antimicrobial Resistance & Infection Control. 2017; 6:21
Manuscript preparation Udomluk Leungtongkam, Rapee Thummeepak, Kannipa Tasanapak and Sutthirat Sitthisak. Acquisition and transfer of antibiotic resistance genes and integrons of Acinetobacter baumannii by conjugation (This manuscript will submit in Scientific report)

41
บทน า (Introduction)
A. bauamanii เปนเชอแบคทเรยแกรมลบ รปทอนกลม พบไดทวไปในธรรมชาตรวมทงสงแวดลอมภายใน
โรงพยาบาล มบทบาทส าคญในการกอใหเกดโรคตดเชอภายในโรงพยาบาล โรคตดเชอในระบบทางเดนหายใจ โรคตด
เชอในกระแสเลอด และโรคตดเชอบรเวณบาดแผล โดยเฉพาะในผปวยทพกรกษาตวอยภายในหอผปวยหนกเปน
เวลานาน (Towner KJ ,2009 ) จากการรายงานของศนยเฝาระวงเชอดอยาแหงชาต กระทรวงสาธารณสข ประเทศไทย
ในป ค.ศ. 2014 พบวาแยกเชอ A. baumannii ไดเปนอนดบหนงจากตวอยางเสมหะทงหมดทแยกไดจากสงสงตรวจ
(NARST, 2017) การศกษาล าดบการพฒนาการดอยาของเชอ A. baumannii พบวาในชวง ค.ศ.1970 พบการตดเชอท
มสาเหตมาจาก Acinetobacter spp. จากสงสงตรวจเปนจ านวนมาก แตยงท าการรกษาไดงาย โดยใชยาตานจลชพใน
กลม β-lactam เชน ampicillin, second-generation cephalosporins, minocycline, colistin หรอ gentamicin
(Livermore and Woodford, 2006) ตอมามการใชยาตานจลชพมากเกนความจ าเปน สงผลใหในระหวางป ค.ศ. 1980
ถง ค.ศ. 1990 พบเชอดอตอยาตานจลชพสามขนานขนไปทเรยกวา multidrug-resistant A. baumannii (MDR-AB)
และดอตอยาในกลม carbapenem ไดแก imipenem และ meropenem ทเรยกวา carbapenem-resistant A.
baumannii (CR-AB) ซงเปนยาทมฤทธยบยงกวาง นยมใชรกษาการตดเชอจากแบคทเรยแกรมลบ รวมทง A.
baumannii (Poirel and Nordmann, 2006) โดยมรายงานในประเทศฮองกง พบ carbapenem-resistant A.
baumannii (CR-AB) เพมสงขนจากรอยละ 2.6 ในป ค.ศ. 1997 เปนรอยละ 29.4 ในป ค.ศ. 2008 (Wu, 2011) ใน
ประเทศไทย มรายงานการศกษาพบวา ในระหวางป ค.ศ.1996 ถง ค.ศ.1997 ภายในโรงพยาบาล ศรราชพบความชกของ
MDR-AB รอยละ 57.6 (Aswapokee et al.,1998) สวนภายในโรงพยาบาลมหาราชนครเชยงใหม ในป ค.ศ.2003 พบ
ความชกของ MDR-AB รอยละ 23 และเชอดอตอยาในกลม carbapenem มากทสด (Chaiwarith et al., 2005) ในขณะ
ทโรงพยาบาลพระมงกฎเกลา พบ CR-AB ถงรอยละ 84.2 โดยทกไอโซเลตทแยกไดไวตอยา colistin และ tigecycline
(Aimsaad et al., 2009) สวนในผปวยทใชเครองชวยหายใจภายในหอผปวยอายรกรรม โรงพยาบาลสงขลานครนทร ใน
ป ค.ศ. 2012 พบวาผปวยมการตดเชอ A. baumannii รอยละ 55 โดยทกไอโซเลตทแยกไดดอตอยา imipenem ซง
จดเปนยาในกลม carbapenem ปจจปนพบรปแบบการดอยาของเชอ A. baumannii เพมขนอกรปแบบหนง คอเชอ A.
baumannii ทดอยาทกกลม ยกเวนยาในกลม tigecyclin และ colistin เรยกเชอ A. baumannii ทดอยากลมนวา

42
extensively-drug resistance Acinetobacter baumannii หรอ XDR-AB (Inchai, et al., 2015) โดยการดอยา
รปแบบนเรมพบไดมากขนพบการระบาดในหลายประเทศทวโลก ไดแก ประเทศเลบานอน ไทย อหราน จน ไตหวน
เนปาลเปนตน (Inchai, et al., 2015; Moghnieh, et al., 2016; Farshadzadeh, et al., 2015; Joshi, et al., 2017)
ซงเหนไดวาพบอตราการตดเชอ A. baumannii ทดอตอยา เพมสงขนในระยะเวลาอนรวดเรว จดเปนปญหาทาง
สาธารณสขส าคญทพบทวโลก รวมทงประเทศไทยดวย การดอยาเกดจากกลไกการดอยาหลายกลไกดวยกน ไดแก การ
สรางเอนไซมท าลายยา การดดแปลงโครงสรางของ outer membrane หรอ penicillin binding proteins ซงเปน
เปาหมายในการจบของยา และการขบยาออกนอกเซลลโดย effluk pumps (Peleg et al., 2008) โดยการดอยากลม
ตางๆและยนดอยาทพบใน A. baumannii แสดงดงตารางท 6
ตาราง 6 แสดงยนดอยาทพบใน A. baumannii
ยน ต าแหนงทพบ ดอยา แหลงทมา
OXA-type carbapenemase - blaOXA-23 - blaOXA-24 - blaOXA-51 - blaOXA-58
พลาสมดหรอโครโมโซม พลาสมดหรอโครโมโซม
โครโมโซม พลาสมดหรอโครโมโซม
คารบาพเนม คารบาพเนม คารบาพเนม คารบาพเนม
Peleg et al.,2008 Peleg et al.,2008 Peleg et al.,2008 Peleg et al.,2008
blaAmpC
โครโมโซม
เซฟาโลสฟอรน ทกชนด
Poirel and Nordmann, 2006
blaIMP พลาสมด คารบาพเนม Poirel et al.,2011 blaVIM พลาสมด คารบาพเนม Poirel et al.,2011 blaNDM โครโมโซม คารบาพเนม Poirel et al.,2011 blaTEM พลาสมดหรอโครโมโซม เซฟาโลสฟอรน
รนท 3 ทกชนด Manchanda et
al.,2010 blaSHV
พลาสมดหรอโครโมโซม
เซฟาโลสฟอรน รนท 3 ทกชนด
Manchanda et al.,2010

43
การดอยาในกลม β-lactams และยนดอยาทพบใน A. baumannii มกลไกส าคญและพบไดบอย คอ
การสรางเอนไซม β-lactamase ออกมาท าลายยา มการจดกลมเอนไซมตามคณสมบตทางอณชววทยาหรอโครงสราง
ระดบโมเลกล แบงออกเปน 4 กลม ตามการจดจ าแนกของ Ambler (Ambler classification) ไดแก Class A beta-
lactamases ซ ง ม หม serine อย บ ร เ วณ active site ถ กย บย ง ด ว ย clavalanic acid พบมาก ในแบคท เ ร ย
Enterobacteriaceae ต อ ม า พ บ KPC-like enzyme แ ล ะ GES-like enzyme ใ น carbapenem-resistant
A.baumannii (CR-AB) ในประเทศเปอรโตรโกและปารสตามล าดบ (Robledo et al., 2010; Bonnin et al., 2012),
Class B metallo-beta-lactamases (MBLs) ใน A. baumannii พบยน blaIMP, blaVIM และ blaSIM ซงพบมากใน
แถบภมภาคเอเชยแปซฟก ไดแก ประเทศญปน ฮองกง เกาหลใต และออสเตรเลย (Mendes et al., 2009), Class C
AmpC-beta-lactamases มหม serine อยบรเวณ active site ถกยบยงดวย boronic acid พบยน blaAmpC เพมการ
แสดงออกของ β-lactamase เปนผลใหดอตอยา ceftazidime แตไมดอยาในกลม carbapenems (Poirel and
Nordmann; 2006) และ Class D serine-carbapenemase หรอ OXA type carbapenemases พบมากทสดใน
carbapenem-resistant A.baumannii (CR-AB) (Zarrilli et al., 2009) OXA type carbapenemases ถกพบครง
แรกในปค .ศ .1985 ในเชอ A. baumannii ทแยกไดจากโรงพยาบาลในประเทศสกอตแลนด ใหชอวา ARI-1
(Acinetobacter Resistant to Imipenem) หรอทเรยกวา OXA-23 (Paton et al., 1993) ในปจจบนพบ OXA type
carbapenemases หลายชนด ไดแก OXA-23-like , OXA-40-like , OXA-51-like , OXA-58-like และ OXA-143-
like โดยพบ OXA-51-like มากทสด การแพรกระจายของ OXA type carbapenemases พบไดทวโลก เชน OXA-
23-like และ OXA-58-like พบในประเทศบราซล เวเนซเอลา โคลมเบย โบลเวย และชล OXA-40-like พบในประเทศ
บราซลและพบมากในบางประเทศแถบยโรป สวน OXA-143-like พบในประเทศบราซล (Higgins et al., 2009;
Mendes et al., 2009; Zarrilli et al., 2013) ส าหรบในประเทศไทย มรายงานพบยนดอยาในกลม OXA type
carbapenemases ไดแก blaOXA-23 , blaOXA-24/40 และ blaOXA-58 (Mendes et al., 2009) โดยพบ blaOXA-23 ใน
โรงพยาบาลศรราช จงหวดกรงเทพมหานคร, โรงพยาบาลพทธชนราช จงหวดพษณโลก, โรงพยาบาลหวหน จงหวด
ประจวบครขนธ และโรงพยาบาลสรรพสทธประสงคจงหวดอบลราชธาน (Niumsup et al., 2009; Thapa et al.,
2010; Jantama et al. 2013; Santimaleeworagun et al., 2014) สวน blaOXA40-like พบในโรงพยาบาลหวหน

44
จงหวดประจวบครขนธ ซงม sequence ใกลเคยง blaOXA40 ถง 99% (GenBank accession no. KJ748571) พบเปน
ครงแรกในประเทศไทย และ blaOXA-58 ในโรงพยาบาลสรรพสทธประสงค จงหวดอบลราชธาน
การแพรกระจายของยนดอยา มกเกดจากการถายทอดยนดอยาในแนวราบ (horizontal gene transfer) โดย
เปนการถายทอดยนระหวางแบคทเรยชนดหนงไปสแบคทเรยอกชนดหนง โดยอาศยกระบวนการทส าคญ คอ ทรานส
ฟอรเมชน, ทรานสดกชน และคอนจเกชน (Huddleston, 2014) มรายงานการศกษาพบวาในเชอ Acinetobacter
spp. กระบวนการทรานสฟอรเมชนสามารถถายทอดยนดอยา blaOXA-23, blaOXA-24, blaOXA-58 และ blaNDM-1ไปส
แบคทเรยผรบได ท าใหเกดการแพรกระจายของยนดอยาภายในโรงพยาบาลเพมสงขน (Bertini et al., 2010; Rumbo
et al., 2011; Fu et al., 2012) สวนกระบวนการทรานสดกชน เปนการถายทอดยนดอยาจากแบคทเรยชนดหนงไปส
แบคทเรยอกชนด โดยอาศยแบคเทอรโอเฟจเปนตวนาพายนดอยา และกระบวนการคอนจเกชน เปนการถายทอดยนดอ
ยาจากแบคทเรยชนดหนงไปสแบคทเรยอกชนดโดยอาศยอวยวะพเศษทเรยกวา pili ดงเซลลแบคทเรยสองตวมาใกลกน
เกดการถายทอดยนดอยาใหแกกน มรายงานการศกษากอนหนาพบวาเชอ Acinetobacter spp. สามารถถายทอดยน
ดอยา blaOXA-23, blaOXA-58, blaGES-11, blaIMP-1 และ blaNDM-1 ไปสแบคทเรยผรบ โดยอาศยกระบวนการคอนจเกชน
(Bertini et al., 2010; Fu et al., 2012; Charfi-Kessis et al., 2014; Zhang et al., 2014; Yang et al., 2015)
จากการระบาดของโรคตดเชอทเกดจาก A. baumannii เพมสงขนจ านวนมากในระยะเวลาอนรวดเรว อกทง
เชอยงดอตอยาตานจลชพหลายขนานพรอมกน และยงพบการแพรกระจายของยนดอยาในวงกวาง ทงในทวปยโรป
ทวปอเมรกาใต และทวปเอเชย รวมทงประเทศไทยสงผลใหการรกษาโรคตดเชอท าไดยาก และท าใหผปวยเกดการ
เสยชวตในอตราสง จงน าไปสวตถประสงคของงานวจย คอ เพอศกษารปแบบการดอยา ตรวจหายนดอยาและการ
แพรกระจายของยนดอยาภายในเชอ A. baumannii ทแยกไดจากโรงพยาบาล 4 แหงในประเทศไทยและโรงพยาบาล
1 แหงจากประเทศเนปาล จากนนคดเลอกเชอทดอยาแบบรนแรง (XDR-AB) น ามาศกษาการถายทอดยนดอยาในแนบ
ราบ โดยวธทรานสฟอรเมชน, ทรานสดกชน และคอนจเกชน

45
วธด าเนนการวจย (Material and Method)
1. สายพนธแบคทเรยและแบคเทอรโอเฟจทใชในการศกษา
สายพนธแบคทเรยทใชในการศกษาครงน ไดแก Acinetobacter spp. จ านวน 347 ไอโซเลตโดยจดเปนเชอ A.
baumannii จ านวน 339 ไอโซเลตทแยกไดจากผปวยโรงพยาบาลสวรรคประชารกษจงหวดนครสวรรค โรงพยาบาล
เชยงรายประชานเคราะห จงหวดเชยงราย โรงพยาบาลพจตร จงหวดพจตร โรงพยาบาลจากจงหวดฉะเชงเทรา ประเทศ
ไทย และเชอ A. baumannii จ านวน 44 ไอโซเลตทแยกไดจากผปวยโรงพยาบาลของ Tribhuvan University ประเทศ
เนปาล และ Acinetobacter spp. จ านวน 19 ไอโซเลตทแยกจากแหลงน าธรรมชาตภายในจงหวดพษณโลก สวน
แบคเทอรโอเฟจทใชในการศกษา ไดแก bacteriophage ØABP-02, ØABP-19, ØABP-39, ØABP-44 ทจ าเพาะตอเชอ
A. baumannii ทแยกจากบอบ าบดน าเสย โรงพยาบาลพทธชนราช จงหวดพษณโลกและ bacteriophage ØABP-29 ท
จ าเพาะตอเชอ A. baumannii ทแยกจากบอบ าบดน าเสย โรงพยาบาลบางระก า จงหวดพษณโลก (ธวชชย กตต., 2555)
ดงแสดงในตาราง 7
ตาราง 7 แสดงสายพนธของแบคเทอรโอเฟจทใชในการศกษา
แบคเทอรโอเฟจ สายพนธโฮสต
A. baumannii
แหลงทเกบ
(บอบ าบดน าเสย)
แหลงทมา
ØABP-02 AB22 รพ.พทธชนราช ธวชชย กตต., 2555
ØABP-19 AB22 รพ.พทธชนราช ธวชชย กตต., 2555
ØABP-29 AB22 รพ. บางระก า ธวชชย กตต., 2555
ØABP-39 AB22 รพ.พทธชนราช ธวชชย กตต., 2555
ØABP-44 AB20 รพ.พทธชนราช ธวชชย กตต., 2555
2. การตรวจหาสายพนธของเชอธรรมชาตโดยการวเคราะหล าดบนวคลโอไทดของยน 16s rRNA หรอ rpoB
น าเชอ Acinetobacter spp. จากธรรมชาตมาเพมปรมาณดเอนเอโดยใชเทคนค PCR ดวย primer 16s rRNA-Fและ
16s rRNA-R หรอ rpoB-F และ rpoB-B ดงแสดงตาราง 8 เมอได PCR product แลวท าให PCR product ทไดมความ

46
บรสทธ โดยใชชดสกดส าเรจรป Gel/PCR DNA Fragment extraction (RBC Bioscience, Taipei, Taiwan) สงไป
วเคราะหล าดบนวคลโอไทดทบรษท Macrogen Inc. Service ประเทศเกาหล เมอไดล าดบนวคลโอไทดมาแลวน าล าดบ
นวคลโอไทดทไดมาเปรยบเทยบกบล าดบนวคลโอไทดของสงมชวตอนๆในฐานขอมล NCBI โดยใชโปรแกรม BLASTn
(https://blast.ncbi.nlm.nih.gov/Blast.cgi)
ตาราง 8 แสดงไพรเมอรทใชในการตรวจหายน 16s rRNA หรอยน rpoB
ไพรเมอร ล าดบนวคลโอไทด (5’ 3’) ขนาด PCR product (bp)
อางอง
rpoB-F CCTTCATGACCTGGAAYGGNTA 940 Wang et al., 2014 rpoB-B TCCAGGATCTGNCCNACRTTCAT 16s rRNA-F AGAGTTTGATCCTGGCTCAG 1500 Misbah et al., 16s rRNA-R ACGGCTACCTTGTTACGACTT 2005
3. การทดสอบความไวตอยาตานจลชพโดยวธ disc diffusion
เพาะเลยงเชอแบคทเรย A. baumannii บนจานอาหารเลยงเชอ TSA แลวน าไปบมท 37 องศาเซลเซยส เปนเวลา 24
ชวโมง จากนนน าโคโลนทเจรญบนจานอาหารเลยงเชอมาปรบความขนในสารละลาย 0.85%NaCl ใหไดความขน
เทยบเทากบ 0.5 Mcfarland โดยใชเครองวดความขน (McFarland densitometer: SiaBiosan, Latvia) น าไมพนส าล
ทปราศจากเชอจมลงในเชอทปรบความขนแลว ใหหมาดบรเวณขางหลอดทดลอง จากนนน าเชอมาปาย (swab) บนจาน
อาหารเลยงเชอ Mueller Hhinton agar (MHA) 3 ระนาบใหทวบรเวณผวหนาจานอาหาร รอใหเชอทปายบนอาหาร
เลยงเชอแหง แลวจงน ายาตานจลชพทตองการทดสอบความไวตอยาตานจลชพมาวางบนจานอาหารเลยงเชอ MHA ทท า
การปายเชอทดสอบแลว โดยใชเทคนคปราศจากเชอ จากนนน าไปบมท 37 องศาเซลเซยส เปนเวลา 24 ชวโมง แลว
บนทกผลการทดสอบความไวตอยาตานจลชพโดยการวด zone of inhibition (mm) และแปลผลการทดสอบความไวตอ
ยาตานจลชพโดยใชเกณฑมาตรฐานจาก Clinical and Laboratory Standards Institute (CLSI) antimicrobial
susceptibility testing standards 2014 ดงแสดงในตาราง 1
47
4. การตรวจหายนดอยา
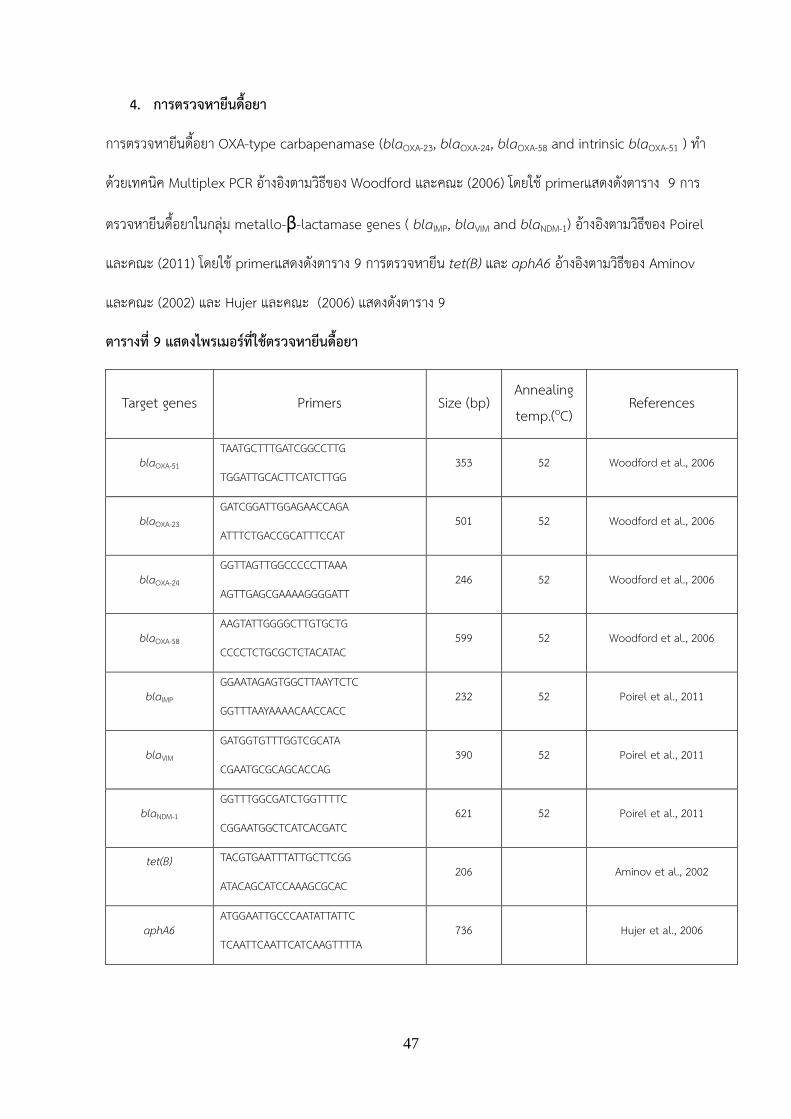
การตรวจหายนดอยา OXA-type carbapenamase (blaOXA-23, blaOXA-24, blaOXA-58 and intrinsic blaOXA-51 ) ท า
ดวยเทคนค Multiplex PCR อางองตามวธของ Woodford และคณะ (2006) โดยใช primerแสดงดงตาราง 9 การ
ตรวจหายนดอยาในกลม metallo-β-lactamase genes ( blaIMP, blaVIM and blaNDM-1) อางองตามวธของ Poirel
และคณะ (2011) โดยใช primerแสดงดงตาราง 9 การตรวจหายน tet(B) และ aphA6 อางองตามวธของ Aminov
และคณะ (2002) และ Hujer และคณะ (2006) แสดงดงตาราง 9
ตารางท 9 แสดงไพรเมอรทใชตรวจหายนดอยา
Target genes Primers Size (bp) Annealing temp.(oC)
References
blaOXA-51 TAATGCTTTGATCGGCCTTG
TGGATTGCACTTCATCTTGG 353 52 Woodford et al., 2006
blaOXA-23 GATCGGATTGGAGAACCAGA
ATTTCTGACCGCATTTCCAT 501 52 Woodford et al., 2006
blaOXA-24 GGTTAGTTGGCCCCCTTAAA
AGTTGAGCGAAAAGGGGATT 246 52 Woodford et al., 2006
blaOXA-58 AAGTATTGGGGCTTGTGCTG
CCCCTCTGCGCTCTACATAC 599 52 Woodford et al., 2006
blaIMP GGAATAGAGTGGCTTAAYTCTC
GGTTTAAYAAAACAACCACC 232 52 Poirel et al., 2011
blaVIM GATGGTGTTTGGTCGCATA
CGAATGCGCAGCACCAG 390 52 Poirel et al., 2011
blaNDM-1 GGTTTGGCGATCTGGTTTTC
CGGAATGGCTCATCACGATC 621 52 Poirel et al., 2011
tet(B)
TACGTGAATTTATTGCTTCGG
ATACAGCATCCAAAGCGCAC 206 Aminov et al., 2002
aphA6 ATGGAATTGCCCAATATTATTC
TCAATTCAATTCATCAAGTTTTA 736 Hujer et al., 2006

48
5. การศกษาการสงผานยนดอยาโดยวธทรานสฟอรเมชนแบบธรรมชาต (Natural transformation)
5.1 การสกดพลาสมดของเชอ extensively drug resistant A. baumannii (XDR-AB)
น าเชอ A. baumannii ซงท าการทดสอบหาความไวตอยาปฏชวนะแลวพบวาเชอดอตอยาตานจลชพทกชนดทใชในการ
ทดสอบ ยกเวน colistin และ tigecycline (XDR-AB) มาเลยงในอาหารเลยงเชอเหลว LB ทอณหภม 37 องศาเซลเซยส
เขยาดวยความเรวรอบ 150 รอบตอนาท เปนเวลา 24 ชวโมง จากนนน าไปปนเหวยงดวยความเรว 4,000 รอบตอนาท
เปนเวลา 15 นาท เพอตกตะกอนเซลลแบคทเรย เทสวนใสดานบนทง แลวน าตะกอนเซลลแบคทเรยทไดมาท าการสกดพ
ลาสมดดวยชดสกดส าเรจรป Plasmid Mini Kit (RBC Bioscience, Taipei, Taiwan) จากนนท าการตรวจสอบรปแบบ
ของพลาสมดทสกดไดโดยใชเทคนคอะกาโรสเจลอเลกโตรโฟเรซส (agarose gel electrophoresis)
5.2 การศกษาการสงผานยนดอยาโดยวธทรานสฟอรเมชน
เพาะเลยงเชอ A. baumannii ATCC19606 ทอณหภม 37 องศาเซลเซยส เขยาดวยความเรวรอบ 150 รอบตอนาท เปน
เวลา 3-4 ชวโมง จากนนน าพลาสมดของแบคทเรย XDR-AB ทคดเลอกมาจ านวน 10 ไอโซเลต ความเขมขน 1 ไมโครกรม
ผสมกบอาหารเลยงเชอเหลว LB ปรมาตร 500 ไมโครลตร และ O/N culture ของเชอ A. baumannii ATCC19606
ปรมาตร 500 ไมโครลตร น าไปบมท 37 องศาเซลเซยส เปนเวลา 1 ชวโมง แลวจงน ามาท าการ spread plate บนจาน
อาหารเลยงเชอแขง LB ทมยาตานจลชพ 3 ชนด ไดแก ticarcillin (1µg/ml) , kanamycin (20µg/ml) และ
tetracycline (20µg/ml) ตามล าดบ จานอาหารเลยงเชอละ 200 ไมโครลตร จากนนน าไปบมท 37 องศาเซลเซยส เปน
เวลา 24 ชวโมง ตรวจนบโคโลนทเจรญบนจานอาหารเลยงเชอ แลวน ามาค านวณหาประสทธภาพของการทรานสฟอร
เมชน (transformation efficiency) ในหนวย cfu/µg (Tu et al., 2005) โดยเรมจากการหา transformant ในหนวย
cfu ซงค านวณไดจากสตร
Transformant cfu = No. of bacteria colonies x original transformation volume plated volume เมอไดจ านวนของ transformant ในหนวย cfu ออกมาแลว น ามาค านวณหาคา transformation efficiency ตอ โดย
ค านวณจากสตร
Transformation efficiency (cfu/µg) = Transformant cfu/plasmid DNA (µg)

49
6. การศกษาการสงผานยนดอยาโดยวธทรานสดกชน
6.1 การเพมจ านวนแบคเทอรโอเฟจทใชในการศกษาและการตรวจนบปรมาณแบคเทอรโอเฟจ โดยวธ
plaque assay
การเพมจ านวนแบคเทอรโอเฟจทใชในการศกษาโดยเพาะเลยงเซลลโฮตสของแบคทเรย A. baumannii ในอาหารเลยง
เชอเหลว LB นาท เปนเวลา 24 ชวโมง จากนนน าเซลลโฮสตปรมาตร 5 มลลลตรใสลงไปในอาหารเลยงเชอเหลว LB
ปรมาตร 50 มลลลตรเขยาดวยความเรวรอบ 150 รอบตอนาทท 37 องศาเซลเซยส บมจนกวาเชอจะเจรญอยในระยะ
log phase หรอมคาดดกลนแสงทความยาวคลน 600 นาโนเมตร เทากบ 0.3-0.4 จากนนเตมแบคเทอรโอเฟจทจ าเพาะ
กบเซลลโฮสตชนดนนๆลงไป 1 มลลลตร แลวเขยาดวยความเรวรอบ 150 รอบตอนาท ท 37 องศาเซลเซยส เปนเวลา
2 ชวโมง หรอจนกระทงอาหารเลยงเชอใส จากนนเตม chloroform ปรมาตร 1 มลลลตร เปนเวลา 20 นาท จากนน
น าไปปนเหวยงดวยความเรว 8,000 รอบตอนาท เปนเวลา 15 นาท ท 20 องศาเซลเซยส เพอแยกเอาตะกอนเซลล
โฮสตออกไป สวนอนภาคของแบคเทอรโอเฟจจะอยสวนใสดานบน น าเอาสวนใสดานบนมากรองผานแผนกรองขนาด
0.45 ไมโครเมตร เกบเอาสวนใสหรออนภาคของแบคเทอรโอเฟจทกรองผานแผนกรองไวทอณหภม 4 องศาเซลเซยส
เพอใชตรวจนบปรมาณแบคเทอรโอเฟจ โดยวธ plaque assay ตอไปโดยน าแบคเทอรโอเฟจทท าการศกษา ปรมาตร
100 ไมโครลตร มาเจอจางใน SM buffer ปรมาตร 900 ไมโครลตร ใหไดระดบความเจอจางท 10-1 - 10-8 หลงจากนน
น าแบคเทอรโอเฟจทท าการเจอจางใน SM buffer แลวปรมาตร 100 ไมโครลตร มาผสมใหเขากนกบเซลลโฮสตของ
แบคทเรย A. baumannii ปรมาตร 100 ไมโครลตร และสารละลาย CaCl2 ความเขมขน 200 mM ปรมาตร 100
ไมโครลตร จากนนน าสวนผสมทงหมดเตมลงในอาหาร soft agar ทหลอมละลายแลวปรมาตร 4 มลลลตร ผสมใหเขา
กน แลวเทสวนผสมทเขากนแลวเทลงบนอาหารเลยงเชอ TSA ทมวนรอยละ 1.5 ตงทงไวรอจนอาหารเลยงเชอแขงตว
น าไปบมทอณหภม 37 องศาเซลเซยส เปนเวลา 18-24 ชวโมง นบจ านวนพลาคทเกดขน แลวน ามาค านวณหา
ปรมาณแบคเทอรโอเฟจ โดยใชสตร
PFU/ml = จ านวนพลาค x สวนกลบของการเจอจาง (dilution factor)
ปรมาณแบคเทอรโอเฟจทใสในวนทเท (ml)

50
6.2 การศกษาคณสมบตจ าเพาะระหวางแบคเทอรโอเฟจทแยกไดจากแหลงธรรมชาต และเซลลโฮสต
XDR-AB โดยวธ spot test
ท าการเพาะเลยงเชอแบคทเรย Acinetobacter spp. ทไวตอยาตานจลชพทท าการทดสอบทง 15 ชนด ซงแยกไดจาก
แหลงธรรมชาตและ XDR-AB ในอาหารเลยงเชอเหลว LB น าไปบมทอณหภม 37 องศาเซลเซยส เขยาดวยความเรวรอบ
150 รอบตอนาท เปนเวลา 24 ชวโมง จากนนน าเชอทบมขามคนแลว ปรมาตร 100 ไมโครลตร เตมลงในอาหารเลยง
เชอ soft agar ทหลอมละลายแลวปรมาตร 4 มลลลตร ผสมเขากนโดยใช vortex จากนนเทสวนผสมทเขากนแลวลง
บนอาหารเลยงเชอ TSA ทมวนรอยละ 1.5 อยางรวดเรว ตงทงไวรอจนอาหารเลยงเชอแขงตว แลวจงหยด (spot) แบ
คเทอรโอเฟจทท าการศกษา ปรมาตร 2 ไมโครลตรตอแบคเทอรโอเฟจหนงตวลงบนจานอาหารเลยงเชอ รอจนกระทง
แบคเทอรโอเฟจทท าการหยดลงไปแหงสนท แลวจงน าไปบมทอณหภม 37 องศาเซลเซยส เปนเวลา 18-24 ชวโมง
ตรวจดโซนใส (clear zone) ทเกดขน
6.3 กระบวนการทรานสดกชน
คดเลอกแบคเทอรโอเฟจทมความจ าเพาะกบเซลลโฮสต XDR-AB แลวน าแบคเทอรโอเฟจมาเตรยม phage lysate ของ
แบคทเรยผใหโดยเพาะเลยงเซลลโฮตส XDR-AB ในอาหารเลยงเชอเหลว LB ทอณหภม 37 องศาเซลเซยส เขยาดวย
ความเรวรอบ 150 รอบตอนาท เปนเวลา 24 ชวโมง จากนนน าเซลลโฮสต XDR-AB ปรมาตร 5 มลลลตรใสลงไปใน
อาหารเลยงเชอเหลว LB ปรมาตร 50 มลลลตร เขยาดวยความเรวรอบ 150 รอบตอนาท ท 37 องศาเซลเซยส บมจนกวา
เชอจะเจรญอยในระยะ log phase หรอมคาดดกลนแสงทความยาวคลน 600 นาโนเมตร เทากบ 0.3-0.4 เตมแบคเทอ
รโอเฟจทจ าเพาะกบเซลลโฮสตชนดนนๆลงไป 1 มลลลตร แลวน ามาเขยาดวยความเรวรอบ 150 รอบตอนาท ทอณหภม
37 องศาเซลเซยส เปนเวลา 2 ชวโมง หรอจนกระทงอาหารเลยงเชอใส เตม chloroform ปรมาตร 1 มลลลตร น าไปบม
ตอโดยการเขยาดวยความเรวรอบ 150 รอบตอนาท ท 37 องศาเซลเซยส เปนเวลา 20 นาท เพอใหเซลลโฮสตแตกแลว
สามารถปลดปลอยอนภาคแบคเทอรโอเฟจออกมาไดดยงขน จากนนน าไปปนเหวยงดวยความเรว 8,000 รอบตอนาท
เปนเวลา 15 นาท ทอณหภม 20 องศาเซลเซยส เพอแยกเอาตะกอนเซลลโฮสตออกไป สวนอนภาคของแบคเทอรโอเฟ
จจะอยสวนใสดานบน น าเอาสวนใสดานบนมากรองผานแผนกรองขนาด 0.45 ไมโครเมตร เกบเอาสวนใสหรออนภาค
ของแบคเทอรโอเฟจทกรองผานแผนกรองแลว ไวทอณหภม 4 องศาเซลเซยส หลงจากนนท าการเพาะเลยงเชอ
Acinetobacter spp. ทแยกไดจากแหลงธรรมชาต ในอาหารเหลว LB น าไปบมทอณหภม 37 องศาเซลเซยส เขยาดวย

51
ความเรวรอบ 150 รอบตอนาท เปนเวลา 24 ชวโมง น าเชอแบคทเรยทเพาะเลยงได ปรมาตร 500 ไมโครลตร มาผสม
กบแบคเทอรโอเฟจ (104pfu/ml) ทเตรยมไวขนตน ปรมาตร 50 ไมโครลตร น าไปบมท 37 องศาเซลเซยส เปนเวลา 30
นาท หลงจากนนเตมอาหารเหลว LB ปรมาตร 10 มลลลตรลงไปผสม แลวน าไปบมท 37 องศาเซลเซยส เปนเวลา1-2
ชวโมง โดยมการเขยารวมดวย เมอครบเวลาทก าหนดแลว น าไปปนเหวยงทความเรวรอบ 12000 g เปนเวลา 3 นาท เท
สวนใสดานบนทง น าตะกอนเซลลทตกอยบรเวณดานลางมาละลายกลบในอาหารเหลว LB ปรมาตร 1 มลลลตร จากนน
น ามา spread plate บนอาหารเลยงเชอ MHA ทมยาตานจลชพ 4 ชนด ไดแก ticarcillin (1µg/ml) , kanamycin
(20µg/ml) และ tetracycline (20µg/ml) ตามล าดบ จานเลยงเชอละ 200 ไมโครลตร น าไปบมท 37 องศาเซลเซยส
เปนเวลา 24-48 ชวโมง ตรวจดโคโลนทเจรญบนอาหารเลยงเชอ โดยเชอทสามารถเจรญบนจานอาหารเลยงเชอนได
เรยกวา transductants
7. การศกษาการสงผานยนดอยาโดยวธคอนจเกชน
7.1 การคดเลอกแบคทเรยผรบทมคณสมบตดอตอ sodium azide เพอใชเปนแบคทเรยผรบ
น าแบคทเรย Acinetobacter spp. ทแยกไดจากธรรมชาตมาศกษาหาคา MIC ของ sodium azide จากนนน าเชอ
ดงกลาวมาเลยงในอาหารเหลว LB ทมสวนผสมของ sodium azide ในระดบ 0.5 x MIC บมทอณหภม 37 องศา
เซลเซยส ความเรวรอบ 150 rpm เปนเวลา 24 ชวโมง จากนนนา overnight culture ไปเลยงในอาหาร LB ทเพม
ความเขมขนของ sodium azide (เพมครงละ 0.5 x MIC) ในอตราสวน 1:10 (overnight culture: fresh LB) เมอ
แบคทเรยผรบสามารถเจรญในระดบความเขมขนของ sodium azide ตามทตองการแลว น ามาศกษา susceptibility
patterns ของทงยาตานจลชพและ sodium azide กอนน ามาใชเปนแบคทเรยผรบตอไป
7.2 กระบวนการคอนจเกชน
น าแบคทเรยผใหทเปน XDR-AB มาทดสอบการสงผานยนดอยาโดยวธคอนจเกชนกบแบคทเรยผรบ โดยคดเลอก
แบคทเรย XDR-AB ทมคณสมบตดอตอยาแตไวตอ sodium azide (MIC ≤ 150 μg/ml) มาศกษาการสงผานยนดอ
ยาโดยวธ conjugation โดยใช broth culture mating method เรมจากน าแบคทเรยผให (XDR-AB) และแบคทเรย
ผรบทมคณสมบตดอตอ sodium azide มาเพาะเลยงในอาหารเลยงเชอแขง LB บมท 37 องศาเซลเซยส เปนเวลา
ขามคน จากนนน าแบคทเรยผใหและแบคทเรยผรบ มาปรบปรมาณเซลลใหไดเทากบ 0.5 McFarland ในสารละลาย
0.85% NaCl น าแบคทเรยผใหและแบคทเรยผรบมาผสมกน ในอตราสวน 1:3 ชดควบคมใชแบคทเรยผรบปรมาตร 180

52
ไมโครลตร และ 0.85% NaCl ปรมาตร 60 ไมโครลตร ผสมลงในอาหารเลยงเชอเหลว LB ปรมาตร 760 ไมโครลตร
จากนนน าไปบมท 37 องศาเซลเซยส เปนเวลา 4 ชวโมง คดเลอก transconjugants บนอาหารเลยงเชอ MHA ทม 50
μg/ml ticarcillin + 300 μg/ml sodium azide อาหาร MHA ทม 20 μg/ml kanamycin + 300 μg/ml sodium
azide และอาหาร MHA ทม 20 μg/ml tetracycline + 300 μg/ml sodium azide โดยเชอทสามารถเจรญบนจาน
อาหารเลยงเชอนไดเรยกวา transconjugants จานควบคมจะท าการ spread แบคทเรยผรบบนจานอาหารเลยงเชอ
MHA ทม sodium azide 300 μg/ml และยาตานจลชพ 3 ชนด ไดแก ticarcillin (50 μg/ml) kanamycin (20
μg/ml) และ tetracycline (20 μg/ml) น าผลการทดลองทได มาค านวณหาคา frequency of conjugation โดยใช
สตรดงตอไปน
Frequency of conjugation = ปรมาณของ transconjugants (CFU/ml) ปรมาณของ recipient (CFU/ml)
8. การตรวจหายนดอยา class D β-lactamase, tet(B) และ aphA6 ใน transformants,
tranductants และ transconjugants ทแยกได
สกดดเอนเอของ transformants, transductants และ transconjugants ทแยกไดโดยใชวธ boiling method เมอ
ไดดเอนเอออกมาใชเปนดเอนเอแมแบบ (DNA template) ตรวจหายนดอยาโดยใชไพรเมอรทจ าเพาะกบยนในกลม
Class D carbapenemase ยนดอยา tetracycline (tet(B)) และยนดอยา kanamycin (aphA6) (ตาราง 8) จากนน
ท าการตรวจสอบดเอนเอทไดจากการเพมปรมาณ (PCR product) โดยใชเทคนคอะกาโรสเจลอเลกโตรโฟเรซส
(agarose gel electrophoresis)

53
ผลการวจย (Result)
1. การตรวจหาสายพนธของเชอธรรมชาตโดยการวเคราะหล าดบนวคลโอไทดของยน 16s rRNA หรอ rpoB
จากการตรวจหาสายพนธของเชอ Acinetobacter spp. ทแยกไดจากแหลงธรรมชาต จ านวน 19 ไอโซเลต โดยท าการ
เพมปรมาณดเอนเอดวยเทคนค PCR ไพรเมอรทใชในการศกษา ไดแก 16S rRNA-F, 16S rRNA-R, rpoB-F และ rpoB-
R พบวาเมอน าล าดบนวคลโอไทดทวเคราะหไดไปเปรยบเทยบกบล าดบนวคลโอไทดของสงมชวตอนๆในฐานขอมล
NCBI โดยใชโปรแกรม BLASTn พบวาเชอ Acinetobacter spp. ทแยกไดจากแหลงธรรมชาต จ านวน 19 ไอโซเลต
สามารถจ าแนกเปน 3 สายพนธ ไดแก A. baumannii จ านวน 9 ไอโซเลต Acinetobacter soli จ านวน 8 ไอโซเลต
และ Acinetobacter nosocomialis จ านวน 2 ไอโซเลต
2. การทดสอบความไวตอยาตานจลชพโดยวธ disc diffusion
ผลการท า Antibiotiogram ของเชอ A. baumannii clinical isolates โดยวธ disk diffusion method พบเชอมการ
ดอยาดงแสดงในตารางท 10 โดยเชอ A. baumannii clinical isolates จากโรงพยาบาลสวรรคประชารกษ พบเชอท
ไมดอตอยา จ านวน 15 ตวอยาง (21.73%) พบเชอทเปน Multidrug resistant A. baumannii (MDR-AB) 52
ตวอยาง (75.36%) carbapenem resistance A. baumannii (CR-AB) 45 ตวอยาง (65.21%) และ extensively
drug resistant A. bauamannii (XDR-AB) 8 ตวอยาง (21.11%) ผลการท า Antibiotiogram ของเชอ A.
baumannii clinical isolates จากโรงพยาบาลพจตร พบเชอทไมดอตอยา จ านวน 4 ตวอยาง (7.27%) พบเชอท
เปน MDR-AB 51 ตวอยาง (92.73%) CR-AB 43 ตวอยาง (78.18%) และ XDR-AB 4 ตวอยาง (7.27%) ผลการท า
Antibiotiogram ของเชอ A. baumannii clinical isolates จากโรงพยาบาลเชยงราย พบเชอทไมดอตอยา จ านวน 14
ตวอยาง (12.84%) พบเชอทเปน Multidrug resistant A. baumannii 95 ตวอยาง (87.16%) CR-AB 92 ตวอยาง
(84.40%) และ XDR-AB 6 ตวอยาง (5.5%) ผลการท า Antibiotiogram ของเชอ A. baumannii clinical isolates
จากโรงพยาบาลจงหวดฉะเชงเทรา ไมพบเชอทไมดอตอยา พบเชอทเปน Multidrug resistant A. baumannii 115
ตวอยาง (98.3%) CR-AB 110 ตวอยาง (94.01%) และ XDR-AB 1 ตวอยาง (2.27%) ผลการท า Antibiotiogram
ของเชอ A. baumannii clinical isolates จากโรงพยาบาล Tribhuvan University hospital พบเชอทไมดอตอยา
จ านวน 1 ตวอยาง (2.27%) พบเชอทเปน Multidrug resistant A. baumannii 43 ตวอยาง (97.7%) CR-AB 43

54
ตวอยาง (97.7%) และ XDR-AB 9 ตวอยาง (7.7%) สวนเชอจากธรรมชาตพบเชอทไมดอตอยา จ านวน 13 ตวอยาง
(68.42 %) พบเชอทเปน Multidrug resistant A. baumannii และ CR-AB เปน 5 ตวอยาง (29.4%)
ตารางท 10 ผล Antibiotiogram ของเชอ A. baumannii clinical isolates และ เชอAcinetobacter
spp. ทแยกจากธรรมชาต
ชอยา จ านวน (%) ของเชอทแยกจากผปวยในแตละโรงพยาบาลและแหลงน าธรรมชาต สวรรค
ประชารกษ (N=66)
พจตร (N=55)
เชยงรายประชานเคราะห (N=109)
จงหวดฉะเชงเทรา (N=117)
Tribhuvan University hospital (N=44)
Acinetobacter spp. จากธรรมชาต (N=19)
amikacin (AK) 39 (59.09) 30 (54.55) 54 (49.54) 41(35.04) 19(43.2) 3 (15.79) cefepime (FEP) 45 (68.18) 45 (81.82) 62 (56.88) 107(91.45) 39(88.6) 5 (26.32) Cefoperazone/ sulbactam (SCF)
20 (30.30) 11 (20.00) 14 (12.84) 4 (3.4) 10(22.72) 1 (5.26)
cefotaxime (CTX) 47 (71.21) 38 (69.10) 87 (79.82) 114(97.4) 43(97.7) 5 (26.32) ceftazidime (CAZ) 49 (74.24) 49 (89.10) 85 (77.98) 114(97.4) 42(95.4) 5 (26.32) ceftriaxone (CRO) 47 (71.21) 50 (90.91) 88 (80.73) 115(98.3) 41(93.2) 5 (26.32) ciprofloxacin (CIP) 50 (75.75) 51 (92.73) 89 (81.65) 112(95.72) 43(97.7) 5 (26.32) colistin (CT) 1 (1.51) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) gentamicin (CN) 49 (74.24) 31 (56.36) 70 (64.22) 39(33.33) 23 (52.3) 5 (26.32) imipenem (IPM) 42 (63.63) 39 (70.91) 90 (82.57) 110(94.01) 43(97.7) 4 (21.05) Meropenem (MEM) 42 (63.63) 41 (75.44) 91 (83.49) 110(94.01) 43(97.7) 5 (26.32) piperacillin/tazobactam (TZP)
45 (68.18) 47 (85.46) 86 (78.90) 112(95.72) 43(97.7) 5 (26.32)
tetracycline (TE) 26 (39.39) 40 (72.73) 76 (69.72) 90(76.92) 21(41.7) 2 (10.53) tigecycline (TGC) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) Trimethoprim/ sulfamethoxazole (TMX/SXT)
33 (50.0) 44 (80.00) 46 (42.20) 92(78.6) 41(93.2) 6 (31.58)
3. การตรวจหายนดอยา OXA-type carbapenamase และ blaNDM-1
ผลการตรวจหา antibiotic resistance genes กลม OXA-type carbapenamase ของเชอ A. baumannii clinical
isolates แสดงดงภาพ 7 เชอ A. baumannii clinical isolates จากโรงพยาบาลสวรรคประชารกษ พบยน blaOXA-
51 จ านวน 62 ตวอยาง (93.94%) blaOXA-23 จ านวน 45 ตวอยาง (68.18%) blaOXA-24 จ านวน 1 ตวอยาง (1.51%)

55
และ blaOXA-58 จ านวน 3 ตวอยาง ( 4.54%) โดยตรวจไมพบยน blaNDM-1, blaIMP, blaVIM (ตารางท 11) ผลการตรวจหา
antibiotic resistance genes ของเชอ A. baumannii clinical isolates โรงพยาบาลพจตรพบยน blaOXA-51 จ านวน
54 ตวอยาง (98.18%) blaOXA-23 จ านวน 41 ตวอยาง (74.54%) blaOXA-58 จ านวน 6 ตวอยาง (10.91%) โดยตรวจ
ไมพบ blaOXA-24, blaNDM-1, blaIMP, blaVIM (ตารางท 11) ผลการตรวจหา antibiotic resistance genes ของเชอ A.
baumannii clinical isolates โรงพยาบาลเชยงรายประชานเคราะห พบยน blaOXA-51 จ านวน 106 ตวอยาง
(97.25%) blaOXA-23 จ านวน 83 ตวอยาง (76.15%) blaOXA-58 จ านวน 8 ตวอยาง (7.34%) blaNDM-1 จ านวน 9 ตวอยาง
(8.2%) โดยตรวจไมพบ blaOXA-24 (ตารางท 11) ผลการตรวจหา antibiotic resistance genes ของเชอ A. baumannii
clinical isolates โรงพยาบาลฉะเชงเทรา พบยน blaOXA-51 จ านวน 117 ตวอยาง (100%) blaOXA-23 จ านวน 113
ตวอยาง (96.5%) blaOXA-58 จ านวน 3 ตวอยาง (2.56%) blaNDM-1 จ านวน 22 ตวอยาง (18.8%) blaVIM จ านวน 1
ตวอยาง (0.85%) โดยตรวจไมพบ blaOXA-24 และ blaIMP ผลการตรวจหา antibiotic resistance genes ของเชอ A.
baumannii clinical isolates โรงพยาบาล Tribhuvan University พบยน blaOXA-51 และ blaOXA-23 จ านวน 44
ตวอยาง (100%) blaNDM-1 จ านวน 6 ตวอยาง (13.6%) โดยตรวจไมพบ blaOXA-24, blaOXA-58, blaIMP, blaVIM ผลการ
ตรวจหา antibiotic resistance genes ของเชอ Acinetobacter spp. จากแหลงน าธรรมชาต พบยน blaOXA-51
จ านวน 14 ตวอยาง (73.68%) blaOXA-23 จ านวน 4 ตวอยาง (21.05 %) โดยตรวจไมพบยน blaNDM-1, blaOXA-24,
blaOXA-58, blaIMP, blaVIM (ตารางท 11)
ภาพ 7 แสดงผล PCR product ของ antibiotic resistance genes

56
แถว M คอ molecular weight marker แถว 1 คอ blaOXA51 แถว 2 คอ blaoxa23 และ blaOXA51 แถว 3 คอ
blaoxa24 และ blaOXA51 แถว 4 คอ blaoxa58 และ blaOXA51.
ตารางท 11 แสดงผลการตรวจหา antibiotic resistance genes ของเชอ A. baumannii clinical isolates
และเชอ Acinetobacter spp. ทแยกจากธรรมชาต
ยนดอยา โรงพยาบาล
สวรรค
ประชารกษ
(N=66)
โรงพยาบาล
พจตร
(N=55)
โรงพยาบาล
เชยงราย
ประชาน
เคราะห
(N=109)
โรงพยาบาล
จงหวด
ฉะเชงเทรา
(N=117)
โรงพยาบาล
Tribhuvan
University
(N=44)
Acinetobacter
spp. จาก
ธรรมชาต
(N=19)
blaOXA-51 62 (93.9) 54 (98.1) 106(97.25) 117(100) 44(100) 14 (73.6)
blaOXA-23 45 (68.1) 41 (74.5) 83(76.15) 113(96.5) 44(100) 4 (21.0)
blaOXA-24 1 (1.5) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0)
blaOXA-58 3 (4.5) 6 (10.9) 8(7.34) 3(2.56) 0 (0) 0 (0)
blaNDM-1 0 (0) 0 (0) 9(8.2) 22(18.8) 6(13.6) 0 (0)
blaIMP 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0)
blaVIM 0 (0) 0 (0) 0 (0) 1 (0.85) 0 (0) 0 (0)
4. การคดเลอกแบคทเรยผรบและแบคทเรยผให
ท าการคดเลอกเชอแบคทเรยผให โดยการทดสอบความไวตอยาตานจลชพ จ านวน 14 ชนด จากเชอ A. baumannii ท
แยกไดจากผปวย คดเลอกเชอ A. baumannii ทดอตอยาตานจลชพ ในรปแบบ XDR-AB หรอ blaNDM-1 postive
strains จ านวน 14 ไอโซเลตน ามาตรวจหายนดอยา OXA-type carbapenemase ยนดอยา tetracycline (tet(B)) และ
ยนดอยา kanamycin (aphA6) พบวาเชอทง 14 ไอโซเลต สามารถตรวจพบยนดอยา blaOXA-23 ในทกไอโซเลต และยน
ดอยา tet(B) 12 ไอโซเลต ส าหรบยนดอยา aphA6 ตรวจพบ 4 ไอโซเลต ดงแสดงในตาราง 12 สวนในแบคทเรยผรบ

57
คดเลอก จากเชอ Acinetobacter spp. ทแยกไดจากแหลงธรรมชาต คดเลอกเฉพาะสายพนธทไวตอยาตานจลชพทก
ชนด (no resistance) แลวท าการตรวจหายนดอยาชนดเดยวกบแบคทเรยผให จากการศกษาพบวาไดเชอ
Acinetobacter spp. จ านวน 4 ไอโซเลต ไดแก สายพนธ NU005-1, NU013, NU015 และ NI003 ทไวตอยาตานจล
ชพทกชนดทใชในการทดสอบ และตรวจไมพบยนดอยา OXA-type carbapenemase ยนดอยา tet(B) และยนดอยา
aphA6 เมอคดเลอกแบคทเรยผใหและแบคทเรยผรบแลว ท าการศกษาการถายทอดยนดอยาในแนวราบ โดยอาศย
กระบวนการ ทรานสฟอรเมชน ทรานสดกชน และคอนจเกชนตอไป
ตาราง 12 แสดงผลการคดเลอกแบคทเรยผใหและแบคทเรยผรบ
Strains Source of isolates
Antibiotic resistance genes
blaOXA-23 blaNDM-1 tet(B) aphA6 blaPER-1 AB135 pus + - + - - AB136 urine + - + - + AB140 sputum + - + + - AB229 urine + - + - - AB095 sputum + - + - - AB180 sputum + - + - - AB183 sputum + - + - - AB241 sputum + - + - - AB246 sputum + - + - - AB269 sputum + - + - - AB120 sputum + + + - - AB352 sputum + + - + + AB364 ascitic swab + + + + + AB405 sputum + + - + +
5. การศกษาการสงผานยนดอยาโดยวธทรานสฟอรเมชนแบบธรรมชาต (Natural transformation)
การศกษาโดยการน าพลาสมดของเชอ XDR-AB ซงใชเปนแบคทเรยผให จ านวน 10 ไอโซเลต มาท าการทดสอบการ
สงผานยนดอยาโดยอาศยกระบวนการทรานสฟอรเมชนแบบธรรมชาต (Natural transformation) ไปสเชอแบคทเรย
ผรบ คอ Acinetobacter spp. ทแยกมาจากแหลงธรรมชาต และไวตอยาตานจลชพทกชนดทใชในการทดสอบ จ านวน
4 ไอโซเลต พบวา เชอแบคทเรยผให XDR-AB สายพนธ AB135, AB136, AB140, AB180, AB183 และ AB241 สามารถ

58
สงผานการดอยา kanamycin ไปสแบคทเรยผรบสายพนธ NU015 ได สงผลใหตรวจพบโคโลนเจรญบนจานอาหารเลยง
เชอทสวนประกอบของยา kanamycin 20 μg/ml สวนอาหารอนไมพบ transformant เมอค านวณหาคาความถในการ
เกดกระบวนการทรานสฟอรเมชนแบบธรรมชาต (Frequency of transformation) พบวาอยในชวง 1.1 x 10-10 ถง 4.4
x 10-11 สวนคาประสทธภาพของการทรานสฟอรเมชน (Transformation efficiency) อยในชวง 2 x 101 ถง 8 x 102
CFU/μg (ตาราง 13) เมอน าโคโลนของ transformant ทไดมาตรวจหายนดอยา kanamycin (aphA6) จากการศกษา
พบวาตรวจไมพบยนดอยา aphA6 ใน transformants ทได
6. การศกษาการสงผานยนดอยาโดยวธทรานสดกชน
การศกษาการสงผานยนดอยาโดยอาศยกระบวนการทรานสดกชน เรมจากการเพมจ านวนแบคเทอรโอเฟจ จ านวน
5 สายพนธ ไดแก ØABP-02, ØABP-19, ØABP-29, ØABP-39 และ ØABP-44 จากนนน าแบคเทอรโอเฟจทท าการเพม
จ านวนไดมาท าการนบจ านวน โดยใชวธ plaque assay แลวศกษาคณสมบตความจ าเพาะระหวางแบคเทอรโอเฟจกบ
เซลลโฮสต (XDR-AB) โดยวธ spot test จากการศกษาพบวาแบคเทอรโอเฟจ ØABP-02 และ ØABP-39 มความจ าเพาะ
กบเชอ XDR-AB สายพนธ AB183 สวนแบคเทอรโอเฟจ ØABP-19, ØABP-29 และ ØABP-44 มความจ าเพาะกบเชอ
XDR-AB สายพนธ AB135 และ AB229 น าเชอ XDR-AB สายพนธทมความจ าเพาะกบแบคเทอรโอเฟจนนๆมาใชเปน
เซลลโฮสตในการเพมจ านวนและน าพายนดอยาไปสแบคทเรยผรบ จากการศกษาการสงผานยนดอยาโดยวธทรานส
ดกชน พบวาเชอ XDR-AB สายพนธ AB135 ทใชแบคเทอรโอเฟจ ABP19 (ØABP-19) เปนตวน าพาการดอยา สามารถ
น าพาการดอยา tetracycline ไปสเชอแบคทเรยผรบสายพนธ NU013 และ NU015 จงตรวจพบโคโลนเจรญบนจาน
อาหารเลยงเชอทมสวนประกอบของยา tetracycline 20 μg/ml เมอน ามาค านวณหาคาความถในการเกด
กระบวนการทรานสดกชน (Frequency of transduction) โดยอาศยแบคเทอรโอเฟจเปนตวน าพายนดอยานน พบวาม
คาอยในชวง 4.8 x 10-9 ถง 9.1 x 10-9 หลงจากนนท าการตรวจหายนดอยา tetracycline (tet(B)) ใน transductants
ทได (ตาราง 14) จากการศกษาพบวาตรวจพบยนดอยา tet(B) ในทกไอโซเลตของ transductants ดงแสดงใน ภาพ 8

59
ภาพ 8 แสดง PCR product จากการตรวจสอบหายนดอยา tet(B) ใน transductants ทไดจากกระบวนการ ทรานสดกชน Lane M: 100 bp ladder; Lane 1: Negative control (น ากลน); Lane 2: PCR product ยนดอยา blaOXA-51 ขนาด 353 bp และยนดอยา tet(B) ขนาด 206 bp ของแบคทเรยผใหสายพนธ AB135; Lane 3-4: PCR product ยนดอยา blaOXA-51 ของแบคทเรยผรบ สายพนธ NU013 และ NU015 ตามล าดบ และ Lane 5-8: PCR product ยนดอยา blaOXA-51 และ tet(B) ของ transductants ทไดจ านวน 4 ไอโซเลต
7. การศกษาการสงผานยนดอยาโดยวธคอนจเกชน
คดเลอกแบคทเรยผรบโดยการทดสอบความไวตอยาตานจลชพ (susceptibility patterns) และความสามารถในการ
เจรญในสภาวะทม sodium azide เขมขนมากกวา 300 μg/ml จากการศกษาพบวาสามารถคดเลอกแบคทเรยผรบได
จ านวน 2 ไอโซเลต คอ NU013 และ NU015 ทไวตอยาตานจลชพทกชนด และมคา MIC ของ sodium azide มากกวา
300 μg/ml เมอไดแบคทเรยผรบแลว น ามาท าการทดลองการสงผานยนดอยาโดยวธการคอนจเกชน ผลการศกษา
พบวาสามารถตรวจพบโคโลนเจรญบนอาหารเลยงเชอทมยาตานจลชพ ticarcilin 50 μg/ml และ kanamycin 20
μg/ml (transconjugants) ซงใชเชอ XDR-AB สายพนธ AB140 เปนแบคทเรยผให และ Acinetobacter spp. สาย
พนธ NU013 และ NU015 เปนแบคทเรยผรบ เมอน าโคโลนของ transconjugants ทเจรญบนจานอาหารเลยงเชอทม
ยาตานจลชพ ticarcillin 50 μg/ml มาท าการตรวจหายนดอยาในกลม OXA-type carbapenemase พบวาตรวจไม
พบยนดอยาในกลม OXA-type carbapenemase แตตรวจพบยน blaPER-1 สวน transconjugants ทเจรญบนอาหาร
เลยงเชอทมยาตานจลชพ kanamycin 20 μg/ml เมอน ามาตรวจหายนดอยา aphA6 พบวาสามารถตรวจพบยนดอยา
aphA6 ใน transconjugants ดงภาพ 9 แสดงใหเหนวาเชอ XDR-AB สายพนธ AB140 ซงเปนแบคทเรยผใหสามารถ
เกดการสงผานยนดอยา aphA6 ไปสแบคทเรย ผรบสายพนธ NU013 และ NU015 สงผลใหเชอแบคทเรยผรบดอตอยา

60
kanamycin ตามมา (ตาราง 15) จากนนค านวณหาคา frequency of conjugation (CF) พบวาคา CF มคาเทากบ 1.0
x 10-4 – 8.6 x 10-5
ภาพ 9 แสดง PCR product จากการตรวจสอบหายนดอยา aphA6 ภายใน transconjugants ทไดจากกระบวนการคอนจเกชน Lane M: 100 bp ladder; Lane 1: Negative control (น ากลน); Lane 2: PCR product ยนดอยา blaOXA-51 ขนาด 353 bp และ ยนดอยา aphA6 ขนาด 736 bp ของแบคทเรยผใหสายพนธ AB140; Lane 3-4: PCR product ยนดอยา blaOXA-51 ของแบคทเรยผรบ สายพนธ NU013 และ NU015 ตามล าดบ และ Lane 5-6: PCR product ยนดอยา blaOXA-51 และ aphA6 ของ transconjugants ทได
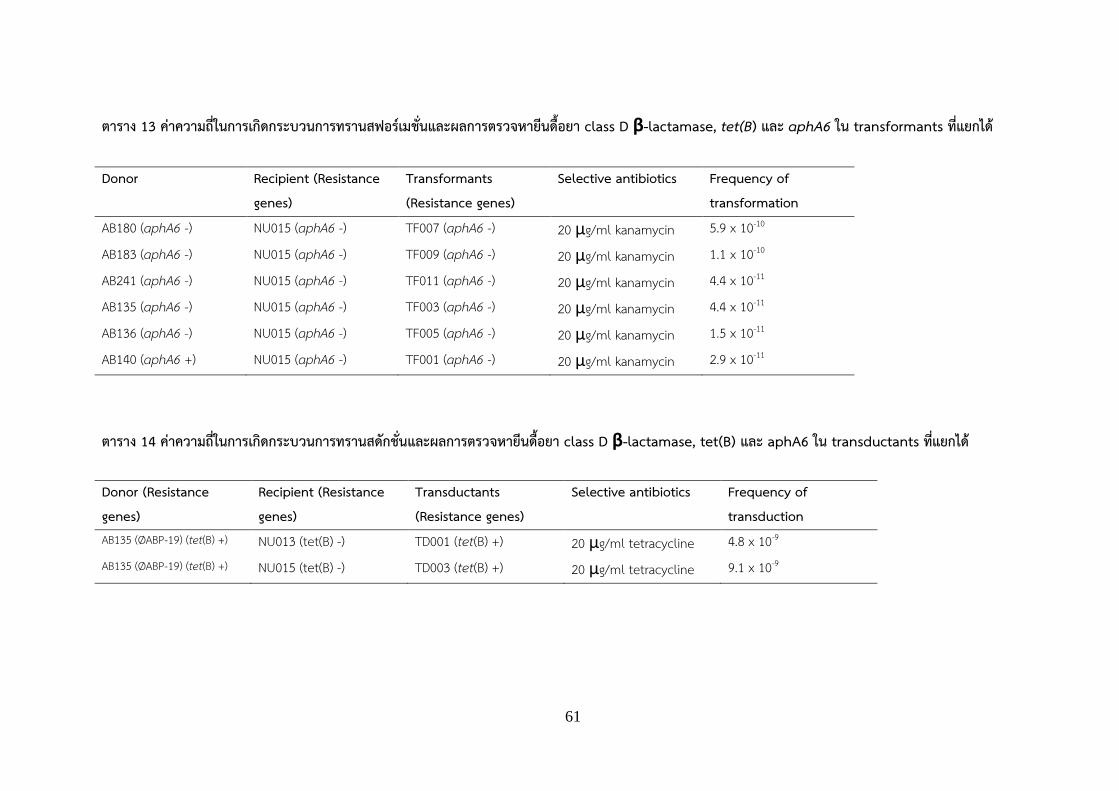
61
ตาราง 13 คาความถในการเกดกระบวนการทรานสฟอรเมชนและผลการตรวจหายนดอยา class D β-lactamase, tet(B) และ aphA6 ใน transformants ทแยกได Donor Recipient (Resistance
genes) Transformants (Resistance genes)
Selective antibiotics Frequency of transformation
AB180 (aphA6 -) NU015 (aphA6 -) TF007 (aphA6 -) 20 μg/ml kanamycin 5.9 x 10-10
AB183 (aphA6 -) NU015 (aphA6 -) TF009 (aphA6 -) 20 μg/ml kanamycin 1.1 x 10-10
AB241 (aphA6 -) NU015 (aphA6 -) TF011 (aphA6 -) 20 μg/ml kanamycin 4.4 x 10-11
AB135 (aphA6 -) NU015 (aphA6 -) TF003 (aphA6 -) 20 μg/ml kanamycin 4.4 x 10-11
AB136 (aphA6 -) NU015 (aphA6 -) TF005 (aphA6 -) 20 μg/ml kanamycin 1.5 x 10-11
AB140 (aphA6 +) NU015 (aphA6 -) TF001 (aphA6 -) 20 μg/ml kanamycin 2.9 x 10-11
ตาราง 14 คาความถในการเกดกระบวนการทรานสดกชนและผลการตรวจหายนดอยา class D β-lactamase, tet(B) และ aphA6 ใน transductants ทแยกได Donor (Resistance genes)
Recipient (Resistance genes)
Transductants (Resistance genes)
Selective antibiotics Frequency of transduction
AB135 (ØABP-19) (tet(B) +) NU013 (tet(B) -) TD001 (tet(B) +) 20 μg/ml tetracycline 4.8 x 10-9 AB135 (ØABP-19) (tet(B) +) NU015 (tet(B) -) TD003 (tet(B) +) 20 μg/ml tetracycline 9.1 x 10-9

62
ตาราง 15 คาความถในการเกดกระบวนการคอนจเกชนและผลการตรวจหายนดอยา class D β-lactamase, tet(B) และ aphA6 ใน transconjugants ทแยกได Donor (Resistance genes)
Recipient (Resistance genes)
Transconjugants (Resistance genes)
Selective antibiotics Frequency of conjugation
AB140 (blaOXA-23 +) NU013 R (blaOXA-23 -) TC011 (blaOXA-23 -, blaPER-1 )
50 μg/ml ticarcillin 1.0 x 10-4
AB140 (blaOXA-23 +) NU015 R (blaOXA-23 -) TC016 (blaOXA-23 - blaPER-1)
50 μg/ml ticarcillin 8.6 x 10-5
AB140 (aphA6 +) NU013 R (aphA6 -) TC021 (aphA6 +) 20 μg/ml kanamycin 8.5 x 10-5
AB140 (aphA6 +) NU015 R (aphA6 -) TC026 (aphA6 +) 20 μg/ml kanamycin 6.3 x 10-5

63
ขอวจารณ (Discussion) 1. รปแบบการดอยาของเชอ A. baumannii
ในชวง 10 ปทผานมา พบอบตการณการดอตอยาตานจลชพของเชอ A. baumannii เพมสงขนอยางรวดเรว จาก
การศกษาความไว ตอยาตานจลชพ (susceptibility patterns) ของเชอ A. baumannii ทแยกไดจากสงสงตรวจภายใน
โรงพยาบาล 5 แหงในประเทศไทยและประเทศเนปาลพบวาเชอ A. baumannii ดอตอยาตานจลชพ cefotaxime
(CTX), ceftazidime (CAZ), ceftriaxone (CRO), ciprofloxacin (CIP), imipenem (IPM), meropenem (MEM)
และ piperacillin/tazobactam (PIP) มากกวารอยละ75 โดยทกไอโซเลตไวตอยาตานจลชพ colistin และ tigecycline
เมอน ามาจดรปแบบการดอตอยาตานจลชพ พบวา A. baumannii จดอยในรปแบบของ MDR-AB มากทสด คดเปนรอย
ละ 75-98 รองลงมา ไดแก CR-AB รอยละ 65-98 และ XDR-AB รอยละ 5.5-21 ตามล าดบ เปรยบเทยบกบการศกษา
ของ Aimsaad et al. ในป ค.ศ. 2009 พบอบตการณการดอตอยาตานจลชพตงแต สามขนานขนไปของเชอ A.
baumannii (MDR-AB) ในโรงพยาบาลพระมงกฎเกลา คดเปนรอยละ 21.1 (Aimsaad et al., 2009) ตอมาในป ค.ศ.
2014 ภายในโรงพยาบาลศรราช พบอบตการณของ MDR-AB เพมเปนรอยละ 88.7 (Chaisathaphol and
Chayakulkeeree, 2014) สวนรปแบบการดอตอยาตานจลชพของเชอ A. baumannii แบบ XDR-AB มผศกษาในป
ค.ศ. 2011 พบอบตการณของ XDR-AB คดเปนรอยละ 30.6 (Anunnatsiri and Tonsawan, 2011) สวนในป ค.ศ.
2015 ภายในโรงพยาบาลมหาวทยาลยเชยงใหม พบวาเชอ A. baumannii ทแยกไดจากสงสงตรวจทงหมด ดอตอยา
ตานจลชพทกชนดทใชในการทดสอบ ยกเวน colistin และ tigecycline คดเปนรอยละ 65.3 (Inchai et al., 2015) และ
อบตการณการดอตอยาตานจลชพในกลมคารบาพเนมของเชอ A. baumannii (CR-AB) ภายในประเทศไทยยงเพม
สงขน จากรอยละ 46.7 ในป ค.ศ. 2009 เปนรอยละ 81.8 ในป ค.ศ. 2012 (Dejsirilert et al., 2012; Werarak et al.,
2012) ซงจาก ผลการศกษาในครงน แสดงใหเหนวาเชอ A. baumannii ทแยกไดจากโรงพยาบาลบรเวณภาคเหนอของ
ประเทศไทยในปจจบนนน มอบตการณการดอตอยาตานจลชพเปนจานวนมาก สงผลใหจ าเปนตองตระหนกในเรองของ
การใชยาตานจลชพในการรกษาโรคตดเชอใหเหมาะสมมากยงขน เพอชวยลดอบตการณการดอตอยาตานจลชพใน
อนาคตตอไป

64
2. การตรวจหายนดอยาของเชอ A. baumannii
ผลการตรวจสอบหายนดอยาในกลม acquired OXA-type carbapenemase ซงไดแกยนดอยา blaOXA-23, blaOXA-24
และ blaOXA-58 ของเชอ A. baumannii ทแยกไดผปวยจากการศกษาพบวาตรวจพบยนดอยา blaOXA-23 มากทสดในทก
โรงพยาบาล คดเปนรอยละ 68-100 (ตาราง 11) สอดคลองกบการศกษาทงในประเทศไทยและในทวปเอเชย ทพบ
อบตการณของยนดอยา blaOXA-23 มากทสด อยระหวางรอยละ 80-100 ในเชอ A. baumannii ทแยกไดจาก สงสงตรวจ
ของผปวย (Niumsup et al., 2009; Thapa et al., 2010; Jantama et al. 2013; Santimaleeworagun et al.,
2014) นอกจากนยงตรวจพบยนดอยา blaOXA-58 ในโรงพยาบาลทง 4 แหงในประเทศไทย คดเปนรอยละ 2.5 -7.3
ใกลเคยงกบการศกษาของ Jumroon และ Santanirand ในป ค.ศ. 2013 ทพบยนดอยา blaOXA-58 ภายในเชอ A.
baumannii ทแยกไดจากโรงพยาบาลรามาธบด จงหวดกรงเทพมหานคร คดเปนรอยละ 7.7 สวนยนดอยา blaOXA-24
ตรวจพบในเชอ A. baumannii ทแยกไดจากโรงพยาบาลสวรรคประชารกษเพยง 1 ไอโซเลท (1.5%) ตรงกนขามกบ
การศกษาของ Jumroon และ Santanirand ในป ค.ศ. 2013 ทตรวจพบยนดอยา blaOXA-24 รอยละ 7.7 ภายในเชอ A.
baumannii ทใหผลบวกกบการทดสอบทางฟโนไทปเพอตรวจสอบหาเอนไซม ในกลม metallo-β-lactamases
(MBLs) (Jumroon, & Santanirand, 2013) การศกษาครงนยงตรวจพบยน blaNDM-1 ในโรงพยาบาล 2 แหงในประเทศ
ไทยและในโรงพยาบาลจากประเทศเนปาล คดเปนรอยละ 8.2-18.8 ซงการพบยน blaNDM-1 พบครงแรกในประเทศไทย
ในป ค.ศ. 2012 จากตวอยางสงสงตรวจในเชอ Escherichia coli, Klebsiella pneumoniae และ Citrobacter
freundii ซงชใหเหนถงการแพรกระจายของยน blaNDM-1 ในโรงพยาบาลในประเทศไทย สวนยน blaVIM ตรวจพบในเชอ
A. baumannii ทแยกไดจากโรงพยาบาลจงหวดฉะเชงเทราเพยง 1 ไอโซเลท (0.8.5%) โดยยนนมรายงานการตรวจพบ
ในเชอ A. baumannii ทแยกจากโรงพยาบาลหวหน จงหวดประจวบครขนธ (Santimaleeworagun et al., 2016)
นอกจากนการศกษาครงนยงพบยนดอยา 2 ยน ไดแก blaOXA-23 และ blaOXA-58 อยรวมกนภายในไอโซเลตเดยวกน (co-
existence) จ านวน 2 ไอโซเลต เปนรายงานครงทสองในประเทศไทย โดยในครงแรกนนพบในเชอ CR-AB ทแยกไดจาก
ผปวยโรงพยาบาลพระมงกฎเกลา ในชวงเดอนมกราคม ค.ศ. 2012 ถงเดอนธนวาคม ค.ศ. 2013 จานวน 3 ไอโซเลต คด
เปนรอยละ 0.9 (Juntanawiwat et al., 2016) และเปนรายงานครงแรกทพบยนดอยา 3 ยนไดแก blaOXA-23, blaOXA-
58 and blaNDM-1 จ านวน 1 ไอโซเลตอยรวมกนภายในไอโซเลตเดยวกน (co-existence)

65
3. การสงผานยนดอยาโดยวธทรานสฟอรเมชนแบบธรรมชาต (Natural transformation)
การศกษาการสงผานยนดอยาโดยวธทรานสฟอรเมชน ใชพลาสมดจากเชอในกลม XDR-AB จ านวน 10 ไอโซเลต เปน
แบคทเรยผให และเชอ Acinetobacter spp. ทไวตอยา ตานจลชพ จ านวน 4 ไอโซเลต ไดแก NU005-1, NU013,
NU015 และ NI003 เปนแบคทเรยผรบ พบวาภายหลงจากการทดลองโดยวธทรานสฟอรเมชนแบบธรรมชาต (Natural
transformation) แลวน ามาท าการคดเลอก transformant บนจานอาหารเลยงเชอทมยาตานจลชพ ไดแก ticarcillin
(50 μg/ml), kanamycin (20 μg/ml) และ tetracycline (20 μg/ml) พบโคโลนเจรญไดเฉพาะบนอาหารเลยงเชอท
มยาตานจลชพ kanamycin (20 μg/ml) เมอน าโคโลนทไดไปท าการตรวจหายนดอยา aphA6 ซงเปนยนทท าใหเชอดอ
ตอยาตานจลชพ kanamycin พบวาตรวจไมพบยนดอยา aphA6 ใน transformants ทน ามาศกษา ซงยนทท าใหเชอ A.
baumannii ดอตอยาตานจลชพ kanamycin มอยดวยกนหลายยน เชน ยนในกลม phosphotransferases ไดแก
aphA1, aphA6 ยนในกลม acetyltransferases ไดแก aacC1, aacC2, aacA4 และยนในกลม
nucleotidyltransferases ไดแก aadB, aadA1 (Nemec et al., 2004) เพราะฉะนนเปนไปไดวาในเชอแบคทเรยผให
อาจมยนดอยาชนดอนทท าใหเชอดอตอยาตานจลชพ kanamycin เชนกน จงท าใหสามารถตรวจพบโคโลนเจรญบน
อาหารเลยงเชอดงกลาวได ในป ค.ศ. 2011 มรายงานการศกษาของ Rumbo et al. พบวาประสบความส าเรจในการ
สงผานยนดอยาจากแบคทเรยผใหไปสแบคทเรยผรบ โดยอาศยวธทรานสฟอรเมชนแบบธรรมชาต กลาวคอใช Outer
membrane vesicles (OMVs) ของเชอ A. baumannii สายพนธ AbH120-A2 และ AbH120-CU3 จ านวน 2 ไอโซ
เลต ซงมพลาสมดทมยนดอยา blaOXA-24 เปนแบคทเรยผให และเชอ A. baumannii ATCC17978 ซงไวตอยาตานจล
ชพในกลมคารบาพเนมเปนแบคทเรยผรบ จากการศกษาทดลองพบวาเชอ A. baumannii มการปลดปลอยของ Outer
membrane vesicles (OMVs) ซงภายในบรรจยนดอยา blaOXA-24 สามารถสงผานยนดอยานไปส A. baumannii
ATCC17978 ได สงผลใหเชอแบคทเรยผรบดอตอยาตานจลชพในกลมคารบาพเนม ไดแก imipenem, meropenem
และ doripenem (Rumbo et al., 2011) แตมการศกษาอนๆ รายงานวาการสงผานยนดอยาโดยวธทรานสฟอรเมชน
สวนใหญมกตองใชกระแสไฟฟาเขามาชวยในการกระตนใหแบคทเรยผรบพรอมทจะรบดเอนเอหรอพลาสมดทมยนดอ
ยาเขามาภายในเซลลใหมประสทธภาพดยงขน ทเรยกวา eletrotransformation เหนไดจากการศกษาของ Bertini และ
คณะทประสบความส าเรจในการสงผานยนดอยาจากพลาสมดของเชอ A. baumannii Ab120066 ทมยนดอยา blaOXA-

66
58 ไปสเชอ A. baumannii BM4547 ซงใชเปนแบคทเรยผรบ ท าใหเชอแบคทเรยผรบดอตอยาตานจลชพในกลมคาร
บาพเนมตามมา (Bertini et al., 2010) และยงมการศกษาในป ค.ศ. 2012 พบวาพลาสมดของเชอ Acinetobacter spp.
ทมยนดอยา blaNDM-1 สามารถสงผาน ยนดอยา blaNDM-1 ไปสเชอ A. baylyi ADP1 ไดโดยอาศยวธ
eletrotransformation และพบการสงผานยนดอยา blaOXA-58 จากพลาสมดในเชอ Acinetobacter spp. ไปสเชอ
แบคทเรยผรบ A. baumannii LS0148 ได สงผลใหตรวจพบยนดอยา blaOXA-58 และ การดอตอยาในกลมคารบาพเนม
เพมสงขน 2-32 เทา ใน transformants ทได (Fu et al., 2012; 2014) ผลการทดลองมาค านวณหาประสทธภาพของ
การทรานสฟอรเมชน (Transformation efficiency: TE) ในการศกษานพบวามคาอยในชวง 2.0 X 101- 8.0 x 102
transformants/μg plasmid DNA ซงแตกตางจากการศกษาของ Patwardhan et al. ทท าการศกษาการสงผานการ
ดอยาระหวาง พลาสมด pUPI281 ของเชอ A. baumannii A24 และเชอ E. coli HB101 โดยอาศยกระบวนการทรานส
ฟอรเมชนแบบใชสารเคมเหนยวน า (CaCl2) ซงใหคา TE เทากบ 4.3 x 104 transformants/μg plasmid DNA
(Patwardhan et al., 2008) เหนไดวาประสทธภาพของการทรานสฟอรเมชนในครงนต ากวาการศกษาของ
Patwardhan et al. เนองจากการศกษาของ Patwardhan et al. เปนการศกษาการสงผานยนดอยาโดยอาศย
กระบวนการ ทรานสฟอรเมชนแบบใชสารเคมเหนยวน า plasmid DNA เขาสแบคทเรยผรบ จงท าใหประสทธภาพใน
การทรานสฟอรเมชนดกวาการศกษาครงน จากนนน าผลการทดลองมาค านวณหาคาความถในการเกดกระบวนการท
รานสฟอรเมชน (Frequency of transformation) พบวามคา อยในชวง 1.1 X 10-10 ถง 4.4 X 10-11 ซงมคาต ากวา
การศกษาของ Doughari et al. ทท าการศกษาการสงผานการดอยาผานกระบวนการทรานสฟอรเมชนแบบธรรมชาต
โดยใชเชอ E. coli ทดอตอยาตานจลชพเปนแบคทเรยผให และใชเชอ A. lwoffii และ A. haemolyticus เปนแบคทเรย
ผรบจากการศกษาพบวาคา Frequency of transformation อยในชวง 1.3 - 5.4 X 10-6 (Doughari et al., 2012)
4. การสงผานยนดอยาโดยวธทรานสดกชน
การศกษาการสงผานยนดอยาโดยวธทรานสดกชนโดยอาศยแบคเทอรโอเฟจ เปนตวน าพายนดอยาจากแบคทเรยผให
ไปสแบคทเรยผรบ พบวาเชอแบคทเรยผให XDR-AB สายพนธ AB135 ทมแบคเทอรโอเฟจ ABP19 (ØABP-19) เปนตว
น าพายนดอยา สามารถน าพายนดอยา tet(B) ซงท าใหเชอดอตอยา tetracycline ไปสแบคทเรยผรบสายพนธ NU013
และ NU015 ได สงผลใหตรวจพบยนดอยา tet(B) ใน transductants ทได ดงแสดงใน ภาพ 7 กระบวนการทรานส

67
ดกชนนน สวนใหญมกเกดขนแบบทรานสดกชนทวไป (generalize transduction) เรมจากแบคเทอรโอเฟจทบรรจยน
ดอยาจากแบคทเรยผใหจะเขาไปเกาะตดกบเซลลแบคทเรยผรบ จากนนแบคเทอรโอเฟจจะท าการปลดปลอยยนดอยา
จากแบคทเรยผใหไปสแบคทเรยผรบ โดยยนดอยาจะแทรกเขาไปภายในโครโมโซมของแบคทเรยผรบ สงผลใหแบคทเรย
ผรบไดรบยนดอยาและดอตอยาตานจลชพตามมา ซงภายในเชอ A. baumannii ยงไมมผท าการศกษาทดลองเกยวกบ
การสงผานยนดอยาโดยวธทรานสดกชน แตมรายงานการศกษาอนๆในแบคทเรยแกรมลบ เชน ในเชอ E. coli โดย
Shousha et al. ซงท าการศกษาการสงผานยนดอยาโดยใชเชอ E. coli ทดอตอยาตานจลชพเปนแบคทเรยผให เชอ E.
coli ATCC 13706 เปนแบคทเรยผรบ และมแบคเทอรโอเฟจเปนตวน าพายนดอยา จากการศกษาพบวาประสบความสา
เรจในการสงผานยนดอยา aphA1, tet(A), floR และ blaTEM-1 จากแบคทเรยผใหไปสแบคทเรยผรบ (Shousha et al.,
2015) นอกจากนยงมการศกษาการสงผานยนดอยา โดยอาศยกระบวนการทรานสดกชนในแบคทเรยแกรมบวก โดยใช
เชอ S. aureus USA300 เปนแบคทเรยผให แบคเทอรโอเฟจ Ø80α, ØJB เปนตวนาพายนดอยา สวนเชอแบคทเรยผรบ
คอ S. aureus 07/235 และ 07/759 จากการศกษาพบวาแบคเทอรโอเฟจ Ø80α และ ØJB สามารถนาพายนดอยา
blaZ, cadD และ tet(K) ไปสแบคทเรยผรบได โดยมคา transduction frequency เทากบ 10-5-10-6 จะเหนไดวามคา
สงกวาการศกษาวจยในครงนทใช แบคเทอรโอเฟจของแบคทเรยแกรมลบ คอเชอ A. baumannii เปนตวน าพายนดอยา
(4.8 – 9.1 X 10-9) (Varga et al., 2012) จากทกลาวมาแสดงใหเหนวา แบคเทอรโอเฟจอาจเปนตวกลางส าคญทสามารถ
น าพายนดอยาไปโดยเฉพาะยนดอยากลม tetracycline ในเชอ A. baumannii ทไมดอตอยาตานจลชพ สงผลใหตรวจ
พบอตราการดอตอยาตานจลชพกลม tetracycline ภายในเชอ A. baumannii เพมสงขนในปจจบนอกดวย
5. การสงผานยนดอยาโดยวธคอนจเกชน
กระบวนการคอนจเกชน เปนกระบวนการสงผานยนดอยา โดยอาศยอวยวะพเศษ ทเรยกวา sex pilus ในการสงผานยน
ดอยาจากแบคทเรยผใหไปสแบคทเรยผรบ จากการศกษา ในครงนพบวาเชอ XDR-AB ทมยนดอยา kanamycin
(aphA6) สามารถสงผานยนดอยา aphA6 ซงท าใหเชอดอตอยาตานจลชพ kanamycin ไปสแบคทเรยผรบ A.
baumannii สายพนธ NU013 และ NU015 ไดโดยอาศยกระบวนการคอนจเกชน ท าใหสามารถตรวจพบยนดอยา
aphA6 ใน transconjugants เมอน ามาค านวณหาคา frequency of conjugation (CF) พบวามคาเทากบ 6.3 - 8.5 x
10-5 สวน transconjugants ทสามารถเจรญบนจานอาหารทมยาตานจลชพ ticarcillin 50 μg/ml เมอน ามาตรวจหา

68
ยนดอยาในกลม OXA-type carbapenemase (blaOXA-23, blaOXA-24, blaOXA-58 ) ตรวจไมพบยนดอยา สมพนธกบ
การศกษาของ Fu et al. ทรายงานวาการสงผานยนดอยาในกลม OXA-type carbapenemase (blaOXA-58) โดยอาศย
เชอ Acinetobacter spp. ไดแก A. pitti, A. nosocomialis และ A. baumannii เปนแบคทเรยผใหไปสเชอแบคทเรย
ผรบ A. baumannii ไมประสบความสาเรจในการสงผาน ยนดอยา (Fu et al., 2014) จงมความเปนไปไดวาอาจจะเปน
ยนดอยาในกลม β-lactam ชนดอน ทเกดการสงผานยนดอยาโดยวธคอนจเกชน สงผลใหเชอดอตอยาในกลมคารบาพ
เนมเชนกน จงท าใหตรวจพบโคโลน (transconjugants) เจรญบนจานอาหารเลยงเชอทมยาตานจลชพจงท าการตรวจหา
ยนอนเพมเตมไดแกยน blaPER-1 ซงเปนยนทสรางเอนไซม Class A beta-lactamase เปนเอนไซมทท าลายยาในกลม
penicillin และ cephalosporin สามารถพบยน blaPER-1 ในแบคทเรยผใหและใน transconjugant ดงนนการดอยา
ticarcillin นาจะเกดจากการสงผานยนดอยา blaPER-1 ในแบคทเรยผใหสงไปยงแบคทเรยผรบ
ปจจบนมการศกษารายงานวาการสงผานยนดอยาโดยอาศยกระบวนการคอนจเกชนสามารถสงผานยนดอยา blaOXA-23,
blaOXA-58, blaGES-11, blaIMP-1 และ blaNDM-1 จากแบคทเรยผให Acinetobacter spp. ไปสแบคทเรยผรบได สงผลให
ตรวจพบยนดอยาและอตราการดอยา ตานจลชพของแบคทเรยเพมสงขนอยางรวดเรว (Bertini et al., 2010; Charfi-
Kessis et al., 2014; Zhang et al., 2014; Yang et al. 2015) คา frequency of conjugation (CF) ทค านวณได
จากการศกษาทดลองในครงน อยในชวง 1.0 x 10-4 – 8.6 x 10-5 สมพนธกบการศกษากอนหนาในป ค.ศ. 2008 ท
ท าการศกษาการสงผานยนดอยาโดยอาศยกระบวนการคอนจเกชน ในการสงผานยนดอยา blaOXA-58 ระหวางเชอ A.
baumannii ทดอตอยาตานจลชพ imipenem ไปสเชอ A. baumannii AbF ทไวตอยาตานจลชพ imipenem พบวาม
คา CF อยท 2.0 x 10-5 – 1.5 X 10-6 (Zarrilli et al., 2008) อกทงยงสอดคลองกบการศกษาของ Talreja และคณะ ใน
ป ค.ศ. 2014ทท าการศกษาการสงผานยนดอยาจากเชอ MDR-AB ไปสเชอ E. coli J53 ซงใชเปนแบคทเรยผรบ จาก
การศกษาพบวาเชอ MDR-AB สามารถสงผานยนดอยา quinolone (aaC (6’)-lb) และ ยนดอยา tetracycline (tet(E),
tet(B)) ไปสเชอแบคทเรยผรบได โดยมคา CF เทากบ 10-4 (Talreja et al., 2014)

69
สรปและขอเสนอแนะ (Conclusion and Recommendation) สรปผลการวจย
การศกษาคณสมบตทางอณชววทยาของ class D β-lactamase และการถายทอดยน ในเชอ A. baumannii ทแยกได
จากตวอยางสงสงตรวจของผปวยในโรงพยาบาล 4 แหงในประเทศไทย และ โรงพยาบาล 1 แหงในประเทศเนปาลพบวา
สวนใหญมอตราการดอตอยาตานจลชพทใชในการทดสอบมากกวารอยละ 70 ในทกไอโซเลตไวตอยาตานจลชพ
colistin และ tigecycline เมอจดตามรปแบบการดอตอยาตานจลชพ พบวาจดอยในรปแบบ MDR-AB รอยละ 75-98
CR-AB รอยละ 65-98 และ XDR-AB รอยละ 2-21 จากนนท าการตรวจหายนดอยาในกลม OXA-type
carbapenemase พบวาตรวจพบยนดอยา blaOXA-23 มากทสด รองลงมาไดแก ยนดอยา blaOXA-58 ตามดวยยน
blaOXA-24 ตรวจพบเพยง 1 ไอโซเลต การตรวจหายนดอยาในกลม metallo beta-lactamase ตรวจพบ blaNDM-1 ใน
โรงพยาบาล 3แหงเปนโรงพยาบาลในประเทศไทย 2 แหงและเนปาล 1 แหง รอยละ 8-18 ตอมาศกษาการถายทอดยน
ดอยาในแนวราบ โดยอาศยกระบวนการทรานสฟอรเมชน ทรานสดกชน และ คอนจเกชน โดยใชเชอ XDR-AB เปน
แบคทเรยผให Acinetobacter spp. ซงแยกไดจากแหลงน าธรรมชาตในมหาวทยาลยนเรศวร และโรงพยาบาลพทธชน
ราช ทมคณสมบตไวตอยาตานจลชพทกชนดทใชในการทดสอบ และไมมยนดอยาเปนแบคทเรยผรบ จากการศกษา
พบวาเชอ XDR-AB สามารถน าพาการดอยา kanamycin, tetracycline และ ticarcillin ไปสแบคทเรยผรบได สามารถ
ตรวจพบยนดอยา tet(B) ซงท าใหเชอดอยา tetracycline ในแบคทเรยผรบของกระบวนการ ทรานสดกชน สามารถ
ตรวจพบยนดอยา blaPER-1 ซงท าใหเชอดอยา ticarcillin ในแบคทเรยผรบของกระบวนการคอนจเกชนและตรวจพบยน
ดอยา aphA6 ซงท าใหเชอดอยา kanamycin ในแบคทเรยผรบของกระบวนการคอนจเกชน
ขอเสนอแนะ
ผลทไดจากการศกษาทางอณชววทยาของ class D β-lactamase และการถายทอดยนในเชอ A. baumannii แสดงให
เหนวาปจจบนประเทศไทยพบอบตการณการดอตอยา ตานจลชพของเชอ A. baumannii เพมสงขนมาก สงผลใหการ
รกษาท าไดยาก ผปวยมอตราการเสยชวตสง อกทงยงพบยนดอยา โดยเฉพาะยนดอยา class D β-lactamase
แพรกระจายในวงกวางและเรมพบยนดอยาในกลม metallo beta-lactamase ไดแก blaNDM-1 เพมขน และยนดอยา
สามารถสงผานการดอยาหรอยนดอยาเหลานโดยอาศยกระบวนการถายทอดยนในแนวราบ ซงประกอบดวย 3
กระบวนการ ไดแก ทรานสฟอรเมชน ทรานสดกชน และคอนจเกชน ระหวางแบคทเรยดวยกน ซงในการศกษาครงนเปน

70
การศกษาขอมลเบองตนโดยไดจ าลอง การสงผานยนดอยาระหวางแบคทเรยผให A. baumannii ทแยกไดจากสงสง
ตรวจในโรงพยาบาลและแบคทเรยผรบ Acinetobacter spp. ทแยกไดจากสงแวดลอม เพราะฉะนนเพอใหเขาใจถง
กระบวนการถายทอดยนดอยาในแบคทเรยมากขน จงจ าเปนตองมการศกษาเพมเตม โดยใชเทคนค whole genome
sequencing เพอใหรสวนประกอบทงหมดภายในจโนมของแบคทเรยผใหและแบคทเรยผรบ แลวน าขอมลมาท านาย
การสงผานยนดอยาชนดอนๆทพบในจโนม ของแบคทเรย นอกจากนควรศกษาถงต าแหนงทพบยนดอยา โดยใชเทคนค
DNA-DNA hybridization หรอ southern blot และศกษากระบวนการสงผานยนดอยาอนๆเพมเตม เชน การสงผาน
ยนดอยาโดยใช outer membrane vesicle เพอใหเกดความเขาใจในการถายทอด ยนดอยามากยงขน รวมทงควรหาวธ
ทเหมาะสมในการก าจดแบคทเรยดอยาในโรงพยาบาล ไมใหมการปนเปอนออกมาสสงแวดลอม และน าผลการศกษาท
ไดไปประยกตใชในการควบคมปองกนการแพรกระจายของแบคทเรยดอยาทงภายในประเทศไทยและตางประเทศตอไป

71
เอกสารอางอง (reference)
1. Aimsaad, L., Diraphat, P., Utrarachkij, F., Thunyaharn, S., Samakoses, R., & Siripanichgon K.
(2009). Epidemiological characteristics of Acinetobacter baumannii infections at
Phramongkutklao Hospital. Journal of the Medical Association of Thailand, 92(7), 164-172.
2. Aminov, R. I., Chee-Sanford, J. C., Garrigues, N., Teferedegne, B., Krapac, I. J., White, B. A. &
Mackie, R. I. (2002). Development, validation, and application of PCR primers for detection of
tetracycline efflux genes of gram-negative bacteria. Applied and Environmental Microbiology,
68(4), 1786-1793.
3. Anunnatsiri, S. & Tonsawan, P. (2011). Risk factors and clinical outcomes of multidrug
resistant Acinetobacter baumannii bacteremia at a university hospital in Thailand. The
Southeast Asian Journal of Tropical Medicine and Public Health, 42(3), 693-703.
4. Aswapokee, N., Tiengrim, S., Charoensook, B. & Sangsiriwut, K. (1998). Antimicrobial resistant
pattern of Acinetobacter spp. Journal of Infectious Diseases and Antimicrobial Agents, 15(2),
43-48.
5. Bertini, A., Poirel, L., Mugnier, P. D., Villa, L., Nordmann, P., & Carattoli, A. (2010).
Characterization and PCR-Based Replicon Typing of Resistance Plasmids in Acinetobacter
baumannii. Antimicrobial Agents and Chemotherapy, 54(10), 4168-4177.
6. Bonnin, R.A., Poirel, L. Naas, T., Pirs, M., Seme, K., Schrenzel, J. & Nordmann. P. (2012).
Dissemination of New Delhi metallo-β-lactamase-1-producing Acinetobacter baumannii in
Europe. Clinical Microbiology and Infection, 18(9), E362-E365.
7. Chaiwarith, R., Mahatthanaphak, S., Boonchoo, M., Supparatpinyo, K. & Sirisanthana, T. (2005).
Pandrug-Resistant Acinetobacter baumannii at Maharaj Nakorn Chiang Mai Hospital. Journal
of Infectious Diseases and Antimicrobial Agents, 22(1), 347-352.

72
8. Chaisathaphol, T. & Chayakulkeeree, M. (2014). Epidemiology of infections caused by
multidrug-resistant gram-negative bacteria in adult hospitalized patients at Siriraj Hospital.
Journal of the Medical Association of Thailand, 97(Suppl 3), S35-S45.
9. Charfi-Kessis, K., Mansour, W., Ben Haj Khalifa, A., Mastouri, M., Nordmann, P., Aouni, M. &
Poirel, L. (2014). Multidrug-resistant Acinetobacter baumannii strains carrying the blaOXA-23
and the blaGES-11 genes in a neonatology center in Tunisia. Microbial Pathogenesis, 74, 20-
24.
10. Dejsirilert, S., Tiengrim, S., Sawanpanyalert, P., Aswapokee, N. & Malathum, K. (2012).
Antimicrobial resistance of Acinetobacter baumannii: six years of National Antimicrobial
Resistance Surveillance Thailand (NARST) surveillance. Journal of the Medical Association of
Thailand. 92(Suppl 4), S34-45.
11. Doughari, H. J., Ndakidemi, P. A., Human, I. S. & Benade, S. (2012). Virulence, resistance genes,
and transformation amongst environmental isolates of Escherichia coli and Acinetobacter
spp. Journal of Microbiology and Biotechnology, 22(1), 25-33.
12. Farshadzadeh, Z., Hashemi, F. B., Rahimi, S., Pourakbari, B., Esmaeili, D., Haghighi, M. A., &
Bahador, A. (2015). Wide Distribution of Carbapenem Resistant Acinetobacter baumannii in
Burns Patients in Iran. Frontiers in Microbiology, 6, 1-10.
13. Fu, Y., Du, X., Ji, J. Chen, Y., Jiang, Y. & Yu, Y. (2012). Epidemiological characteristics and
genetic structure of blaNDM-1 in non-baumannii Acinetobacter spp. in China. Antimicrobial
Chemotherapy Advance, 67(9), 2114-2122.
14. Higgins, P. G., Poirel, L., Lehmann, M., Nordmann, P. & Seiftert, H. (2009). OXA-143, a novel
carbapenem-hydrolyzing class D beta-lactamase in Acinetobacter baumannii. Antimicrobial
Agents and Chemotherapy, 53(12), 5035-5038.

73
15. Huang, X. Z., Frye, J. K., Chahine, M. A., Glenn, L. M., Ake, J. A., Su, W., Nikolich, M. P., &
Lesho, E. P. (2012). Characteristics of plasmids in multi-drug-resistant Enterobacteriaceae
isolated during Prospective Surveillance of a Newly Opened Hospital in Iraq. Plos one, 7(7),
e40360.
16. Huddleston, J. R. (2014). Horizontal gene transfer in the human gastrointestinal tract:
potential spread of antibiotic resistance genes. Journal of Infection and Drug Resistance, 7,
167-176.
17. Hujer, K. M., Hujer, A. M., Hulten, E. A., Bajaksouzian, S., Adams, J. M., Donskey, C. J. Ecker, D.
J., Massire, C., Eshoo, M. W., Sampath, R., Thomson, J. M., Rather, P. N., Craft, D. W., Fishbain,
J. T., Ewell, A. J., Jacobs, M. R., Paterson, D. L., & Bonomo, R. A. (2006). Analysis of antibiotic
resistance genes in multidrug-resistant Acinetobacter sp. isolates from military and civilian
patients treated at the Walter Reed Army Medical Center. Antimicrobial Agents and
Chemotherapy, 50(12), 4114-4123.
18. Inchai, J., Liwsrisakun, C., Theerakittikul, T., Chaiwarith, R., Khositsakulchai, W. & Pothirat, C.
(2015). Risk factor of multidrug-resistant, extensively drug-resistant and pandrug-resistant
Acinetobacter baumannii ventilator-associated pneumonia in a Medical Intensive Care Unit
of University Hospital in Thailand. Journal of Infection and Chemotherapy, 21(8), 570-574.
19. Jantama, S. S. , Kunawatanakul, T., Semaram, S., Sangsanon, S., Teerawattanasuk, N. &
Jantama. K. 2013. Detection of carbapenemase and blaOXA-23-like gene in carbapenem-
resistant Acinetobacter baumannii at Sunpasitthiprasong Hospital. I. J. P. S., 9, 99-109.
20. Juntanawiwat, P., Thunyaharn, S., Visawapoka, U., Samosornsuk, W. & Samosornsuk, S. (2016).
Prevalence of OXA–type carbapenemase genes in carbapenem-resistant Acinetobacter
baumannii isolates from patients in intensive care unit at Phramongkutklao Hospital. Journal
of Medical Technology and Physical Therapy, 28(2), 120-128.

74
21. Jumroon, N., & Santanirand, P. (2013). Multiple combination patterns of oxa-type
carbapenem hydrolyzing and metallo-β-lactamases encoding genes among clinically
isolated Acinetobacter baumannii. International Journal of pharma and bio sciences, 4,
908-917.
22. Joshi, P. R., Acharya, M., Kakshapati, T., Leungtongkam, U., Thummeepak, R., & Sitthisak, S.
(2017). Co-existence of blaOXA-23 and blaNDM-1 genes of Acinetobacter baumannii isolated
from Nepal: antimicrobial resistance and clinical significance. Antimicrobial resistance and
infection control, 6(21). 1-7.
23. Livermore, D. M. & Woodford, N. (2006). The β-lactamase threat in Enterobacteriaceae,
Pseudomonas and Acinetobacter. Trends in Microbiology, 14(9), 413-420.
24. Mendes, R. E., Bell, J. M., Turnidge, J. D., Castanheira, M. & Jones, R. N. (2009). Emergence and
widespread dissemination of OXA-23, -24/40 and -58 carbapenemases among Acinetobacter
spp. in Asia-Pacificnations: report from the SENTRY Surveillance Program. Journal of
Antimicrobial Chemotherapy, 63(1), 55–59.
25. Misbah, S., Hassan, H., Yusof, M. Y., Hanifah, Y. A. & AbuBakar, S. (2005). Genomic species
identification of Acinetobacter of clinical isolates by 16S rDNA sequencing. Singapore
Medical Journal, 46(9), 461-464.
26. Moghnieh, R., Siblani, L., Ghadban, D., El Mchad, H., Zeineddine, R., Abdallah, D., & El-Imad,
Z. (2016). Extensively drug-resistant Acinetobacter baumannii in a Lebanese intensive care
unit: risk factors for acquisition and determination of a colonization score. Journal of
Hospital Infection, 92(1), 47-53.
27. National Antimicrobial Resistance Surveillance Center, Thailand (NARST), Retrieved November
10, 2017, from http://narst.dmsc.moph.go.th/amrs.html

75
28. Nemec, A., Dolzani, L., Brisse, S., Broek. V. P. & Dijkshoorn, L. (2004). Diversity of
aminoglycoside-resistance genes and their association with class 1 integrons among strains of
pan-European Acinetobacter baumannii clones. Journal of Medical Microbiology, 53(12),
1233-1240.
29. Niumsup, P. R., Boonkerd, N., Tansawai, U. & Tiloklurs, M. (2009). Carbapenem-Resistant
Acinetobacter baumannii producing OXA-23 in Thailand. Japanese Journal of Infectious
Diseases, 62(2), 152-154.
30. Paton, R., Miles, R. S., Hood, J., Amyes, S. G., Miles, R. S. & Amyes, S. G. (1993). ARI 1: beta-
lactamase-mediated imipenem resistance in Acinetobacter baumannii. International Journal
of Antimicrobial Agents, 2(2), 81-87.
31. Patwardhan, R. B., Dhakephalkar, P. K., Niphadkar, K. B. & Chopade, B. A. (2008). A study on
nosocomial pathogens in ICU with special reference to multiresistant Acinetobacter
baumannii harbouring multiple plasmids. Indian Journal of Medical Research, 128(2), 178-
187.
32. Peleg, A.Y., Seifert, H. & Paterson, D. L. (2008). Acinetobacter baumannii: emergence of a
successful pathogen. Clinical Microbiology Reviews, 21(3), 538-582.
33. Poirel, L. & Nordmann, P. (2006). Carbapenem resistance in Acinetobacter baumannii:
mechanisms and Epidemiology. Clinical Microbiology and Infectious Diseases, 12(9), 826–836.
34. Poirel, L., Bonnin, R. A., & Nordmann, P. (2011). Genetic basis of antibiotic resistance in
pathogenic Acinetobacter species. International Union of Biochemistry and Molecular
Biology, 63(12), 1061-1067.
35. Ramirez, M. S., Merkier, A. K., Quiroga, M. P. & Centron, D. (2012). Acinetobacter baumannii is
able to gain and maintain a plasmid harbouring In35 found in Enterobacteriaceae isolates
from Argentina. Current Microbiology, 64(3), 211–213.

76
36. Robledo, I. E., Aquino, E. E., Santé, M. I., Santana, J. L., Otero, D. M., León, C. F, & Vázquez, G.
J. (2010). Detection of KPC in Acinetobacter spp. in Puerto Rico. Antimicrobial Agents and
Chemotherapy, 54(3), 1354-1357.
37. Rumbo, C., Ferna´ndez-Moreira, E., Merino, M., Poza, M., Mendez, J. A. & Soares, N. C.
Mosquera, A., Chaves, F., Bou, G. (2011). Horizontal transfer of the OXA-24 carbapenemase
gene via outer membrane vesicles: a new mechanism of dissemination of carbapenem
resistance genes in Acinetobacter baumannii. Antimicrobial Agents and Chemotherapy,
55(7), 3084–3090.
38. Santimaleeworagun, W., Thathong, A., Samret, W., Preechachuawong, P., Sae-lim, W. &
Jitwasinkul, T. (2014). Identification and characterization of carbapenemase genes in clinical
isolates of carbapenem-resistant Acinetobacter baumannii from general hospital in Thailand.
The Southeast Asian Journal of Tropical Medicine and Public Health, 45(4), 874-850.
39. Shousha, A., Awaiwanont, N., Sofka, D., Smulders, F. J., Paulsen, P., Szostak, M. P., Humphrey,
T., Hilbert, F. (2015). Bacteriophages isolated from chicken meat and the horizontal transfer
of antimicrobial resistance genes. Applied and Environmental Microbiology, 81(14), 4600-
4606.
40. Talreja, D., Muraleedharan, C., Gunathilaka, G., Zhang, Y., Kaye, K. S., Walia, S. K. & Kumar, A.
(2014). Virulence properties of multidrug resistant ocular isolates of Acinetobacter
baumannii. Current Eye Research, 39(7), 695-704.
41. Thapa, B., Tribuddharat, C., Srifuengfung, S. & Dhiraputra, C. (2010). High prevalence of
bla(OXA)-23 in oligoclonal carbapenem-resistant Acinetobacter baumannii from Siriraj
Hospital, Mahidol University, Bangkok, Thailand. The Southeast Asian Journal of Tropical
Medicine and Public Health, 41(3), 625-635.

77
42. Towner, K.J. (2009). Acinetobacter: an old friend, but a new enemy. Journal of Hospital
Infection, 73(4), 355-363.
43. Tu, Z. (2005). An improved system for competent cell preparation and high efficiency
plasmid transformation using different Escherichia coli strains. Electronic Journal of
Biotechnology, 8(1), 114-120.
44. Varga, M., Kuntová, L., Pantucek, R., Mašlanová, I., Ružicková, V. & Doškar, J. (2012). Efficient
transfer of antibiotic resistance plasmids by transduction with in methicillin-resistant
Staphylococcus aureus USA300 clone. FEMS Microbiology Letters, 332(2), 146-152.
45. Wang, J., Ruan, Z., Feng, Y., Fu, Y., Jiang, Y., Wang, H. & Yu, Y. (2014). Species distribution of
clinical Acinetobacter isolates revealed by different identification techniques. PLoS One, 9(8),
e104882.
46. Werarak, P., Waiwarawut, J., Tharavichitkul, P., Pothirat, C., Rungruanghiranya, S., Geater, S. L.,
Wiwatworapan, T. Thummakul T, Mootsikapun P, Rungsrithong N, Supawita S,
Chuchotthavorn C, Tongsai S, & Thamlikitkul V. (2012). Acinetobacter baumannii nosocomial
pneumonia in tertiary care hospitals in Thailand. Journal of the Medical Association of
Thailand, 95(Suppl 2), S23-33.
47. Woodford, N., Ellington, M. J., Coelho, J. M., Turton, J. F., Ward, M. E., Brown, S., Amyes, S. G.,
& Livermore, D.M. (2006). Multiplex PCR for genes encoding prevalent OXA carbapenemases
in Acinetobacter spp. International Journal of Antimicrobial Agents, 27(4), 351-353.
48. Wu, T. C. (2011). Carbapenem-resistant or multidrug-resistant Acinetobacter baumannii -a
clinician’s perspective. British Medical Bulletin, 16(4), 6-9.
49. Yang, Y. S., Lee, Y. T., Wang, Y. C., Chiu, C. H., Kuo, S. C., Sun, J. R., , Yin, T, Chen, TL, Lin, J. C.,
Fung, C. P., & Chang, F. Y. (2015). Molecular epidemiology of carbapenem non-susceptible

78
Acinetobacter nosocomialis in a medical center in Taiwan. Infection, Genetics and Evolution,
31, 305-311.
50. Zarrilli, R., Giannouli, M., Tomasone, F., Triassi, M. and Tsakris, A. (2009). Carbapenem
resistance in Acinetobacter baumannii: the molecular epidemic features of an emerging
problem in health care facilities. The Journal of Infection in Developing Countries, 3(5), 335-
341.
51. Zarrilli, R., Pournaras, S., Giannouli, M., & Tsakris, A. (2013). Global evolution of multidrug-
resistant Acinetobacter baumannii clonal lineages. International Journal of Antimicrobial
Agents, 41(1), 11-19.
52. Zhang, R., Hu, Y. Y., Yang, X. F., Gu, D. X., Zhou, H. W., Hu, Q. F., Zhao, K., Yu, S. F., & Chen, G.
X. (2014). Emergence of NDM-producing non-baumannii Acinetobacter spp. isolated from
China. European Journal of Clinical Microbiology & Infectious Diseases, 33(5), 853-860.

79
บทท 3
การศกษาทางอณชววทยาของเชอ extensively-drug
resistance Acinetobacter baumannii (XDR-AB)
Manuscript preparation
Phattaraporn Kongthai, Udomluk Leungtongkam, Rapee Thummeepak, Thawatchai kitti , Aunchalee
Thanwisai, Narisara Chantratita, Sutthirat Sitthisak. Molecular epidemiology of extensively-drug
resistance Acinetobacter baumannii (XDR-AB) isolated from four hospitals in Thailand.

80
บทน า (Introduction)
Extensively-drug resistance Acinetobacter baumannii หรอ XDR-AB คอ เชอ A. baumannii ทดอยา
ทกกลม ยกเวนยาในกลม tigecyclin และ colistin (Inchai, et al., 2015) เชอ XDR-AB เปนเชอทกอโรครนแรงและ
เปนปญหาตอการเลอกยาปฏชวนะทใชในการรกษาผปวย นอกจากน ยงพบวา XDR-AB มการระบาดมาตงแตในอดต
จนถงปจจบนและพบไดในหลายประเทศทวโลก ไดแก ประเทศไตหวน จน เลบานอน อหราน อหราน จน อยปตและ
ไทย เปนตน ซงพบความชกในการระบาดคอนขางสงในบางประเทศดงแสดงในตาราง 16 (Chan, et al., 2014; Li, et
al., 2015; Farshadzadeh, et al., 2015; Inchai, et al., 2014; Moghnieh, et al., 2016; Hasanin, et al., 2016)
แบคทเรย A. baumannii มอตราการดอยาทสงขน จงท าใหการรกษาโรคตดเชอไดยากมากขน ซงการดอยาของเชอ A.
baumannii เกดจากหลายกลไกและมเอนไซมเกยวของหลายชนดดงแสดงในตาราง 17 โดยกลไกทส าคญทสดทพบใน
เชอ A. baumannii คอการสรางเอนไซมบตาแลคทาเมส ซงสามารถแบงไดหลายกลม โดยจดกลมตามโครงสราง
โมเลกล (molecule classification หรอ Ambler classification) ได 4 กลม (class) ไดแก class A, B, C และ D
ตาราง 16 ความชกของการระบาดของ XDR-AB และยนดอยาทพบ
ประเทศ ยนทตรวจพบ ความชก เอกสารอางอง Thai NT 65.3% Inchai, et al., 2015 Lebanon NT 15.6% Moghnieh, et al.,
2016 Iran blaOXA-23, blaPER-1, ISAba1, Class1integron 77% Farshadzadeh, et
al., 2015 Egypt NT 63.8% Hasanin, et al.,
2016 China blaOXA-51 67.31% Li, et al., 2017 Nepal Thailand
blaOXA-23, blaNDM-1, ISAba, ISAba125, blaADC, aphA6 blaOXA-23, blaOXA-24, blaOXA-58, blaNDM-1
2.33% 7.9%
Joshi, et al., 2017 Leungtongkam, et al., 2017
NT = not tested

81
ตาราง 17 กลไกการดอยาของ A.baumannii ตอยาแตละกลม
เอนไซมในกลมบตาแลคทาเมส (Beta-lactamase)
เอนไซมบตาแลคทาเมส สามารถแบงไดหลายกลม โดยจดกลมตามโครงสรางโมเลกล (molecule
classification หรอ Ambler classification) ได 4 กลม (class) ไดแก class A, B, C และ D และสามารถแบงเปนกลม
ใหญได 2 กลม คอ Serine beta-lactamase และ Metallo-beta-lactamase โดย Serine beta-lactamase เปน
เอนไซมบตาแลคทาเมส ทมต าแหนงการเกดปฏกรยา (active site) อยบรเวณกรดอะมโน serine ของโมเลกล สวน
Metallo beta-lactamase จะเกดปฏกรยา กตอเมอมโมเลกลของโลหะไอออน คอ Zn2+ มารวมในปฏกรยา จงจะ
Drug class Resistance mechanism Examples
β-lactam - Inactivating enzymes - Decreased outer membrane protein express - Alter penicillin-binding protein expression - Efflux pump
β-lactamase (AmpC, TEM, VEB, PER, CTX-M, SHV) Carbapenemase (OXA-23, -40,-51, VIM, NDM) CarO, OprD-like protein PBP-2 AdeAB
Fluoroquinolones -Target modification -Efflux pump
Mutations in gyrA and parC AdeABC, AdeM
Aminoglycoside - Aminoglycoside modification enzymes - Efflux pump - Ribosomal methylation
AAC, ANT, APH AdeABC, AdeM ArmA
Tetracycline -Efflux pump -Ribosomal protection
AdeABC, TetA,TetB tetM
Glycylcycline Polymixins(colistin) -Target modification Mutation in the PmrA/Btwo component
system(LPS modification), mutation in LPS biosynthesis gene

82
สามารถออกฤทธท าลายยาได โดยเอนไซม serine beta-lactamase ประกอบดวย 3 class คอ class A, class C และ
class D สวน metallo beta-lactamase ไดแก class B (Dahiya, et al., 2015)
Class A beta-lactamase เปนเอนไซมทท าลายยาในกลม penicillin และ cephalosporin ไดมากกวาการ
ท าลายยากลม carbapenem นอกจากนยงสามารถท าลายยาในกลมmonobactam ได เอนไซมในกลมนถกยบยงดวย
กรด clavulanic acid ตวอยางเอนไซมในกลมน ไดแก TEM, SHV, GES, CTX-M, SCO, PER, VEB, KPC และ CARB
เปนตน (ภทรชย และอนชา, 2555; Dahiya, et al., 2015; Lee, et al., 2017) เอนไซมในกลมนทสามารถพบไดใน
A. baumannii ไดแก PER-1 (Hojabri, et al., 2014, Farshadzadeh, et al., 2015), CTX-M (Alyamani, et al.,
2015), SHV (Zhao, et al., 2015) และ GES (Bogaerts, et al., 2010)
Class B beta-lactamase หรอ metallo beta-lactamase เปนเอนไซมทตองอาศยโมเลกลของโลหะไอออน
คอ Zn2+ มารวมในปฏกรยา จงจะสามารถท างานได และสามารถถกยบยงไดดวย EDTA เชอทมการสรางเอนไซมกลมน
สงผลใหดอตอยาในกลม penicillin, cephalosporins และ carbapenem เอนไซมในกลมน ไดแก IMP, NDM, VIM,
GIM เปนตน (Lee, et al., 2017) เอนไซมในกลมนทสามารถพบไดใน A. baumannii ไดแก VIM (Zhao et al., 2015;
Gomaa et al., 2017), NDM (Gomaa, et al., 2017)
Class C beta-lactemase ไดแก AmpC หรอ cephalosporinase เปนเอนไซมทสามารถท าลายยาในกลม
cephalosporin ไดดเนองจากมความจ าเพาะตอยากลมน พบยนควบคมการสรางเอนไซมนอยบนโครโมโซม แตใน
ปจจบนพบวามยนทสรางเอนไซมนอยบนพลาสมดดวย โดยในสภาวะปกตยนทควบคมการสรางเอนไซม AmpC มการ
แสดงออกในระดบต า (repression) แตถาเชอจะสรางเอนไซมนกตอเมอเจรญในสภาวะทมยา ampicillin, imipenem
รวมถง clavulanic acid ยนจงมการแสดงออกในภาวะกระตนหรอ inducible expression ท าใหสรางเอนไซมออกมา
ในปรมาณมากกวาปกตและดอตอยาในกลม cephalosporin ได (ภทรชย และอนชา, 2555; Dahiya, et al., 2015)
เอนไซมในกลมน ไดแก MOX, CIT, ACC, ADC, EBC, FOX และ DHA เอนไซมในกลมนทสามารถพบไดใน A.
baumannii ไดแก ADC ( Liu Y and Liu X., 2015)
Class D beta-lactamase หรอ oxacillinases ไดแก เอนไซมในวงศ OXA เอนไซมในกลมนสามารถท าลาย
ยา oxacillin ได และท าใหเชอดอตอยาในกลม carbapenem การท างานของเอนไซมถกยบยงการท างานดวย
โซเดยมคลอไรด โดยพบยนทสรางเอนไซมมต าแหนงอยบนโครโมโซม และหนวยพนธกรรมเคลอนท ไดแก class 1

83
integron บนพลาสมด สมาชกของเอนไซมในวงศ OXA มความหลากหลายของโครงสรางโมเลกลคอนขางมาก เอนไซม
ในวงศ OXAs มหลายกลมยอย ไดแก OXA-2, OXA-10, OXA-20, OXA-23, OXA-24, OXA-51, OXA-58, OXA-143
และ OXA-235 ซงเอนไซม OXA-51 สามารถพบไดใน A. baumannii ทกตว เนองจากเปนยนทอยบนโครโมโซมของ
เชอน (intrinsic gene) (ภทรชย และอนชา, 2555; Ahmed & Alp, 2015; Lee, et al., 2017) นอกจากนเอนไซมใน
กลมนทพบไดมากยงม OXA-23 (Mammina., 2012; Zarrilli, et al., 2013), OXA-24 (Hojabri., 2014; Zarrilli, et
al., 2013) และ OXA 58 (Mammina., 2012; Zarrilli, et al., 2013) เปนตน
การจดกลมแบคทเรย A. baumannii โดยเทคนคทางอณชววทยา (molecular technique)
แบคทเรย A. baumannii เปนเชอทมอบตการณการดอยาสง เนองจากยนดอยาในตวเชอสามารถถายทอดส
แบคทเรยสายพนธเดยวกนหรอชนดอนได เปนเหตใหมเชอดอยาแพรกระจายอยางทวถงภายในโรงพยาบาลเดยวกน
หรออาจกระจายไปยงตางโรงพยาบาลหรอพนทอนๆได การสบหาตนตอหรอแหลงการแพรกระจายเชอ สามารถท าได
โดยการจดกลมของเชอทแยกไดจากผปวย หากเชอทแยกจากผปวยทอยคนละแหงกน แตพบวา เชอทแยกได เปนเชอ
สายพนธเดยวกน อาจระบไดวา มการแพรกระจายของเชอมาจากทได โดยดจากขอมลการพบเชอกอนหนา ซงการจด
กลมแบคทเรยจงถอเปนการวเคราะหขอมลทางระบาดวทยา (epidimiology) ของเชอไดเทคนคทใชในการจดกลมของ
แบคทเรย A. baumannii ไดแก
1. Multilocus sequence typing (MLST)
เปนเทคนคการจดกลมของเชอ โดยการจดกลมทไดจะออกมาในรปแบบของ sequence type (STs) ท าการ
เปรยบเทยบความเหมอนของ house keeping genes จ านวน 7 ยน (ตาราง 18) ของแตละเชอทน ามาจดกลม โดย
เปรยบเทยบแตละยนของเชอ ถายนใดยนหนงของเชอทน ามาเปรยบเทยบมความเหมอนกน เชอเหลานนจะถกจดอยใน
ST เดยวกน โปรแกรมจะแสดงเปนตวเลข ST เดยวกน (ภาพ 10) แตถาเชอทน ามาเปรยบเทยบกน มบางเชอทมล าดบ
เบสของยนเดยวกนทแตกตางกน กจะถกจดอยอก ST ทตางกน เลข ST กตางกนไปดวย เมอประมวลผลรวมทง 7 ยน
ของเชอทกตว โปรแกรมจะแสดงผล การจดกลมของเชอได เทคนคนมการใชโปรแกรมเขามาวเคราะหขอมล ซง
โปรแกรมทนยมใช คอ PubMLST (http://pubmlst.org/abaumannii/) จากการวเคราะหขอมลแสดงดงภาพ 10 จะ
เหนวายนแรกเปรยบเทยบกนทง 3 สายพนธ คอ ยน abcZ พบวา strain A และ C มล าดบเบสของยนนเหมอนกนจงได

84
รหสยนเปนเลข 1 เหมอนกน แต strain B มล าดบเบสของยนนแตกตางจาก strain A และ C จงไดรหสยนเปนเลข 2
โปแกรมจะท าการวเคราะห 7 ยนของแตละเชอแลวแสดงผลออกมาเปน ST
ภาพ 10 ตวอยางแสดงการเปรยบเทยบ STs ระหวางเชอสายพนธ A B และ C
ทมา: http://www.applied-maths.com/applications/mlst
ตาราง 18 House keeping gene ของเชอ A. baumannii ทใชในรปแบบ Oxford scheme (Bartual, et
al., 2005) และ Pasteur scheme (http://pubmlst.org/abaumannii/)
Oxford scheme Pasteur scheme
gltA cpn60 gyrB fusA gdhB gltA recA pyrG
cpn_60 recA gpi rplB
rpoD rpoB
Bartual และคณะ (ค.ศ. 2005) ท าการพฒนารปแบบของการท า multilocus sequence typing (MLST) ใน
แบคทเรย A. baumannii โดยท าการน าเชอ A. baumannii ทแยกไดในป 1990-2002 มาท าการสกด DNA และท า
การจดกลมเพอดรปแบบการระบาดดวยวธทางอณชววทยาหลายวธ ไดแก MLST, PFGE ซงพบวาการจดกลมการ
ระบาดของเชอโดยใชวธ MLST สามารถจดกลมเชอเปน sequence type (STs) ได 21 STs ไดแก ST1, ST2, ST3,

85
ST5, ST6, ST7, ST8, ST9, ST10, ST11, ST12, ST13, ST14, ST15, ST16, ST17, ST18, ST19, ST20 แ ล ะ ST21
ซงรปแบบ house keeping gene ทใชเปนแบบ Oxford scheme (Bartual, et al., 2005)
Mammina และคณะ (ค .ศ . 2012) ท าการศ กษาการระบาดของ เช อ carbapenem resistance A.
baumannii (CR-AB) ทแยกไดจากแผนกผปวยหนก ในเมอง Palermo ประเทศอตาล โดยวธทใชในการดรปแบบการ
ระบาดของเชอ คอ วธ MLST และ REP-PCR พบวาผลจากการจดกลมการระบาดโดยวธ MLST สามารถจดกลมได 2
STs คอ ST2 และ ST78 ซงรปแบบ house keeping gene ทใชเปนเปน Pastuer scheme ซง ST78 เรยกวา Italian
clone เนองจาก การระบาดของ ST78 พบมากทสดในประเทศอตาล (Mammina, et al., 2012)
Jiang และคณะ (ค.ศ.2016) ท าการศกษาการระบาดของเชอ MDR-AB ทแยกไดจากตวอยางของผปวย ไดแก
เสมหะ หนอง แผล ปสสาวะ เปนตน ในแผนกผปวยวกฤต (ICU) ของโรงพยาบาล 10 แหงในประเทศจน โดย
ท าการศกษาเชอ A. baumannii ทแยกไดและจดกลมเพอดรปแบบการระบาดโดยใชวธ MLST พบวาผลจากการจด
กลมการระบาดโดยวธ MLST สามารถจดกลมได 12 STs ไดแก ST369, ST 208, ST195, ST191, ST368, ST540,
ST218, ST469, ST451, ST809, ST794 และ ST381 จากผลการจดกลม มตวอยางผปวยจากโรงพยาบาลคนละแหง
แตมถกจดอยใน ST เดยวกน แสดงวาอาจมการแพรกระจายของเชอระหวางโรงพยาบาลเกดขน (Jiang, et al., 2016)
Seruga และคณะ (ค .ศ .2017) ท าการศ กษาการระบาดของ เช อ extensively-drug resistance A.
baumannii (XDR-AB) ทแยกไดจากผปวยโรคปอดอกเสบทพกรกษาในโรงพยาบาล น าเสยทปลอยออกจาก
โรงพยาบาล แมน าบรเวณใกลเคยงกบโรงพยาบาล ในประเทศ Croatia โดยวธทใชในการดรปแบบการระบาดของเชอ
คอ วธ MLST ซงสามารถจดกลมได 2STs คอ ST195 และ ST1421 พบวา เชอทแยกไดจากผปวยโรคปอดอกเสบ เชอ
ทแยกไดจากน าเสยบรเวณโรงพยาบาลและเชอทแยกไดจากแมน าบรเวณใกลกบโรงพยาบาล มบางเชอทถกจดกลมเปน
ST เดยวกน แสดงใหเหนวามการกระจายเชอระหวางผปวยโรคปอดอกเสบทพกฟนในโรงพยาบาลไปสสงแวดลอม คอ
น าเสยบรเวณโรงพยาบาลและแมน าบรเวณใกลกบโรงพยาบาล (Seruga, et al., 2017)
2. blaOXA-51 Sequence base typing (blaOXA-51 SBT)
เปนการท า PCR เพอเพมปรมาณยนทสนใจเพยงยนเดยว คอ blaOXA-51 ซงเปนยนทพบไดใน A. baumannii ทกตว
(Ahmed and Alp, 2015) จากนนท าการ sequencing ล าดบนวคลโอไทดของยน blaOXA-51 และแปลงล าดบนวคล
โอทเปนล าดบกรดอะมโนของโปรตน OXA-51 จากนนท าการจดกลมเชอ โดยเปรยบเทยบความเหมอนของล าดบ

86
กรดอะมโนกบฐานขอมล หากมล าดบกรดอะมโนของโปรตน OXA-51 เหมอนกน โปรแกรมจะแสดงผลใหเหนวาเปน
subgroup ของโปรตน OXA-51 กลมเดยวกน แตถาเชอทน ามาเปรยบเทยบมล าดบกรดอะมโนของโปรตน OXA-51 ท
แตกตางกน โปรแกรมจะแสดงผลใหเหนวาเชอทน ามาเปรยบเทยบกนนนเปน subgroup คนละกลม
Evans และคณะ (ค.ศ. 2008) ท าการศกษาการระบาดของเชอ A. baumannii โดยใชวธ blaOXA-51 SBT ผล
การศกษาพบการระบาดของ subgroup ของโปรตน OXA-51 ไดหลากหลาย ไดแก OXA-69, -66, -71, -64, -65, -68,
-82, -109, -108, -83, -112, -106, -107, -110, -111 และ OXA-98 ซง subgroup ของโปรตน OXA-51 ทพบไดมาก
ทสด คอ OXA-66 รองลงมา คอ OXA-69 และ OXA-71 ตามล าดบ (Evans, et al., 2008)
Pournaras และคณะ (ค.ศ. 2014) ท าการศกษาการระบาดของเชอ A. baumannii โดยใชวธ blaOXA-51 SBT
ซงแยกเชอไดจากหลายประเทศ ไดแก อตาล กรซ เลบานอน ตลก อเมรกาและเยอรมน ผลการศกษาพบการระบาดของ
subgroup ของโปรตน OXA-51 ไดหลายกลม ไดแก OXA-66, -69, -71, -64, -90, -94, -365, -51, -128, -86, -100
และ OXA-65 เปนตน (Pournaras, et al., 2014)
ตอมาในป ค.ศ. 2015 Rafei และคณะ ท าการศกษาการระบาดของเชอ A. baumannii โดยใชวธ blaOXA-51
SBT ซงแยกเชอไดจากโรงพยาบาล ในเมอง Tripoli ประเทศเลบานอน ผลการศกษาพบการระบาดของ subgroup
ของโปรตน OXA-51 ไดหลายกลมและเรยงจากปรมาณการตรวจพบตามล าดบ ไดแก OXA-66 (พบมากทสด), -69, -
64, -94, -120, -70, -71, -104, -121, -132 และ OXA-406 (เปนโปรตนชนดใหมทพบครงแรกของ subgroup ของ
โปรตน OXA-51) เปนตน (Rafei, et al., 2015)
3. Repetitive-PCR (REP-PCR)
วธ REP-PCR เปนการท า PCR โดยเพมปรมาณยน ซงไพรเมอรจะไปจบบรเวณทมเบสซ าๆกน จากนน
เปรยบเทยบรปแบบลายพมพดเอนเอ (DNA fingerprint) โดยแบคทเรยสายพนธเดยวกน จะมความเหมอนกนของชวง
ล าดบเบสซ า (repeat sequence) บรเวณต าแหนง non-coding region (Ben-Darif, et al., 2010) จงสามารถ
น ามาใชในการเปรยบเทยบและจดกลมเพอดรปแบบการระบาดของตวอยางแบคทเรยทสนใจได ในป ค.ศ. 2000 Bou
และคณะ ท าการศกษาการระบาดของเชอ A. baumannii ทดอตอยา imipenem และ meropenem ซงแยกไดจาก
ผปวยทตดเชอเนองจากนอนพกรกษาในโรงพยาบาลเปนเวลานาน (nosocomial infection) และท าการจดกลมเชอ
เพอดรปแบบการระบาดดวยวธ REP-PCR พบวาสามารถจดกลมรปแบบการระบาดไดเปน 8 กลม ไดแก กลมท 1, 2, 3,

87
4, 5, 6, 7 และ 8 จากขอมล พบวา ในการระบาดทถกจดกลมในรปแบบท 1 มการระบาดระหวางแผนกการรกษาใน
โรงพยาบาล ไดแก แผนกผปวยหนกทวไป แผนกผปวยหนกทมการผาตด แผนกผปวยผวหนงเนองจากถกจดอยในกลม
รปแบบการระบาดเดยวกน คอ รปแบบการระบาดกลมท 1 (Bou, et al., 2000) ตอมาในป ค .ศ. 2014
Santimaleeworagun และคณะ ท าการศกษาเชอ A. baumannii ทดอยาในกลม carbapenem ซงเปนเชอทแยกได
จากผปวยทนอนพกรกษาในโรงพยาบาล ในประเทศไทย และจากการศกษาสามารถจดกลมรปแบบการระบาดได 4
ก ล ม ค อ clone A, B, C แ ล ะ D ซ ง จ า ก เ ช อ ท แ ย ก ไ ด พบ เ ป น ก า ร ระ บ าด ร ป แบบ clone A ม า กท ส ด
(Santimaleeworagun, et al., 2014)
จากการศกษาทผานมาโดยอดมลกษณ เหลองทองค าและคณะ (Leungtongkam et al., 2017) ไดศกษา A.
baumannii จ านวน 339 ไอโซเลท จากผปวยทมารกษาในโรงพยาบาล 4 แหง ในประเทศไทย ซงเปนโรงพยาบาล
ประจ าจงหวด พบ XDR-AB 27 ไอโซเลท คดเปนรอยละ 7.96 แตยงไมมการจดกลมศกษาถงรปแบบการระบาดของ
XDR-AB ทแยกได รวมถงยงไมมการศกษาเกยวกบยนดอยาในกลมบตาแลคทาเมส (Beta-lactamase) ครบทง 4 class
การศกษาในครงนจงเปนการศกษาเพอจดกลมเชอ XDR-AB และประเมนทางดานระบาดวทยาโมเลกล ซงขอมลทได
สามารถเปนตวบงชถงตนตอการระบาดและเปนขอมลทเพอเปนแนวทางก าหนดมาตรการในการลดความเสยงของการ
แพรกระจายเชอเพมมากขน

88
วธด าเนนการวจย (Material and Method)
1. แบคทเรยทใชในการศกษาวจย
แบคทเรยทใชในการศกษาวจยเปนแบคทเรยทเกบจากโรงพยาบาลประจ าจงหวด 4 แหงในประเทศไทย ไดแก
โรงพยาบาลในจงหวดเชยงราย พจตร นครสวรรคและจงหวดฉะเชงเทรา น าเชอทแยกไดมาท าการระบชนดของเชอวา
เปน Acinetobacter baumannnii โดยท าการทดสอบ ทางชวเคมและท าการสง sequence เพอระบชนดของเชอ โดย
ดล าดบเบสและเปรยบเทยบล าดบเบสกบฐานขอมล NCBI ดวยการท า BLASTN และท าการทดสอบรปแบบการ
ดอตอยา 14 ชนด โดยวธ disk diffusion พบวาดอยา 12 ชนด ไดแก amikacin (AK), ciprofloxacin (CIP),
trimethoprim- sulfamethoxazole (SXT), cefotaxime (CTX), ceftazidime (CAZ), imipenem (IMP),
meropenem (MEM), piperacillin-tazobactam (PTZ), tetracycine (TE), ceftriazone (CRO),
cefepime (FEP), gentamicin (CN) โดยขอมลการดอยา ทไดมการรวบรวมจากการศกษามากอนหนาน
(Leungtongkam, et al., 2017) เชอทแยกไดทงหมดเกบใน glycerol stock ท -40 ˚C
2. การตรวจหายนดอยาในเชอ XDR-AB โดยวธ PCR
ตรวจหายนดอยาในเชอ XDR-AB โดยท า PCR เพอตรวจหายนดอยาในกลมบตาแลคทาเมส (Beta-lactamase) ทง 4
class (Class A, B, C, D) จ านวน 17 genes จากเชอ XDR-AB ทง 27 ไอโซเลท โดยใช primer ตามตาราง 19 โดย
อางองวธท าตามรายงานทมมากอนหนา (Woodford, et al., 2006; Kalai, et al., 2007; Lee et al., 2008; Kolar, et
al.,2010; Poirel, et al., 2011; Hout, et al., 2015; Liu Y& Liu X., 2015; เมอท า PCR เสรจแลว น า PCR product
ทไดมาท าการตรวจหา DNA ดวยวธ วธอะกาโรสเจลอเลคโตรโฟรสส เพอหาขนาดของ PCR product โดยเทยบกบด
เอนเอมาตรฐาน 100 bp (solis bioline) โดยใช 1% เจลอะกาโรสทมเอทเดยมโบรไมด ใชความตางศกย 100
โวลต เปนเวลา 30 นาท จากนนตรวจดผลภายใตแสง ยว

89
ตาราง 19 ขอมล primer ของ drug-resistance gene
Class of
β-lactamase
genes Primer sequence
(5’3’)
Product
size
(bp)
Tm
(˚C)
references
A blaSHV CTTTACTCGCTTTATCG TCCCGCAGATAAATCAC
635 50 Kolar, et al., 2010
A blaTEM ATGAGTATTCAACATTTCCG CCAATGCTTAATCAGTGAGC
569 54 Kalai, et al., 2007
A A A
blaCTX-M
blaPER-1
blaGES
ATGTGCAGYACCAGTAARGTKATGGC TGGGTRAARTARGTSACCAGAAYCAGC ATGAATGTCATTATAAAAGC AATTTGGGCTTAGGGCAAGAAA ATGCGCTTCATTCACGCAC CTATTTGTCCGTGCTCAGG
593 900 864
54 52 52
Hout, et al., 2015 Lee et al., 2008 Poirel, et al.,2000
B blaSPM AAAATCTGGGTACGCAAACG ACATTATCCGCTGGAACAGG
271 56 Poirel, et al.,2011
B blaSIM TACAAGGGATTCGGCATCG TAATGGCCTGTTCCCATGTG
570 58 Poirel, et al.,2011
B blaNDM-1 GGTTTGGCGATCTGGTTTTC CGGAATGGCTCATCACGATC
621 52 Poirel, et al.,2011
C blaACC AACAGCCTCAGCAGCCGGTTA TTCGCCGCAATCATCCCTAGC
346 56 Liu Y& Liu., 2015
C blaADC TAAACACCACATATGTTCCG ACTTACTTCAACTCGCGACG
633 56 Liu Y & Liu., 2015
C blaMOX GCTGCTCAAGGAGCACAGGAT CACATTGACATAGGTGTGGTGC
520 56 Liu Y& Liu., 2015
C blaCIT TGGCCAGAACTGACAGGCAAA TTTCTCCTGAACGTGGCTGGC
462 56 Liu Y& Liu., 2015
C blaDHA AACTTTCACAGGTGTGCTGGGT CCGTACGCTTACTGGCTTTGC
405 56 Liu Y& Liu., 2015
C blaEBC TCGGTA AAGCCGATGTTGCGG CTTCCACTGCGGCTGCCAGTT
302 56 Liu Y& Liu., 2015

90
Class of
β-lactamase
genes Primer sequence
(5’3’)
Product
size
(bp)
Tm
(˚C)
references
D blaOXA-23 GATCGGATTGGAGAACCAGA ATTTCTGACCGCATTTCCAT
501 52 Woodford, et al., 2006
D blaOXA-24 GGTTAGTTGGCCCCCTTAAA AGTTGAGCGAAAAGGGGATT
246 52 Woodford, et al., 2006
D blaOXA-51 TAATGCTTTGATCGGCCTTG TGGATTGCACTTCATCTTGG
353 52 Woodford, et al., 2006
D blaOXA-58 AAGTATTGGGGCTTGTGCTG CCCCTCTGCGCTCTACATAC
599 52 Woodford, et al., 2006
3. การจดกลม XDR-AB โดยใชวธ blaOXA-51 sequence base typing
น าเชอ XDR-AB ทง 27 ไอโซเลท มาท า PCR เพอตรวจหายน blaOXA-51 โดยใช primer OXA-69 (CTAATAATTGATC
TACTCAAG) และ OXA-69B (CCAGTGGATGGATGGATAGATTATC) และมสภาวะในการท า PCR ดงน ขนท 1 95
องศาเซลเซยส 2 นาท ขนท 2 95 องศาเซลเซยส 1 นาท, 52 องศาเซลเซยส 30 วนาท, 72 องศาเซลเซยส 1 นาท ขนท
3 72 องศาเซลเซยส 2 นาท เมอพบ PCR product ของไอโซเลททใหผลบวก น า PCR product
ของเชอไอโซเลทมาท าการ purified โดยใชชด kit GF-1 AMBICLEAN KIT PCR & Gel (Vivantis) กอนทจะท าการสง
PCR product ใหกบบรษท Macrogen ประเทศเกาหล เพอวเคราะหล าดบเบส (DNA sequences) ท าการวเคราะห
ล าดบเบส DNA โดยใชโปรแกรม Seqman จากนนเปลยน DNA sequence ใหเปน amino acid sequence
โดยใชโปรแกรม ORF finder (http://www.bioinformatics.org/sms2/orf_find.html) และน า amino acid sequ
ence วเคราะหในโปรแกรม Blast ใน NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi.PAGE=Proteins) เพอด s
ubgroup (% similarlity) ของยน blaOXA-51จากนนท าการจดกลมและท า phylogenetic tree โดยใชโปรแกรม M
EGA

91
2. การจดกลม XDR-AB โดยใชวธ Repetitive Element Palindromic-PCR (REP-PCR)
น าเชอ XDR-AB ทง 27 ไอโซเลททเพาะเลยงไวขามคนมาท าการสกด Genomic DNA โดยใช GF-1 bacterial DNA
extraction kit (Vivantis, Malaysia) จากนนน า Genomic DNA มาใชเปน template ในการเพมจ านวนในเครอง PCR
โดยใช ERIC-2 primer และสภาวะการท า PCR อางองตามวธของ Reboli และคณะ (1994) D’Arezzo และคณะ
(2009) การแปลผล PCR-banding patterns และ rep-PCR types อางองตามวธของ D’Arezzo และคณะ (2009)
3. การจดกลม XDR-AB โดยวธ MLST
ท าการสกด genomic DNA ของเชอ โดยใชชด kit GF-1 BACTERIAL DNA EXTRACTION (vivantis) จากนนท า PCR
ของ house keeping gene จ านวน 7 ยน ของเชอ XDR-AB ทง 27 ไอโซเลท โดยใช primer ของ house keeping
gene 7 ยนตามตาราง 20 จากนนน า PCR product ทไดมาท าการ purified โดยใชชด kit GF 1 AMBICLEAN KIT
PCR& Gel (Vivantis) กอนทจะท าการสง PCR product ใหกบบรษท Macrogen ประเทศเกาหล เพอวเคราะหล าดบน
วคลโอไทด (DNA sequences) ท าการวเคราะหล าดบนวคลโอไทดโดยใชโปรแกรม Seqman และโปรแกรม BLASTN
ใน NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi.PAGE_TYPE=BlastSearch) การจดกลมเปน sequence type
(ST) โดยใชโปรแกรมจาก website PubMLST (http://pubmlst.org/abaumannii/)

92
ตาราง 20 ขอมล primer ของ house keeping gene 7 ยน
gene Primer sequence
(5’3’)
Product
size
(bp)
Tm
(˚C)
references
rpoD ACCCGTGAAGGTGAAATCAG TTCAGCTGGAGCTTTAGCAAT
672 56 Wisplinghoff, et al., 2010
recA CCTGAATCTTCYGGTAAAAC GTTTCTGGGCTGCCAAACATTAC
425 56 Wisplinghoff, et al., 2010
gltA AATTTACAGTGGCACATTAGGTCCC GCAGAGATACCAGCAGAGATACACG
722 56 Wisplinghoff, et al., 2010
gryB TGAAGGCGGCTTATCTGAGT GCTGGGTCTTTTTCCTGACA
594 56 Wisplinghoff, et al., 2010
cpn_60 GGTGCTCAACTTGTTCGTGA CACCGAAACCAGGAGCTTTA
640 58 Wisplinghoff, et al., 2010
gpi GGAATTTCCGGAGCTCACAA TCAGGAGCAATACCCCACTC
456 56 Wisplinghoff, et al., 2010
gdhB GCTACTTTTATGCAACAGAGCC GTTGAGTTGGCGTATGTTGTGC
774 58 Wisplinghoff, et al., 2010

93
ผลการวจย (Result)
1. ผลการตรวจหายนดอยา (drug resistance genes) ในเชอ XDR-AB
ผลการตรวจหายนดอยา class A, B, C และ D ในเชอ XDR-AB ทง 27 ไอโซเลทแสดงดงภาพ 11 พบวา ยนดอยาใน
class A ตรวจพบเฉพาะยน blaPER-1 1 ไอโซเลท คดเปนรอยละ 3.7 (1/27) สวนยนอนใน class A ไดแก blaTEM,
blaSHV , blaCTX-M และ blaGES ตรวจไมพบ ยนดอยาใน class B ตรวจพบยน blaNDM-1 1 ไอโซเลทจาก XDR-
AB ทงหมด 27 ไอโซเลท คดเปนรอยละ 3.7 (1/27) สวนยนอนใน class B ไดแก blaVIM, blaGIM, blaSIM, blaSPM ตรวจ
ไมพบ ยนดอยาใน class C ตรวจพบยน blaADC รอยละ 100 (27/27) สวนยนอนใน class C ไดแก blaACC, blaMOX,
blaCIT, blaDHA, blaEBC ตรวจไมพบ ยนดอยาใน class D ตรวจพบยน blaOXA-23 และ blaOXA-51 รอยละ 100 (27/27)
สวนยน blaOXA-24 และ blaOXA-58 ตรวจไมพบ
ภาพ 11 รอยละความชก (% prevalence) ของยนดอยา (drug resistance genes) ทพบไดในแตละ class
(class A; blaPER-1, class B; blaNDM-1, class C; blaADC, class D; blaOXA-23, blaOXA-51)

94
2. การศกษารปแบบทางระบาดวทยาของเชอ XDR-AB โดยวธ blaOXA-51 sequence base typing
จากการจดรปแบบการระบาดของยน blaOXA-51 ในเชอ XDR-AB ทง 27 ไอโซเลท โดยการท า PCR ตรวจหา
ยน blaOXA-51 และน าไปผลการวเคราะหล าดบนวคลโอไทดไปแปลงเปนกรดอะมโนและเปรยบเทยบกบฐานขอมล
กรดอะมโนของโปรตน OXA-51 เพอระบกลมยอย (subgroup) สามารถจดกลมไดเปน 4 กลมยอย คอ กลม OXA-64
จ านวน 1 ไอโซเลท กลม OXA-83 จ านวน 24 ไอโซเลท กลม OXA-104 จ านวน 1 ไอโซเลท และ กลม OXA-508
จ านวน 1 ไอโซเลท (ตาราง 21)
3. การศกษารปแบบทางระบาดวทยาของเชอ XDR-AB โดยวธ REP-PCR
ผลของ REP-PCR types ของเชอทงหมด 339 ไอเลทของเชอ 4 จ านวนโรงพยาบาลในประเทศไทย ให REP-
PCR types จ านวน 33 รปแบบโดยใหรปแบบของ DNA patterns ทม amplicon sizes ขนาดอยในชวง 400 to 5,000
bp. โดย genotype จะตงชอเปน R1 to R33 ดงแสดงในภาพ 12A โดยเชอ A. baumannii จากโรงพยาบาลในจงหวด
นครสวรรคให REP-PCR type จ านวน 14 genotypes เชอ A. baumannii จากโรงพยาบาลในจงหวดพจตรให REP-
PCR type จ านวน 6 genotypes เชอ A. baumannii จากโรงพยาบาลในจงหวดเชยงรายให REP-PCR type จ านวน
19 genotypes และ เชอ A. baumannii จากโรงพยาบาลในจงหวดฉะเชงเทราให REP-PCR type จ านวน 13
genotypes เมอศกษษขอมลเฉพาะเชอ XDR-AB ทง 27 ไอโซเลท พบวา XDR-AB ให REP-PCR type จ านวน 3
genotypes คอ 4 แบบคอ R1, R4, R15 และ R16 (ตาราง 21 และภาพ 12B)

95
ตาราง 21 การระบกลมยอยของโปรตน OXA-51 และ REP-PCR types ของเชอ XDR-AB 27 ไอโซเลท
รหสเชอ % identity REP-PCR types AB16 OXA-83 (99%) 4 AB22 OXA-83 (99%) 4 AB28 OXA-83 (99%) 4 AB53 OXA-83 (99%) 15 AB57 OXA-83 (99%) 15 AB59 OXA-83 (99%) 4 AB81 OXA-83 (99%) 4 AB89 OXA-83 (99%) 4 AB95 OXA-83 (99%) 4 AB135 OXA-83 (99%) 4 AB136 OXA-64 (100%) 16 AB140 OXA-83 (99%) 4 AB180 OXA-83 (99%) 4 AB183 OXA-83 (99%) 4 AB229 OXA-83 (99%) 4 AB241 OXA-83 (99%) 4 AB246 OXA-508 (100%) 4 AB269 OXA-83 (99%) 4 AB320 OXA-83 (99%) 4 AB324 OXA-83 (99%) 4 AB329 OXA-83 (99%) 4 AB335 OXA-83 (99%) 4 AB367 OXA-83 (99%) 4 AB368 OXA-83 (99%) 4 AB371 OXA-83 (99%) 16 AB385 OXA-83 (99%) 4 AB405 OXA-104 (99%) 1

96
ภาพ 12 REP-PCR types ของเชอ A. baumannii จาก โรงพยาบาลในประเทศไทย
A) REP-PCR types ของเชอทงหมด 339 ไอเลทจาก 4 โรงพยาบาลไดแก HA: โรงพยาบาลในจงหวดนครสวรรค HB:
โรงพยาบาลในจงหวดพจตร HC: โรงพยาบาลในจงหวดเชยงราย และ HD: โรงพยาบาลในจงหวดฉะเชงเทรา B)แสดง
REP-PCR types ของเชอ A. baumannii กลมไมดอยา (NR-AB) กลมทมยน balNDM-1 (NDM-AB) และกลมดอยาแบบ
รนแรง (XDR-AB);
4. การศกษารปแบบทางระบาดวทยาของเชอ XDR-AB โดยวธ Multilocus sequence typing (MLST)
การจดรปแบบการระบาดโดยใชวธ MLST สามารถจดกลมเชอ XDR-AB ทง 27 ไอโซเลทโดยเชอจ านวน 23 ไอ
โซเลท สามารถจดกลมไดเปน 6 STs ไดแก ST195, ST208, ST368, ST451, ST457 และ ST1166 (ภาพ 13) และ เชอ
XDR-AB 4 ไอโซเลท ทไมถกจดอยใน STs ใด (non-type) (ภาพ 13) เชอ XDR-AB ทถกจดอยใน ST195 ไดแก AB81,
AB135, AB229 และ AB367 เชอ XDR-AB ทถกจดอยใน ST208 ไดแก AB95, AB180, AB183 และ AB246 ST368
ไดแก AB22 ST451 ไดแก AB16, AB28, AB59, AB89 และ AB324 เชอทอยใน ST457 ไดแก AB241 และ AB269
เชอทถกจดอยใน ST1166 ไดแก AB320, AB329, AB335, AB368, AB371, AB385 และ AB405 สวนเชอทไมสามารถ
จดอยใน STs ใด (non-type) ไดแก AB53, AB57, AB136 และ AB140 ขณะนอยระหวางยนขอ ST ใหม

97
ภาพ 13 แผนภาพตนไมทสรางจาก house keeping genes 7 genes (MLST-base tree) ของเชอ XDR-AB 27 ไอโซเลท หมายเหต :hospital 1 = โรงพยาบาลจงหวดเชยงราย hospital 2 = โรงพยาบาลจงหวดพจตร hospital 3 = โรงพยาบาลจงหวดนครสวรรค hospital 4=โรงพยาบาลจงหวดฉะเชงเทรา; NT = non-type stain; อกษรยอแผนก (ward) ทท าการเกบตวอยาง (MED=แผนกอายรกรรมชาย, SUR=แผนกศลยกรรม, PED=แผนกกมารเวช, ICU=แผนกผปวยหนก, M=สงฆอาพาธ, MED-M=อายกรรมชาย, SUR-W=ศลยกรรมหญง, MWH=รพ.แมวงก

98
ขอวจารณ (Discussion)
1. ผลการตรวจหายนดอยา (drug resistance genes) ในเชอ XDR-AB
แบคทเรย A. baumannii เปนแบคทเรยทสามารถกอโรคตดเชอไดรนแรงและยากตอการรกษา โดยตวเชอน
สามารถกอโรคตดเชอในผปวยทนอนพกรกษาตวในหออภบาลผปวยหนก และผปวยทมภมคมกนต า นอกจากนใน
ปจจบน A. baumannii ยงมการปรบตวตอสภาพแวดลอมใหกลายเปนเชอทดอตอยาปฏชวนะและมระดบการดอยาท
เพมขน โดยเชอ A. baumannii ทดอยาในรปแบบ extensively-drug resistance A. baumannii (XDR-AB) เปน
กลมทมการระบาดไปทวโลกและมความรนแรงในการกอโรค เนองจากดอตอยาปฏชวนะทกกลม ยกเวน colistin และ
tigecycline ในประเทศไทยจากการศกษาของ Inchai และคณะ ในป ค.ศ.2015 พบรอยละความชกของ XDR-AB
เทากบ 65.3 ในการศกษาของผวจยในครงนพบการรอยละความชกการระบาดของ XDR-AB เทากบ 7.96 การตรวจหา
ยนทดอตอยาปฏชวนะในกลมบตาแลคทาเมส ทง 4 classes ไดแก class A, B, C และ D ยนดอยาใน class A ตรวจ
พบยน blaPER-1 เพยงยนเดยว ซงไมพบยนดอยาอนใน class A ไดแก ยน blaCTX-M, blaTEM, blaSHV และ blaGES ยน
ดอยาใน class B ไดแก blaNDM-1, blaGIM, blaVIM และ blaSPM ซงใน class นพบเพยงยน blaNDM-1 เพยงยนเดยว รอย
ละความชกเทากบ 3.7 ใน class C ยนทตรวจหาไดแก blaADC, blaACC, blaMOX, blaCIT, blaDHA, blaEBC ผลตรวจพบ
ยน blaADC เพยงยนเดยวคดเปนรอยละ 100 สวนยนอนใน class C ไดแก blaACC, blaMOX, blaCIT, blaDHA, blaEBC
ตรวจไมพบ ใน class D ยนทตรวจหา ไดแก blaOXA-23, blaOXA-24, blaOXA-51 และ blaOXA-58 จากผลพบยน blaOXA-23
และ blaOXA-51 รอยละ 100 สวนยน blaOXA-24 และ blaOXA-58 นนตรวจไมพบ จากการศกษาในครงนสอดคลองกบ
การศกษาของ Teo และคณะในป ค.ศ.2015 ไดท าการตรวจหายนดอยา แตไมพบยนดอยาใน class B และพบวา ยน
ใน class D สวนใหญจะพบความชกของยน blaOXA-51 มากทสด รองลงมาคอ blaOXA-23 และทพบไดไมมากคอ blaOXA-
58 เนองจากยน blaOXA-51 เปน intrinsic gene ของเชอ A. baumannii จงพบไดในเชอ A. baumannii ทกตว สวน
ยน blaOXA-23 เปนยนทพบไดมากในประเทศแถบเอเชยหรอตอนใตของเอเชย โดยพบ ยน blaOXA-23 ในประเทศ
สงคโปรรอยละ 91 ประเทศจนและเกาหลรอยละ 40-100 (Koh, et al., 2007; Wang, et al., 2013; Teo, et al.,
2015) และนอกจากนการศกษานยงสอดคลองกบการศกษาของ Farshadzadeh และคณะในป ค.ศ.2015 ทท าการ
ตรวจหายนดอยาทง 4 classes ไดแก class A, B, C และ D ในเชอ XDR-AB ทแยกไดจากโรงพยาบาลในประเทศ

99
อหราน จากผลตรวจพบ ยนดอยาใน class A คอ blaPER-1 เพยงยนเดยว ตรวจไมพบยนอนใน class A ไดแก blaCTX-
M, blaTEM และ blaSHV-1 ยนดอยาใน class B ไดแก blaNDM-1, blaGIM, blaVIM, blaSPM, blaVIM ตรวจไมพบ สวนยนดอ
ยาใน class C ทตรวจหา ไดแก blaADC พบไดรอยละ 100 (Farshadzadeh, et al., 2015)
2. การศกษารปแบบทางระบาดวทยาของเชอ XDR-AB
การจดรปแบบการระบาดของเชอโดยวธทางจโนไทป เปนวธทถกตอง แมนย าสงและมความนาเชอถอ ใน
การศกษาครงน ท าการศกษาการจดรปแบบการระบาดโดยวธทางจโปไทป 3 วธ คอ blaOXA-51 sequence base
typing (blaOXA-51 SBT) ว ธ Repetitive-Polymerase chain reaction (REP-PCR) แ ล ะ Multilocus sequence
typing (MLST) ซงทง 3 วธนเปนวธการศกษารปแบบการระบาดทเปนทนยมใชในการศกษาการระบาด แตในแตละวธ
อาจมขอดและขอเสยตางกน วธ blaOXA-51 SBT มขอด คอ วธการศกษาไมยากมากและเปนวธทเหมาะสมใชในขนตอน
การศกษารปแบบการระบาดในเบองตน (screening) แตกมขอเสยคอเนองจากเมอท า PCR ตรวจหายนแลว ตองสงไป
วเคราะหล าดบนวคลโอไทดใชงบประมาณคอนขางสงและขอเสยอกเหตผลหนงคอ การจดรปแบบการระบาดทไมคอย
ละเอยดมากนก (low discriminatory power) วธ REP-PCR มขอด คอ เปนวธทรวดเรว (rapidly) แตขอเสย คอ การ
วเคราะหผล หลงการท า PCR บางครงไมมเกณฑทแนนอนในการแปลผล เนองจากเปนการจดรปแบบโดย DNA
fingerprint ในแตละการวจยอาจแปลผลและจดกลมทตางกนไดและวธ MLST เปนวธการจดรปแบบการระบาดทเปน
มาตรฐาน (gold standard) เปนวธทนาเชอถอ เนองจากผลทวเคราะหล าดบนวคลโอไทดมการเปร ยบเทยบกบ
ฐานขอมลขนาดใหญ ไดแก NCBI นอกจากนวธ MLST ยงเปนวธทสามารถท าซ าในหองปฏบตการคนละแหงแลวใหผล
ทเหมอนกนได(reproducibility) แตขอเสยคอ ตองใชงบประมาณสงเนองจากตองมการสงล าดบนวคลโอไทดไป
วเคราะห (Rafei, et al., 2014)
การจดรปแบบการระบาดโดยวธ REP-PCR ในการศกษานสามารถจดรปแบบการระบาดไดทงหมด 4 รปแบบ
ตามขนาดและจ านวนแถบลายพมพดเอนเอ (DNA fingerprint) คอ รปแบบ 1, 4, 15 และ 16 ซงจะพบวา จ านวนเชอ
ทมรปแบบการระบาดแบบท 4 มจ านวนมากทสดและพบรปแบบนไดในโรงพยาบาลทง 4 แหง คดเปนรอยละเทากบ
81.48 รปแบบการระบาดทมความชกรองลงมาคอ รปแบบการระบาดแบบท 15 คดเปนรอยละ 7.40 และรปแบบท 16
คดเปนรอยละ 7.40 ซงรปแบบท 16 พบไดในเชอทแยกไดจากโรงพยาบาลจงหวดพจตรและโรงพยาบาลจงหวด
ฉะเชงเทรา และรปแบบการระบาดแบบท 15 พบไดเฉพาะเชอทแยกไดจากโรงพยาบาลจงหวดนครสวรรค และรปแบบ

100
การระบาดแบบท 1 ทพบไดนอยสด คดเปนรอยละ 3.7 พบในเชอทแยกไดจากโรงพยาบาลจงหวดฉะเชงเทรา ในการ
แปลผลรปแบบการระบาดโดยวธ REP-PCR อาจไมสามารถบอกขอมลการกระจายตวของเชอไดมาก เนองจากการ
จดรปแบบการระบาดโดยวธน ไมมความละเอยดในการจดกลมมากนก รปแบบการระบาดทพบจงมความหลากหลายไม
มาก แตขอมลรปแบบการระบาด อาจบอกไดเพยงคราวๆ วามการระบาดของเชอรปแบบใดบาง เชน เชอรปแบบการ
ระบาดแบบท 15 พบไดเฉพาะเชอทแยกไดจากโรงพยาบาลจงหวดนครสวรรคและเชอทพบทง 2 ไอโซเลท มาจากทมา
ทตางกน คอ มาจากแผนกกมารเวชและแผนก ICU อายรกรรม อาจแปลผลไดวาเชอนนาจะมการระบาดจากแผนก
หนงไปยงอกแผนกหนงภายในโรงพยาบาลเดยวกน แตหากตองการยนยนวามการระบาดระหวางแผนกในโรงพยาบาล
หรอระบาดข ามโรงพยาบาล ควรใช ว ธ จ ดรปแบบการระบาด อ นยนยนร วมด วย จากการศกษาของ
Santimaleeworagun และคณะ ในป ค.ศ.2014 ท าการศกษารปแบบการระบาดโดยวธ REP-PCR ซงแยกเชอไดจาก
ผปวยแผนก ICU ในโรงพยาบาล ประเทศไทย พบรปแบบการระบาดได 4 กลม คอ clone A, B, C และ D และพบวา
clone A เปนรปแบบการระบาดทพบไดมากทสด (Santimaleeworagun, et al., 2014)
การจดรปแบบการระบาดโดยวธ blaOXA-51 SBT ในการศกษาครงนสามารถพบการระบาดของกลมยอยยน
blaOXA-51 ได 4 กลมยอย ไดแก OXA-64, OXA-83, OXA-104 และ OXA-508 ซงยนกลมยอย OXA-83 พบไดมาก
ทสด และอก 3 กลมยอย ทพบเพยงกลมยอยละ 1 ไอโซเลท คอ OXA-64, OXA-104และ OXA- 508 คดเปนรอยละ
ความชกเทากบ 88.88, 3.7, 3.7 และ 3.7 ตามล าดบ โดยเชอทแยกไดจากโรงพยาบาลจงหวดนครสวรรคพบการระบาด
เพยงรปแบบเดยว คอ รปแบบการระบาดของกลม OXA-83 โรงพยาบาลจงหวดพจตรพบการระบาด 2 รปแบบ คอ
การระบาดของกลม OXA-64 และ OXA-83 เชอทแยกไดจากโรงพยาบาลเชยงรายพบการระบาด 2 รปแบบ คอ
การระบาดของกลม OXA-83 และ OXA-508 และเชอทแยกไดจากโรงพยาบาลจงหวดฉะเชงเทรา พบการระบาด 2
รปแบบ คอ OXA-83 และ OXA-405 พบวาเชอ XDR-AB ทแยกไดจากโรงพยาบาลจงหวดนครสวรรค ทกไอโซเลทถก
จดอยในกลมยอย OXA-83 และพบไดจากเชอทแยกมาจากผปวยคนละแผนกหรอมาจากแผนกเดยวกน ซง เชอทแยก
ไดมาจากผปวยคนละคน เชน เชอ AB22 มรปแบบการระบาดกลมยอย OXA-83 แยกเชอไดจากแผนกอายรกรรมชาย
มรปแบบการระบาดเหมอนกบ AB28 ทแยกไดมาจากแผนกเดยวกนและเหมอนกบ AB53 ทแยกไดจากแผนกกมารเวช
นอกจากนรปแบบการระบาดของกลมยอย OXA-83 ยงพบไดจากเชอทแยกไดจากโรงพยาบาลจงหวดพจตร เชยงราย
และฉะเชงเทรา จากขอมลการระบาดน สามารถแปลผลไดวา รปแบบการระบาดกลมยอย OXA-83 อาจมการระบาด

101
ทงภายในโรงพยาบาลและระหวางโรงพยาบาลทง 4 แหง จากการศกษาของ Pournaras และคณะ ในป ค.ศ. 2014
ท าการศกษาการระบาดโดยวธ blaOXA-51 SBT โดยแยกเชอจากหลายประเทศ ไดแก ไดแก อตาล กรซ เลบานอน ตลก
อเมรกาและเยอรมน ผลการศกษาพบการระบาดของ subgroup ของโปรตน OXA-51 ไดหลายกลม ไดแก OXA-66, -
69, -71, -64, -90, -94, -365, -51, -128, -86, -100 และ OXA-65 เปนตน (Pournaras, et al., 2014) และจาก
การศกษาของ Rafei และคณะ ในป ค.ศ. 2015 ท าการศกษาการระบาดของเชอ A. baumannii โดยใชวธ blaOXA-51
SBT ซงแยกเชอไดจากโรงพยาบาล ในเมอง Tripoli ประเทศเลบานอน ผลการศกษาพบการระบาดของ subgroup
ของโปรตน OXA-51 ไดหลายกลมและเรยงจากปรมาณการตรวจพบตามล าดบ ไดแก OXA-66 (พบมากทสด), -69, -
64, -94, -120, -70, -71, -104, -121, -132 และ OXA-406 (เปนโปรตนชนดใหมทพบครงแรกของ subgroup ของ
โปรตน OXA-51) เปนตน (Rafei, et al., 2015)
การจดรปแบบการระบาดโดยวธ MLST ในการศกษาครงนเชอ XDR-AB จากทงหมด 27 ไอโซเลท สามารถ
จดกลมเปน sequence typing (STs) ได 23 ไอโซเลท สวนเชออก 4 ไอโซเลท ไมถกจดอยในกลมใด (non-type) กลม
ทเปน non-type เมอได allele number ออกมาทง 7 ยนแลว น าไปวเคราะหเปรยบเทยบกบฐานขอมลของเชอ
Acinetobacter baumannii ในเวบไซต PubMLST พบวา ไมตรงกบ STs ใด สวนเชอ XDR-AB 23 ไอโซเลท พบการ
ระบาดในรปแบบ STs ได 6 STs ไดแก ST195, ST208, ST368, ST451, ST457 และ ST1166 จากการน าล าดบน
วคลโอไทดของ house keeping genes ทง 7 ยน มาสรางแผนภาพ pylogenetic tree (MLST-base tree) ท าใหได
รวา มการระบาดของเชอทเปน STs เดยวกน มการระบาดเกดขนทงภายในโรงพยาบาลเดยวกนและระบาดระหวาง
โรงพยาบาล (ภาพ 9) จากเชอทงหมดทแยกไดจากโรงพยาบาลจงหวด 4 แหง เชอจากโรงพยาบาลจงหวดนครสวรรค
ประกอบดวย ST195, ST368, ST451 และเชออก 2 ไอโซเลททเปน non-type เชอทแยกจากโรงพยาบาลจงหวดพจตร
พบวาถกจดอยใน ST195 จ านวน 2 ไอโซเลท และเปน non-type 1 ไอโซเลท เชอทแยกไดจากโรงพยาบาลจงหวด
เชยงราย พบวาถกจดอยใน ST208 จ านวน 4 ไอโซเลท และเปน ST457 2 ไอโซเลท สวนเชอทแยกไดจากโรงพยาบาล
จงหวดฉะเชงเทรา เชอจ านวน 7 ไอโซเลทถกจดอยใน ST1166 1 ไอโซเลท ถกจดอยใน ST195 และเชออก 1 ไอโซ
เลท ทถกจดอยใน ST451 เชอทแยกจากโรงพยาบาลจงหวดนครสวรรคมความหลากหลายมากทสด เนองจากพบวา
สามารถถกจดอยใน STs หลากหลายกลม และในเชอบางไอโซเลท ไดแก AB53 และ AB57 ทไมถกจดอยในกลมใดเลย
(non-type) ม allelic profile ของ house keeping genes 7 ยนเหมอนกน คอ 2-140-15-1-4-18-3 เมอจดกลมการ

102
ระบาดดวยวธ blaOXA-51 SBT ถกจดอยในกลมยอย OXA-83 และถกจดอยในรปแบบการระบาดแบบท 15 เหมอนกน
ทง 2 ไอโซเลท แตทงเชอ AB 53 และ AB57 ถกแยกมาจากคนละแผนก โดยเชอ AB53 ถกแยกมาจากผปวยแผนก
กมารเวช สวน AB57 แยกมาจากผปวยแผนกผปวยวกฤตอายรกรรม จากขอมลนเมอเปรยบเทยบการระบาดของเชอทง
2 ไอโซเลทน จากวธการจดรปแบบการระบาดทง 3 วธ สามารถแปลผลไดวา เชอทง 2 ไอโซเลทน อาจเปนเชอสาย
พนธเดยวกน ทมการระบาดระหวางแผนกภายในโรงพยาบาลเดยวกน เชอทแยกไดจากโรงพยาบาลจงหวดฉะเชงเทรา
จ านวน 7 ไอโซเลท จากทงหมด 8 ไอโซเลท ถกจดอยใน ST1166 แตเชอทง 7 ไอโซเลท ถกแยกมาจากผปวยทงแผนก
เดยวกนและคนละแผนก แตสวนใหญแยกมาจากแผนกเดยวกน คอ แผนกอายร-กรรมชาย มเพยง 1 ใน 7 ไอโซเลทท
แยกมาจากตางแผนก คอ แผนกศลยกรรมหญง จากขอมลนสามารถแปลไดวา อาจมการระบาดของเชอภายใน
โรงพยาบาลน ทงระบาดในแผนกเดยวกนและมการแพรกระจายไปสผปวยตางแผนก ถงแมเชอทง 7 ไอโซเลท เมอ
จดรปแบบการระบาดดวยวธ blaOXA-51 SBT และ REP-PCR อาจมบางไอโซเลท ทมรปแบบการระบาดทตางจากเชอไอ
โซเลทอนทถกจดอยใน STs เดยวกน แตดวยวธการจดรปแบบการระบาดทง 3 วธ การจดรปแบบการระบาดดวยวธ
blaOXA-51 SBT นนเหมาะสมกบการจดรปแบบการระบาดในขนตน วธ REP-PCR เปนการจดรปแบบการระบาดแบบไม
ละเอยดมากนก เนองจากเปนเพยงการเปรยบเทยบความตางและขนาดของแถบลายพมพดเอนเอ (DNA fingerprint)
สวนการจดรปแบบการระบาดดวยวธ MLST เปนวธทเปนมาตรฐาน มความถกตองและแมนย ามากทสด มฐานขอมล
ขนาดใหญส าหรบเปรยบเทยบและวเคราะหผลจาก house keeping genes ทง 7 ยน ในรปแบบ STs การแปลผลการ
ระบาดของเชอทแยกไดจากโรงพยาบาลจงหวดฉะเชงเทรา จงควรเชอถอรปแบบการระบาดทจดโดยวธ MLST จง
สามารถแปลผลไดวา เชอทแยกจากโรงพยาบาลจงหวดฉะเชงเทราทง 7 จาก 8 ไอโซเลท อาจเปนเชอสายพนธเดยวกน
ทมการระบาดระหวางแผนกตางๆ ภายในโรงพยาบาลเดยวกน (epidemic clone) นอกจากนเชอ AB81, AB135,
AB229 และ AB367 ซงเชอทง 4 ไอโซเลทนบางไอโซเลทแยกจากผปวยโรงพยาบาลเดยวกนและบางไอโซเลทแยกจาก
คนละโรงพยาบาล เชอ AB81 แยกไดจากโรงพยาบาลจงหวดนครสวรรค เชอ AB135 และ เชอ AB229 ทง 2 ไอโซเลท
น แยกไดจากแผนกศลยกรรมชายโรงพยาบาลจงหวดพจตร และเชอ AB 367 แยกจากโรงพยาบาลจงหวดฉะเชงเทรา
ซงเชอทง 4 ไอโซเลท ถกจดอยใน STs เดยวกน คอ ST195 ซงอาจแปลผลไดวามการระบาดทงภายในโรงพยาบาล
เดยวกนและมการระบาดระหวางโรงพยาบาล

103
จากการศกษาการระบาดของเชอ A. baumannii ทดอตอยาปฏชวนะ พบวา การระบาดของ ST195
สามารถพบไดบอยในหลายประเทศ ไดแก ประเทศจน โคเอเทย เดนมารกและมาเลเซย (Li, et al., 2015; Liu, et al.,
2016; Seruga, et al., 2017) การระบาดของ ST208 สามารถพบไดประเทศจนและสงคโปร (Li, et al., 2015; Liu,
et al., 2016; Blackwell, et al., 2017) สวนการระบาดของ ST368 สามารถพบการระบาดไดมากทสดในประเทศจน
และพบไดในประเทศอหราน (Farshadzadeh, et al., 2015; Huang, et al., 2016) การระบาดของ ST451 พบวาม
ความชกสงในประเทศจน (Li, et al., 2015; Liu, et al., 2016) ในประเทศฮองกงและจนยงพบการระบาดไดมากของ
ST457 (Cheng, et al., 2015) และการระบาดของ ST1166 ทเพงพบเปน novel ST ทเกดจาก trilocus variant มา
จาก ST208 ซงพบการระบาดครงแรกของ ST1166 ในป ค.ศ. 2017 จาการศกษาของ Blackwell และคณะ ท
ท าการศกษาในเชอ XDR-AB ซงเชอทท าการศกษามาจากเชอทถกเกบใหคงสภาพในโรงพยาบาลประเทศสงคโปร
การศกษาของ Li และคณะ ในป ค.ศ.2015 ยงพบวา มการระบาดของเชอทถกจดใน ST92 ในหลายประเทศทวโลก
(global dissemination) ไดแก ประเทศสเปน เกาหล ฮองกง ออสเตรเลยและอเมรกา นอกจาก ST92 แลวการระบาด
ของเชอทมการรปแบบการระบาดแบบ ST2 ยงเปนการระบาดในกลมท ถกจดวา มการระบาดในหลายประเทศทวโลก
(global dissemination) (Blackwell, et al., 2017)

104
สรปและขอเสนอแนะ (Conclusion and Recommendation)
สรปผลงานวจย
จากการศกษาในครงน เชอ XDR-AB ทใชในการศกษามทงหมด 27 ไอโซเลท ตรวจพบยนดอยา (drug
resistance genes) ทง 4 classes ตรวจพบยนดอยาใน class A เพยงยน blaPER-1 และใน class B ตรวจพบเพยงยน
blaNDM-1 เพยงยนเดยว ยนดอยาใน class C ตรวจพบยน blaADC เพยงยนเดยว ยนดอยาใน class D ตรวจพบยน
blaOXA-23 และ blaOXA-51 การศกษารปแบบการระบาดโดยวธ REP-PCR ในการศกษานสามารถจดรปแบบการระบาด
ไดทงหมด 4 รปแบบ ตามขนาดและจ านวนแบนดของลายพมพดเอนเอ (DNA fingerprint) คอ รปแบบ 1, 4, 15 และ
16 การศกษารปแบบการระบาดโดยวธ blaOXA-51 SBT ในการศกษาครงนสามารถพบการระบาดของกลมยอยยน
blaOXA-51 ได 4 กลมยอย ไดแก OXA-64, OXA-83, OXA-104 และ OXA-508 และจากการจดกลมการระบาดโดยวธ
Multilocus sequence typing (MLST) เชอ XDR-AB ทงหมด 27 ไอโซเลท สามารถจดกลมเปน STs ได 23 ไอโซเลท
และอก 4 ไอโซเลททไมถกจดอยใน STs ใดเลย(non-type) เชอ 23 ไอโซเลท ถกจดอยใน STs ทมความหลากหลาย
ไดแก ST195, ST208, ST368, ST451, ST457 และ ST1166
ขอเสนอแนะ (Conclusion and Recommendation)
จากขอมลรปแบบการระบาดของเชอ XDR-AB หากพบวา เชอทแยกไดจากตวอยางผปวยในแผนกเดยวกน ระหวาง
แผนกในโรงพยาบาลเดยวกนหรอระหวางโรงพยาบาล เปนเชอทมรปแบบการระบาดทเหมอนกน จงอาจสามารถแปล
ผลไดวา มการระบาดของเชอสายพนธเดยวกน ขอมลตรงนจงสามารถเปนขอมลสงกลบใหกบโรงพยาบาลหรอบคลากร
ทางการแพทยใหมการปรบปรงดานสขอนามยทงตอผคนและสงแวดลอมในโรงพยาบาลเพอปองกนและลดการ
แพรกระจายของเชอดอยาใหมอตราการกระจายทลดลง

105
เอกสารอางอง (reference)
1. ภทรชย กรตสน และ อนชา อภสารธนรกษ. (2555). การจดกลมเอนไซมในกลม beta-lactamase, Beta-
lactamase ในแบคทเรยแกรมลบจากความรพนฐานสเวชปฏบต. กรงเทพฯ: ว.เจ. พรนตง
2. Ahmed, S. S. & Alp, E. (2015). Genotyping methods for monitoring the epidemic evolution of
A. baumannii strains. The Journal of Infection in Developing Countries, 9(4), 347-354.
3. Bartual, S. G., Seifert, H., Hippler, C., Luzon, M. A., Wisplinghoff, H., & Rodriguez-Valera, F.
(2005). Development of a multilocus sequence typing scheme for characterization of clinical
isolates of Acinetobacter baumannii. Journal of Clinical Microbiology, 43(9), 4382-4390.
4. Ben-Darif, E., De Pinna, E., Threlfall, E. J., Bolton, F. J., Upton, M., & Fox, A. J. (2010).
Comparison of a semi-automated rep-PCR system and multilocus sequence typing for
differentiation of Salmonella enterica isolates. Journal of Microbiological Methods, 81(1),
11-16.
5. Blackwell, G. A., Holt, K. E., Bentley, S. D., Hsu, L. Y., & Hall, R. M. (2017). Variants of AbGRI3
carrying the armA gene in extensively antibiotic-resistant Acinetobacter baumannii from
Singapore. Journal of Antimicrobial Chemotherapy, 72(4), 1031-1039.
6. Bogaerts, P., Naas, T., El Garch, F., Cuzon, G., Deplano, A., Delaire, T., Huang, T. D., Lissoir, B.,
Nordmann, P., & Glupczynski, Y. (2010). GES Extended-Spectrum β-Lactamases in
Acinetobacter baumannii Isolates in Belgium. Antimicrobial agents and chemotherapy,
54(11), 4872-4878.
7. Bou, G., Cerveró, G., Domínguez, M. A., Quereda, C., & Martínez-Beltrán, J. (2000). PCR-based
DNA fingerprinting (REP-PCR, AP-PCR) and pulsed-field gel electrophoresis characterization of
a nosocomial outbreak caused by imipenem- and meropenem-resistant Acinetobacter
baumannii. Clinical Microbiology and Infection, 6(12), 635-643.

106
8. Chan, M. C., Chiu, S. K., Hsueh, P. R., Wang, N. C., Wang, C. C. & Fang, C. T. (2014). Risk Factors
for healthcare-associated extensively drug-resistant Acinetobacter baumannii Infections: A
Case-Control Study. PLoS One, 9(1), 1-7.
9. Cheng, V. C., Chen, J. H. , Poon, R. W. , Lee, W. M., So, S. Y., Wong, S. C., Chau, P. H., Yip, C.
C., Wong, S. S., Chan, J. F., Hung, I. F., Ho, P. L., & Yuen, K. Y. (2015). Control of hospital
endemicity of multiple-drug-resistant Acinetobacter baumannii ST457 with directly observed
hand hygiene. European Journal of Clinical Microbiology & Infectious Diseases, 34(4), 713-
718.
10. Dahiya, S., Singla, P., Chaudhary, U., & Singh, B. (2015). Carbapeneamase review. International
Journal of Advanced Health Sciences, 2(4), 11-18.
11. D'Arezzo, S., Capone, A., Petrosillo, N., Visca P., GRAB, Ballardini, M., Bartolini, S., Bordi, E., Di
Stefano, A., Galiè, M., Minniti, R., Meledandri, M., Pacciani, L., Parisi, G., Prignano, G., Santini,
C., Valmarin, M., Venditti, M., Ziantoni, S. (2009). Epidemic multidrug-resistant Acinetobacter
baumannii related to European clonal types I and II in Rome (Italy). Clinical Microbiology
and Infection, 15(4), 347–357.
12. Evans, B. A., Hamouda, A., Towner, K. J., & Amyes, S. G. B. (2008). OXA-51-like β-lactamases
and their association with particular epidemic lineages of Acinetobacter baumannii. Clinical
Microbiology and Infection, 14(3), 268-275.
13. Farshadzadeh, Z., Hashemi, F. B., Rahimi, S., Pourakbari, B., Esmaeili, D., Haghighi, M. A.,
Majidpour, A., Shojaa, S., Rahmani, M., Gharesi, S., Aziemzadeh, M., & Bahador, A. (2015). Wide
distribution of carbapenem resistant Acinetobacter baumannii in burns patients in Iran.
Frontiers in Microbiology, 6, 1-10.
14. Gomaa, F. A. M., Helal, Z. H., & Khan, M. I. (2017). High Prevalence of blaNDM-1, blaVIM, qacE,
and qacE 1 Genes and Their Association with Decreased Susceptibility to Antibiotics and

107
Common Hospital Biocides in Clinical Isolates of Acinetobacter baumannii. Microorganisms,
5(2), 1-15.
15. Hasanin, A., Mukhtar, A., El-adawy, A., Elazizi, H., Lotfy, A., Nassar, H., & Ghaith, D. (2016).
Ventilator associated pneumonia caused by extensive-drug resistant Acinetobacter
species:Colistin is the remaining choice. Egyptian. Journal of Anaesthesia, 32(3), 409-413.
16. Hojabri, Z., Pajand, O., Bonura, C., Aleo, A., Giammanco, A., & Mammina, C. (2014). Molecular
epidemiology of Acinetobacter baumannii in Iran: endemic and epidemic spread of
multiresistant isolates. Journal of Antimicrobial Chemotherapy, 69(9), 2383-2387.
17. Hout, B., Oum, C., Men, P., Vanny, V., Supaprom, C., Heang, V., Rachmat, A., Prouty, M.,
Newell, S., Harrison, D., Noor, S., Gollogly, J., Tho, L., Kim, Y. J., & Ford, G. (2015). Drug
resistance in bacteria isolated from patients presenting with wounds at a non-profit Surgical
Center in Phnom Penh, Cambodia from 2011-2013. Tropical Diseases, Travel Medicine and
Vaccines, 1(4), 1-11.
18. Inchai, J., Liwsrisakun, C., Theerakittikul, T., Chaiwarith, R., Khositsakulchai, W. & Pothirat, C.
(2015). Risk factor of multidrug-resistant, extensively drug-resistant and pandrug-resistant
Acinetobacter baumannii ventilator-associated pneumonia in a Medical Intensive Care Unit
of University Hospital in Thailand. Journal of Infection and Chemotherapy, 21(8), 570-574.
19. Joshi, P. R., Acharya, M., Kakshapati, T., Leungtongkam, U., Thummeepak, R., & Sitthisak, S.
(2017). Co-existence of blaOXA-23 and blaNDM-1 genes of Acinetobacter baumannii isolated
from Nepal: antimicrobial resistance and clinical significance. Antimicrobial resistance and
infection control, 6(21), 1-7.
20. Jiang, M., Liu, L., Ma, Y., Zhang, Z., Li, N., Zhang, F., & Zhao, S. (2016). Molecular
epidemiology of multi-drug resistant Acinetobacter baumannii isolated in Shandong, China.
Frontiers in microbiology, 1687(7), 1-9.

108
21. Kalai Blagui, S., Achour, W., Abbassi, M.S., Bejaoui, M., Abdeladhim, A., & Ben Hassen, A.
(2007) Nosocomial outbreak of OXA-18-producing Pseudomonas aeruginosa in Tunisia.
Clinical Microbiology and Infection, 13, 794–800.
22. Kolar, M., Bardon, J., Chroma, M., Hricova, K., Stosova, T., Sauer, P., & Koukalova, D. (2010).
ESBL and AmpC beta-lactamase producing Enterobacteriaceae in poultry in the Czech
Republic. Veterinary Medicine – Czech, 55, 119–124.
23. Koh, T. H., Sng, L. H., Wang, G. C., Hsu, L. Y., & Zhao, Y. (2007). IMP-4 and OXA beta-
lactamases in Acinetobacter baumannii from Singapore. Journal of Antimicrobial
Chemotherapy, 59(4), 627-632.
24. Lee, C. R., Lee, J. H., Park, M., Park, K. S., Bae, I. K., Kim, Y. B., Cha, C. J., Jeong, B. C., & Lee, S.
H. (2017). Biology of Acinetobacter baumannii: pathogenesis, antibiotic resistance
mechanisms, and prospective treatment options. Frontiers in cellular and infection
microbiology, 7(55), 1-35.
25. Lee, H. W., Koh, Y. M., Kim, J., Lee, J. C., Lee, Y. C., Seol, S. Y., Cho, D. T., & Kim, J. (2008).
Capacity of multidrug-resistant clinical isolates of Acinetobacter baumannii to form biofilm
and adhere to epithelial cell surfaces. Clinical Microbiology and Infection, 14(1), 49-54.
26. Leungtongkam, U., Thummeepak, R., Wongprachan, S., Thongsuk, P., Kitti, T., Ketwong, K.,
Runcharoen, C., Chantratita, N. & Sitthisak, S. (2017). Dissemination of blaOXA-23, blaOXA-24,
blaOXA-58, and blaNDM-1 genes of Acinetobacter baumannii isolates from four tertiary
hospitals in Thailand. Microbial drug resistance, 1-8. doi: 10.1089/mdr.2016.0248.
27. Li, Y., Guo, Q., Wang, P., Zhu, D., Ye, X., Wu, S., & Wang, M., (2015). Clonal dissemination of
extensively drug-resistant Acinetobacter baumannii producing an OXA-23 β-lactamase at a
teaching hospital in Shanghai, China. Journal of Microbiology, Immunology and Infection,
48(1), 101-108.

109
28. Liu, Y., & Liu, X. (2015). Detection of AmpC β-lactamases in Acinetobacter baumannii in the
Xuzhou region and analysis of drug resistance. Experimental and Therapeutic Medicine,
10(3), 933-936.
29. Mammina, C., Palma, D. M., Bonura, C., Aleo, A., Fasciana, T., Sodano, C., Saporito, M. A.,
Verde, M. S., Calà, C., Cracchiolo, A. N., & Tetamo, R. (2012). Epidemiology and clonality of
carbapenem-resistant Acinetobacter baumannii from an intensive care unit in Palermo, Italy.
BioMed Central Research Notes, 5(1), 1-8.
30. Moghnieh, R., Siblani, L., Ghadban, D., El Mchad, H., Zeineddine, R., Abdallah, D., Ziade, F.,
Sinno, L., Kiwan, O., Kerbaj, F., & El-Imad, Z. (2016). Extensively drug-resistant Acinetobacter
baumannii in a Lebanese intensive care unit: risk factors for acquisition and determination of
a colonization score. Journal of Hospital Infection, 92(1), 47-53.
31. Poirel, L., Bonnin, R. A., & Nordmann, P. (2011). Genetic basis of antibiotic resistance in
pathogenic Acinetobacter species. IUBMB Life, 63(12), 1061-1067.
32. Pournaras, S., Gogou, V., Giannouli, M., Dimitroulia, E., Dafopoulou, K., Tsakris, A., & Zarrilli, R.
(2014). Single-locus-sequence-based typing of blaOXA-51-like genes for rapid assignment of
Acinetobacter baumannii clinical isolates to international clonal lineages. Journal of Clinical
Microbiology, 52(5), 1653-1657.
33. Rafei, R., Dabboussi, F., Hamze, M., Eveillard, M., Lemarié, C., Gaultier, M. P., Mallat, H.,
Moghnieh, R., Husni-Samaha, R., Joly-Guillou, M. L., & Kempf, M. (2014). Molecular analysis of
Acinetobacter baumannii Strains Isolated in Lebanon Using Four Different Typing Methods.
PLoS ONE, 9(12), 1-15.
34. Rafei, R., Pailhoriès, H., Hamze, M., Eveillard, M., Mallat, H., Dabboussi, F., Joly-Guillou, M. L. &
Kempf, M. (2015). Molecular epidemiology of Acinetobacter baumannii in different

110
hospitals in Tripoli, Lebanon using bla (OXA-51-like) sequence based typing. BMC
microbiology, 15(1), 1-7.
35. Reboli, A. C., Houston, E. D., Monteforte, J. S., Wood, C. A., & Hamill, R.J. (1994).
Discrimination of epidemic and sporadic isolates of Acinetobacter baumannii. Journal of
Clinical Microbiology, 32(11), 2635–2630.
36. Santimaleeworagun, W., Thathong, A., Samret, W., Preechachuawong, P., Sae-lim, W., &
Jitwasinkul, T. (2014). Identification and characterization of carbapenemase genes in clinical
isolates of carbapenem-resistant Acinetobacter baumannii from general hospital in Thailand.
Southeast Asian Journal of Tropical Medicine and Public Health, 45(4), 874-850.
37. Seruga, M. M., Hrenovic, J., Goic-Barisic, I., Hunjak, B., Skoric, D., & Ivankovic, T. (2017).
Emission of extensively-drug-resistant Acinetobacter baumannii from hospital settings to the
natural environment. Journal of Hospital Infection, 96(4), 323-327.
38. Teo, J., Lim, T. P, Hsu, L. Y, Tan, T. Y., Sasikala, S., Hon, P. Y., Kwa, A. L., & Apisarnthanarak A.
2015. Extensively drug-resistant Acinetobacter baumannii in a Thai hospital: a molecular
epidemiologic analysis and identification of bactericidal Polymyxin B-based combinations
Antimicrobial Resistance and Infection Control, 4(1), 2.
39. Wang, Y., Wu, C., Zhang, Q., Qi, J., Liu, H., Wang, Y., He, T., Ma, L., Lai, J., Shen, Z., Liu, Y., &
Shen, J. (2012). Identification of New Delhi metallo-β-lactamase 1 in Acinetobacter lwoffii
of food animal origin. PLoS One, 7(5), e37152.
40. Woodford, N., Ellington, M. J., Coelho, J. M., Turton, J. F., Ward, M. E., Brown, S., Amyes, S. G.,
& Livermore, D.M. (2006). Multiplex PCR for genes encoding prevalent OXA carbapenemases
in Acinetobacter spp. International Journal of Antimicrobial Agents, 27(4), 351-353.

111
41. Zarrilli, R., Pournaras, S., Giannouli, M., & Tsakris, A. (2013). Global evolution of multidrug-
resistant Acinetobacter baumannii clonal lineages. International Journal of Antimicrobial
Agents, 41(1), 11-19.
42. Zhao, S., Jiang, D., Xu, P., Zhang, Y., Shi, H., Cao, H., & Wu, Q. (2015). An investigation of drug-
resistant Acinetobacter baumannii infections in a comprehensive hospital of East China.
Annals of Clinical Microbiology and Antimicrobials, 14(7), 1-8.

112
บทท 4
การศกษาระบบควบคมสมดลคอปเปอรของ
เชอ A. baumannii
Manuscript preparation
Rapee Thummeepak, Aunchalee Thanwisai. Narissara Chantratita and Sutthirat Sitthisak. Molecular
characterization of copper resistance determinants of Acinetobacter baumannii.

113
บทน า (Introduction)
Acinetobacter baumannii เปนแบคทเรยทมการกระจายตวอยางกวางขวางในสงแวดลอมตามธรรมชาต
เชน ผวหนาของน าและดน (Baumann, 1968) นอกจากนยงเปนเชอกอโรคทพบไดบอยในชมชนและโรงพยาบาล
(Nosocomial pathogen) โดยมกพบการตดเชอไดบอยในปอด ระบบทางเดนปสสาวะ ระบบไหลเวยนโลหตและใน
ผปวยทเกยวของกบการใชเครองชวยหายใจ ในระยะ 10 ปทผานมามการระบาดของ A. baumannii เกดขนทวโลก
โดยเฉพาะสายพนธทดอตอยาปฏชวนะหลายชนด (Multidrug-resistant A. baumannii; MDR-AB) ซงดอตอกลมยา
carbapenems จงท าใหการเลอกใชยาปฏชวนะในการรกษาโรคตดเชอ A .baumannii มความซบซอนมากขน
(Rungruanghiranya et al., 2005) สงผลใหผปวยตดเชอมอตราการเสยชวตเพมมากขน นอกจากน A. baumannii ยง
มปจจยความรนแรงในการกอโรค (Virulence factor) ทหลากหลาย เชน สามารถสรางไบโอฟลม (Biofilm) ได ทนตอ
น ายาฆาเชอชนดตางๆได ท าใหเกดการแพรกระจายของเชอไดงาย
คอปเปอรเปนสารอาหารทจ าเปนตอการด ารงชวตส าหรบเซลลแบคทเรย แตตองการในปรมาณนอย
(Trace element) รปแบบของคอปเปอรทพบสวนมากม 2 รปแบบคอ Cu(II) ซงเปนรปแบบทใชรบอเลกตรอนจาก
สารตวอน (Oxidize form) และ Cu(I) ทเปนตวใหอเลกตรอนแกสารตวอน (Reduce form) โดยคอปเปอรทพบใน
ธรรมชาตสวนใหญจะอยในรปแบบ Cu(II) แตเซลลแบคทเรยจะน าคอปเปอรเขาสเซลลในรปแบบ Cu(I) (Solioz et al.,
2002; 2003) คอปเปอรเปนแรธาตทจ าเปนส าหรบการเจรญของแบคทเรย เพราะมบทบาทเปนโคแฟกเตอร (Cofactor)
ทส าคญของ redox enzyme เชน cytochrome c oxidase, lysyl oxidase, superoxide dismutase และยงม
บทบาทในกระบวนการ electron transport system ซงคอปเปอรจะเปนตวน าอเลกตรอนจาก cytochome c แลวสง
ตอใหออกซเจน เพอรดวซออกซเจนใหกลายเปนน า (Peña et al., 1999) แมวาคอปเปอรจะมความส าคญตอการ
เจรญเตบโตของแบคทเรย แตพบวาคอปเปอรในระดบเกนความจ าเปนจะท าใหเกดการเปนพษตอเซลลโดยเกดจาก
อนภาคของ reactive oxygen ทท าปฏกรยากบเซลลท าใหสวนประกอบของเซลลเสยหาย (Halliwell & Gutteridge,
1984) ซงความเปนพษของคอปเปอรถกน ามาใชในการเกษตรและการแพทยเพอใชเปนยาฆาเชอจลนทรยซงน าไปสการ
เลอกแบคทเรยสายพนธททนตอคอปเปอร (Dollwet & Sorenson, 1985)

114
ในรางกายมนษยจะใชโลหะหนกเพอควบคมการรกรานของแบคทเรยระหวางการตดเชอ โดยโฮสตจะจ ากด
ปรมาณโลหะหนกทจ าเปนเชนเหลก หรอสงกะสเพอลดและยบยงการรอดชวตของแบคทเรยซงเรยกสภาวะนวาการ
จ ากดโลหะ (metal limiting conditions) ในทางตรงขามกบสภาวะการจ ากดโลหะของ เหลก พบวาคอปเปอรสามารถ
ถายโอนเขาไปในบรเวณทตดเชอเพอก าจดแบคทเรยเรยกสภาวะนวาสภาวะทองแดงสง (copper excess conditions)
(Kyle & Eric, 2014) โดย Phagocytes สามารถถายโอนคอปเปอรเขาไปใน phagolysosome เพอฆาเชอแบคทเรยแต
แบคทเรยบางชนดกสามารถอยไดในสภาพแวดลอมคอปเปอรสงได (Kyle & Eric, 2014) มการศกษาทแสดงใหเหนวา
หากมการน ายนความตานทานตอคอปเปอรออกไปจากเซลลแบคทเรยกอโรค จะท าใหความรนแรงของการกอโรคลดลง
เชนสญเสยการสรางไบโอฟลม stress tolerance และ genetic transformation (Singh et al., 2015) สงเหลาน
ชใหเหนวาความตานทานคอปเปอรตอมความส าคญอยางยงส าหรบการรอดชวตของแบคทเรยทงในสภาพแวดลอมท
ปนเปอนคอปเปอร และเกยวของกบความรนแรงและการกอโรคของแบคทเรย
การศกษากอนหนานรายงานวาแบคทเรยจะมระบบควบคมสมดลคอปเปอรเพอชวยปองกนพษจากคอปเปอร
ท าใหแบคทเรยรอดชวตในสภาวะทมคอปเปอรสง ดวยกลไกทแตกตางกนหลายกลไกเชน efflux pump หรอ
detoxifying enzyme (Marie et al., 2012; Erik & Michael, 2015) ปจจปนกลไกการทนตอคอปเปอรไดรบการศกษา
โดยละเอยดในแบคทเรยแกรมลบ เชน Escherichia coli และแบคทเรยแกรมบวกเชน Enterococcus hirae (Rensing
& Grass, 2003; Solioz & Stoyanov, 2003) โดย E. coli มสองระบบทควบคมสมดลคอปเปอรเพอตอบสนองกบคอป
เปอรในความเขมขนทสง (Rensing & Grass, 2003) ระบบแรกคอ cue operon ซงประกอบดวยยน cueR, copA และ
cueO โดยมโปรตน CueR เปนตวควบคมทตรวจจบปรมาณ Cu + ใน periplasmic space ของเซลลซงน าไปสการชก
น าใหเกดการแสดงออกของยน copA และ cueO จากนนคอปเปอรในไซโทพลาซมจะถกขบออกส periplasmic space
โดยใช P-type ATPases (CopA) และมเอนไซม multi-copper oxidase (CueO) ท าหนาทในการเปลยน Cu+ ไปเปน
Cu2+ (มความเปนพษนอย) ใน periplasmic space และในสภาวะทม silver ions หรอไมมออกซเจน ระบบ CueO จะ
ไมท างานแตเซลลจะเปลยนมาใชระบบ cus operon ซงม sensor protein (CusS) ท าหนาทตรวจจบปรมาณ Cu + หรอ
silver ions ใน periplasmic space น าไปสการชกน าใหเกดการแสดงออกของยนและโปรตนควบคม regulator
protein (CusR) โดย CusR จะควบคมการแสดงออกของ cusCFBA (RND-efflux pump) เพอไปควบคมสมดลคอป
เปอรในเซลล ดงแสดงในภาพ 14 (Kinga & Zofia, 2013)

115
ภาพ 14 The mechanism of copper homeostasis in E. coli.
(ดดแปลงมาจาก Bondarczuk & Piotrowska-Seget, 2013)
ระบบควบคมสมดลคอปเปอรเปนหนงในกลไกทจ าเปนในเชอ A. baumannii และจากการศกษาขอมลจโนม
ของ A. baumannii สายพนธมาตราฐาน พบวาม copper resistance determinants เปนองคประกอบในโครโมโซม
(Williams et al., 2016; Hassan et al., 2017) จากการศกษาขอมลจโนมทงหมดของ A. baumannii หลายสายพนธ
ในฐานขอมล GeneBank (http://www.ncbi.nlm.nih.gov/genbank) พบวา A. baumannii มกลไกการควบคม
สมดลของคอปเปอรทงหมด 2 ระบบคอ cueR system และ copRS system ซงระบบ cueR จะพบในทกสายพนธแต
ระบบจะม copRS จะพบเพยงบางสายพนธ อยางไรกตามมการศกษานอยมากเกยวกบความชกของการม copper
resistance genes ในเชอ A. baumannii สายพนธทแยกไดจากผปวยและสายพนธทแยกจากธรรมชาตรวมทงศกษา
หนาทของ copper resistance genes ดงนนในการศกษาครงนจงมวตถประสงคเพอศกษารปแบบการควบคมสมดล
ของคอปเปอรของเชอ A. baumannii ในประเทศไทยทแยกไดจากผปวยและทแยกไดจากธรรมชาต ซงเรามงเนนไปท
การตรวจหาการแสดงออกของยนในแตละโอเพอรอนคอ cueR และ copRS gene การศกษาหนาทของ copper
resistance determinants ซงการศกษาครงนจะน ามาซงขอมลเบองตนเพอทน าไปศกษาถงบทบาท หนาทและการ
แสดงออกของแตละโอเพอรอน

116
วธด าเนนการวจย (Material and Method)
1. สายพนธแบคทเรยทใชในการศกษา
สายพนธแบคทเรยทใชในการศกษาครงน ไดแก จ านวน 102 ไอโซเลตโดยจดเปนเชอ A. baumannii ทแยกไดจาก
ผปวยโรงพยาบาลสวรรคประชารกษจงหวดนครสวรรค โรงพยาบาลเชยงรายประชานเคราะห จงหวดเชยงราย
โรงพยาบาลพจตร จงหวดพจตร ประเทศไทย และเชอ A. baumannii ทแยกไดจากทแยกจากแหลงน าธรรมชาต
ภายในจงหวดพษณโลก จ านวน 12 ไอโซเลต เชอ A. baumannii ทงหมดจะถกเพาะเลยงบนอาหาร Mueller-Hinton
หรอ Luria Bertani (LB) media (HiMedia) และน าไปบมทตบมอณหภม 37 องศาเซลเซยส
2. การศกษาคา Minimum inhibitory concentration (MIC) ของคอปเปอร
การศกษาคา Minimum inhibitory concentration (MIC) ของคอปเปอรในเชอ A. baumannii ทแยกไดจากผปวย
และแหลงน าธรรมชาตใชวธ agar dilution method (CLSI, 2014) โดยเตรยมอาหาร Mueller-Hinton agar plates
ทมสวนผสมของ copper sulfate ทความเขมขน 0, 0.25, 0.5, 1, 2, 4, 8, 12, 16, 24 และ 32 mM ตามล าดบ
จากนนเตรยมเชอแบคทเรยโดยเพาะเลยงเชอ A. baumannii แตละไอโซเลทในอาหาร Mueller-Hinton broth จนถง
exponential phase (ใชเวลาประมาณ 4-5 ชวโมง) จากนนปรบความเขมขนของเซลลแบคทเรยใหไดประมาณ 107
cells/ml น าจานอาหารเลยงเชอ Mueller-Hinton agar plates ทมสวนผสมของ copper sulfate ทความเขมขน
ตางๆมาหยดเชอทเตรยมไวปรมาตร 1 ไมโครลตร น าจานอาหารไปบมทตบมอณหภม 37 องศาเซลเซยสเวลาขามคน
จากนนสงเกตการเจรญของแบคทเรย
3. Bacterial growth curves
เตรยมเชอแบคทเรยโดยเพาะเลยงเชอ A. baumannii แตละไอโซเลทจากโคโลนเดยวขามคนในอาหาร Luria Bertani
(LB) media จากนนน า overnight culture ปรมาตร 1 มลลลตรเตมในอาหาร Mueller-Hinton broth ทมหรอไมม
CuSO4 ปรมาตร 100 มลลลตร 1:100 (v/v) จากนนน าไปบมท37 องศาเซลเซยสในตบมแบบเขยาทความเรวรอบ 150
rpm ตรวจวดคาการดดกลนแสงท 600 nm (OD600) ทก 1 ชวโมงเปนเวลา 15ชวโมง จากนนน าคา OD600 ไปสราง
growth curves

117
4. การตรวจหา copper associated genes
น าล าดบ amino acid sequence ของ copper resistance determinants ของแบคทเรยแกรมลบชนดตางๆทไดรบ
การศกษามากอนหนาน เชน E. coli และ Ps. auroginosa มาใชเปน template เพอน าไปเทยบหา copper
resistance proteins ใน databases ของเชอ Acinetobacter genomes โดยใชโปรแกรม Protein BLAST searches
(http://blast.ncbi.nlm.nih.gov/Blast.cgi) จากนนน า nucleotide sequences ของ copper associated genes
ท ไ ด ไ ป ใ ช เ ป น template ใ น ก า ร อ อ ก แ บ บ primers โ ด ย ใ ช online free-ware, Primer-BLAST
(https://www.ncbi.nlm.nih.gov/tools/primer-blast/) จากนนสกด genomic ของเชอ A. baumannii ทแยกได
จากผปวยและแหลงน าธรรมชาต โดยใช (Real Genomics DNA Extraction Kit, RBC Biosciences, Taiwan) เพอ
น ามาใชเปนtemplate ในการตรวจหา copper associated genes โดยวธ PCR ดวย specific primer ทออกแบบ
มาดงตาราง 22
ตาราง 22 Primers ทใชตรวจหา copper associated genes
Gene coding for: Primers Sequence (5'-3') Templates (GenBank accession no.)
Amplicon size (bp)
Annealing temperature
CueR regulator Full-cueR-F ATGAATATCGGTCAGGCA ABAYE2550 402 60 oC Full-cueR-R TTAACTTACGCCTACACCTT P-type ATPase1 F-in-ATPase CAACATTTGGTGGGAATGCA ABAYE2551 400 60 oC R-in-ATPase CAATTAGCATGACAGCCGGT PcoAB PcoAB-138F TCACATGAGTGCTGGAATGATGA ABUW_3228 to 3227 138 61 oC PcoAB-138R GAGAAAATAACTTAGTGATGCGCATG CopAB copAB-606F CCGTGCTCCAGAACGTGAAA ABAYE3201 to 3200 606 61 oC copAB-606R TTAGCCCACCAGGCTCTTTT CopRS copRS-312F GATTCTGCCTCGTGCTCTCA AB57_RS03385 to 03380 312 60 oC copRS-312R GCTGAGTACGGTCCTGAGTTT P-type ATPase2 ATPase2-992F CCCCATATGCGGTATGTCCC ABAYE3205 992 61 oC ATPase2-992R ATCCTAAAGCACACGGGCAA CopCD copCD-488F ACGCTCAACGTAAAGACATCCC AB57_RS03370 to 03365 488 60 oC copCD-488R ACTGTCCCACATCCCAGAAAG OprC oprC-851F GTTGCACGCCGTACATTGAA AB57_0206 851 60 oC oprC-851R GCAACAGCTCGCCATAACAG PitAB pitAB-151F ACAAACGCCTGAAGTTGTGC AUO97_12085 151 61 oC pitAB-151R ACGCTTAAACGTCGTTGTGC

118
5. Copper challenge
น าเชอ A. baumannii แตละไอโซเลทมาเพาะเลยงขามคนในอาหาร Mueller-Hinton broth จากนนน า 0.1 มลลลตร
ไปเตมในอาหาร Mueller-Hinton broth ปรมาตร 10 มลลลตร ( 1:100 (v/v) ) จากนนน าไปบมท37 องศาเซลเซยส
ในตบมแบบเขยาทความเรวรอบ 150 rpm จนถง exponential phase (ใชเวลาประมาณ 4-5 ชวโมง) จากนนน าเชอ
A. baumannii ทอยในชวง exponential phaseมาแบงใส Mueller-Hinton ปรมาตร 15 มลลลตร ทม CuSO4 ท
ความเขมขนตางๆกนบมทอณหภม 37 องศาเซลเซยสเปนเวลา 30 นาทจงเกบเซลลแบคทเรยทไดมาปนเหวยงและน า
เซลลไปสกด RNA โดยใช Total RNA Extraction Mini Kit (RBC Bioscience, Taiwan)
6. cDNA synthesis
น า RNA ทสกดไดจากขอ 5 มาวดความเขมขนโดยใชเครอง NanoDrop 1000 Spectrophotometer (Thermo
Scientific, USA) จากนนก าจด genomic DNA ทปนเปอนโดยน า1 µg ของ total RNA มา treated ดวย 1 unit of
DNase I (Thermo Scientific, USA) อางองวธท าตามคมอ จากนนน า DNase I-treated RNA samples ไปสงเคราะห
first-strand cDNA โดยใช Tetro cDNA Synthesis Kit (Bioline, UK) อางองวธท าตามคมอ
7. Quantitative Real-time PCR (qRT-PCR) analysis
การศกษาการแสดงออกของ copper associated genes เมอถกกระตนดวยโลหะหนกคอปเปอร ท าโดยใชวธ Real-
time qPCR assays ดวยเครอง Spectrum48 Real Time (Esco Technologies, USA) ท าการออกแบบ specific
primer pairs ส าหรบศกษา gene expression ดวยโปรแกรม Primer-BLAST software ดงแสดงในตารางท 23 ยน
gyrB จะถกเลอกเปนยน normalizing ส าหรบปฏกรยา qPCR ทงหมด (Henry et al., 2015) ปฏกรยา RT-qPCR จะ
ใชไพรเมอรแตละคและสภาวะในตาราง 23 โดยใช SYMB green PCR Master Mix (Sensifast SYBR No-Rox Kit,
Bioline, UK) ปฏกรยาทไมมเอนไซม transcriptase จะใชเปน negative control ตวอยางทงหมดจะถกทดสอบซ า 3
ครง
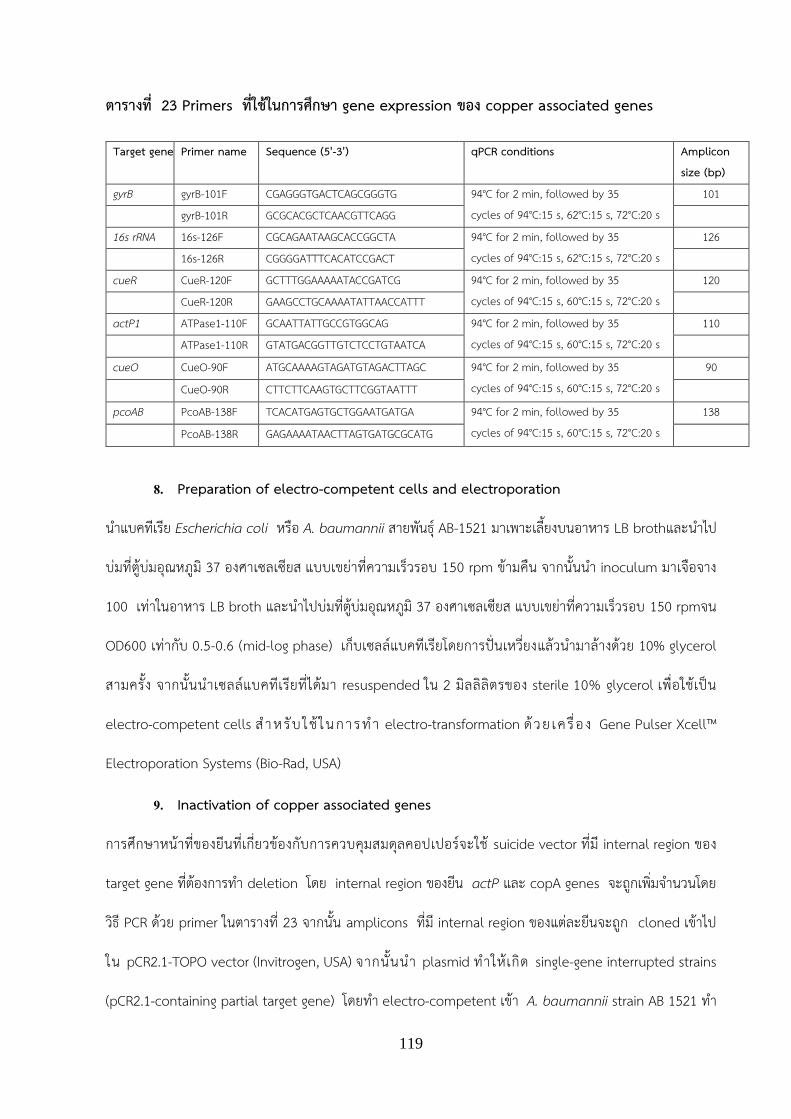
119
ตารางท 23 Primers ทใชในการศกษา gene expression ของ copper associated genes
8. Preparation of electro-competent cells and electroporation
น าแบคทเรย Escherichia coli หรอ A. baumannii สายพนธ AB-1521 มาเพาะเลยงบนอาหาร LB brothและน าไป
บมทตบมอณหภม 37 องศาเซลเซยส แบบเขยาทความเรวรอบ 150 rpm ขามคน จากนนน า inoculum มาเจอจาง
100 เทาในอาหาร LB broth และน าไปบมทตบมอณหภม 37 องศาเซลเซยส แบบเขยาทความเรวรอบ 150 rpmจน
OD600 เทากบ 0.5-0.6 (mid-log phase) เกบเซลลแบคทเรยโดยการปนเหวยงแลวน ามาลางดวย 10% glycerol
สามครง จากนนน าเซลลแบคทเรยทไดมา resuspended ใน 2 มลลลตรของ sterile 10% glycerol เพอใชเปน
electro-competent cells ส าหร บ ใช ในกา รท า electro-transformation ด วย เคร อ ง Gene Pulser Xcell™
Electroporation Systems (Bio-Rad, USA)
9. Inactivation of copper associated genes
การศกษาหนาทของยนทเกยวของกบการควบคมสมดลคอปเปอรจะใช suicide vector ทม internal region ของ
target gene ทตองการท า deletion โดย internal region ของยน actP และ copA genes จะถกเพมจ านวนโดย
วธ PCR ดวย primer ในตารางท 23 จากนน amplicons ทม internal region ของแตละยนจะถก cloned เขาไป
ใน pCR2.1-TOPO vector (Invitrogen, USA) จากนนน า plasmid ท าใหเกด single-gene interrupted strains
(pCR2.1-containing partial target gene) โดยท า electro-competent เขา A. baumannii strain AB 1521 ท า
Target genes Primer name Sequence (5'-3') qPCR conditions Amplicon size (bp)
gyrB gyrB-101F CGAGGGTGACTCAGCGGGTG 94°C for 2 min, followed by 35 cycles of 94°C:15 s, 62°C:15 s, 72°C:20 s
101 gyrB-101R GCGCACGCTCAACGTTCAGG 16s rRNA 16s-126F CGCAGAATAAGCACCGGCTA 94°C for 2 min, followed by 35
cycles of 94°C:15 s, 62°C:15 s, 72°C:20 s 126
16s-126R CGGGGATTTCACATCCGACT cueR CueR-120F GCTTTGGAAAAATACCGATCG 94°C for 2 min, followed by 35
cycles of 94°C:15 s, 60°C:15 s, 72°C:20 s 120
CueR-120R GAAGCCTGCAAAATATTAACCATTT actP1 ATPase1-110F GCAATTATTGCCGTGGCAG 94°C for 2 min, followed by 35
cycles of 94°C:15 s, 60°C:15 s, 72°C:20 s 110
ATPase1-110R GTATGACGGTTGTCTCCTGTAATCA
cueO CueO-90F ATGCAAAAGTAGATGTAGACTTAGC 94°C for 2 min, followed by 35 cycles of 94°C:15 s, 60°C:15 s, 72°C:20 s
90
CueO-90R CTTCTTCAAGTGCTTCGGTAATTT
pcoAB PcoAB-138F TCACATGAGTGCTGGAATGATGA 94°C for 2 min, followed by 35 cycles of 94°C:15 s, 60°C:15 s, 72°C:20 s
138
PcoAB-138R GAGAAAATAACTTAGTGATGCGCATG

120
การค ด เล อก transformed cells บนอาหาร LB agar ท ม 20 µg/ml of kanamycin. ตรวจหา single-gene
interrupted strains ของ actP1 และ copA โดยดการ integrated ของvector เขาไปในโครโมโซมโดยท า PCR
ดวย primers Pr-KanR-F และ Pr-KanR-R (ตาราง 24 )
ตาราง 24 Primers ทใชในการ Inactivation ของ copper associated genes
Target genes Primer name Sequence (5'-3') Templates (GenBank
accession no.)
Amplicon size (bp)
Annealing temperature
actP1 (P-type ATPase1)
F-in-ATPase CGGATCCCAACATTTGGTGGGAATGCA ABAYE2551
400 60oC
R-in-ATPase GCGAATTCCAATTAGCATGACAGCCGG
T
copA (MCO)
F-in-CopA GGAAGCTTTGGAATCAGATGCGAATGC ABAYE3201 300 60oC
R-in-CopA GCCGGATCCCTGATAATTTGCCTGTTT
KanR (pCR2.1 marker)
Pr-KanR-F GTAGAATTCCCGGAATTGCCAGC pCR2.1-TOPO (addgene)
930 60oC Pr-KanR-R GCCGAATTCGAAGAACTCGTCA

121
ผลการวจย (Result)
1. การตรวจหา copper associated genes โดยวธ PCR
Peptide sequences ของ copper resistance determinants จากเชอ E. coli และ Ps. auroginosa ถกใชมาเปน
templates ในการวเคราะห copper resistance determinants จาก genome database ของเชอ A. baumannii
พบวา whole-genome sequenced A. baumannii เกอบทกสายพนธม copper responsive operon ชนด cue
operon ขณะท copRS operon จะพบในบางสายพนธ (ภาพท 14) ผลจากการตรวจโดยใช primer ทจ าเพาะกบยน
ควบคม copper regulator ของทงสอง operon ไดแกยน cueR และ copR ผลของ PCR พบวา cueR ถกตรวจพบใน
ทกสายพนธทน ามาศกษาโดยพบใน 102 ไอโซเลทของเชอจากโรงพยาบาลและ 12 ไอโซเลทของเชอจากธรรมชาต
ขณะทยน copR พบเพยง 19 ไอโซเลท (18.63%) ของเชอจากโรงพยาบาล และ 2 ไอโซเลทของเชอจากธรรมชาต
(ตาราง 25) ผลการทดสอบความไวของเชอ A. baumannii ทงหมดตอคอปเปอรท าโดยวธ agar dilution ผลการศกษา
ใน 102 ไอโซเลทของเชอจากโรงพยาบาลพบมเพยง 25 ใน 102 ไอโซเลททใหคา MIC ทนตอคอปเปอรเทากบ 10 mM
ขณะทแบคทเรยสวนใหญทนคอปเปอรทคา MIC เทากบ 5 mM (75.5%) สวนเชอ A. baumannii สายพนธทแยกได
จากธรรมชาตจากเชอทม copper responsive operon ทง 2 ระบบจ านวน 2 isolates พบวามเพยง 1 ไอโซเลทมคา
MIC ตอCuSO4 มากกวาหรอเทากบ 10 µM เมอศกษาความสมพนธของรปแบบยนและความตานทานตอคอปเปอรถก
ศกษาโดยใชการทดสอบแบบ Fisher's exact test พบวา A. baumannii ทมรปแบบยนแบบ cueR + / copR +
(100%) ซงม copper responsive operon ของทงสองระบบทง 19 สายพนธมความตานทานตอคอปเปอรเทากบ 10
mM (100%) ขณะทสายพนธ cueR + / copR- ซงม copper responsive operon ระบบเดยวจากจ านวนทตรวจพบ
83 สายพนธมคาความตานทานตอคอปเปอรเทากบ 10 mM เพยง 6 สายพนธ (7.23%) แตเมอเปรยบเทยบรปแบบยน
กบคา MIC พบวาไมมความส าคญทางสถต (P <0.01)

122
ภาพท 14 ภาพแสดง copper resistance determinants ในเชอ A. baumannii รป operon แรกประกอบดวย 2 structural gene (ATPase: CueA and Chaperone: CueO) ซงถกควบคมโดย CueR รป และoperon ทสองพบในบางสายพนธของ A. baumannii ถกควบคมโดย CopRS
ตาราง 25 แสดงรปแบบของ copper responsive operon และความตานทานตอคอปเปอร (MIC) ของเชอ
clinical isolates 102 ไอโซเลท และเชอทแยกไดจากทแยกจากแหลงน าธรรมชาต 12 ไอโซเลท
Copper associated gene patterns
Total isolates n (%)
Isolates (%) 5 µM of MIC
CuSO4
Isolates (%) 10 µM of MIC
CuSO4 Clinical isolates n=102 (100) 77 (75.49) 25 (24.51) cueR+/copR+ 19 (100) 0 (0) 19 (100) cueR+/copR- 83 (100) 77 (92.77) 6 (7.23) Environmental isolates n=12 (100) 9 (75.00) 3 (25.00)
cueR+/copR+ 2 (100) 1 (50) 1 (50)
cueR+/copR- 10 (100) 8 (80) 2 (20)
2. ผลของคอปเปอรทมตอการเจรญของแบคทเรย
คดเลอก A. baumannii สองสายพนธไดแก สายพนธ AB1521 และ สายพนธ ATCC19606 ทม copper responsive
operon แตกตางกนมาศกษาผลของคอปเปอรทมตอการเจรญของแบคทเรยผลพบวา A. baumannii สายพนธ
AB1521 ทพบวามความตานทานตอคอปเปอรเทากบ 10 mM (MICcopper = 10 mM) และม copper responsive
operon ของทงสองระบบ ( cue and cop operons) สามารถทนและเจรญไดทความเขมขนคอปเปอร 1.25 mM
(ภาพท 15) ขณะท A. baumannii สายพนธ ATCC19606 ทพบวามความตานทานตอคอปเปอรเทากบ 5 mM

123
(MICcopper = 5 mM) และมเพยง cue operon สามารถทนและเจรญไดทความเขมขนคอปเปอร 0.625 mM (ภาพท
12)
ภาพ 15 ผลของคอปเปอรทมตอการเจรญของ A. baumannii สองสายพนธ
(A) Growth curves แสดงการเจรญของ A. baumannii AB1521 ทม copper responsive operon 2 operon
และ (B) Growth curves แสดงการเจรญของ A. baumannii reference strain ATCC19606 ทมแต cue
operons (B) ในสภาวะทมคอปเปอรความเขมขนตางๆ
3. การศกษาการแสดงออกของ copper resistance determinants ตอโลหะหนกคอปเปอร
การศกษารปแบบของ copper responsive operon และความตานทานตอคอปเปอร พบวา A. baumannii สาย
พนธ 1521 ทม copper responsive operon สองระบสวนใหญมคา MIC ทนตอคอปเปอรสง แสดงวา cop operon
มความเกยวของกบการแสดงออกของ copper resistance phenotype เพอยนยนสมมตฐานนจงมการทดลองการ
แสดงออกของ copper resistance determinants ทงใน cue และ cop operons ตอโลหะหนกคอปเปอร โดยวธ
RT-qPCR ผลพบวาเมอ A. baumannii สายพนธ AB1521 เจรญในสภาวะทม sub-MIC (0.0048 mM) ของคอป
เปอรมการชกน าการแสดงออกของยนใน cue operon ไดแก cueO และ actP1 (ATPase1) อยางไรกตามระดบการ

124
แสดงออกของยน copRS operon ไดแก copA copB actP2 (ATPase2) copC copD จะมการชกน าการแสดงออก
ของยนเมอ A. baumannii เจรญในสภาวะทมคอปเปอรสงขนทความเขมขน 0.625 mM เปนตนไป (ภาพ 16) และ
พบการยบยงการแสดงออกของยน porin encoding gene (oprC) ทงในสภาวะความเขมขนของคอปเปอรต า สวน
A. baumannii สายพนธ ATCC19606 มการชกน าการแสดงออกของยนใน cue operon เมอ A. baumannii
เจรญในสภาวะทมคอปเปอร ตงแต 0.024 mM นอกจากนพบมการแสดงออกของยน PcoAB ตอบสนองตอคอปเปอร
ใน A. baumannii ทงสองสายพนธ โดยไมพบการชกน าแสดงออกของยน ppk และ pitAB ใน A. baumannii ทง
สองสายพนธ
ภาพ 16 Heat map of expression profiles of presumtive genes in the present of various
concentration of copper of strain AB1521 (A) and ATCC19606 (B)

125
4. การศกษาหนาทของยนทเกยวของกบการควบคมสมดลคอปเปอร
การสราง copA deletion mutant (ยน encodeให Multicopper Oxidase จาก CopR operon) และ actP1
deletion mutant (ยน encodeให Copper ATPase1 จาก Cue operon) โดยท า PCR ยน copA สวนดานใน
(internal region) แลวโคลนเขา TA vector ซงเปน vector ทมคณสมบตเปน suicide vector ดงแสดงในภาพท 17
น า vector ทมยน copA เขาสเชอ A. baumannii สายพนธ AB 1521 และคดเลอกโคลน mutant บนอาหาร LB ทม
Kanamycin ซงไดเมอได mutant แลวน า copA deletion mutant และ actP1 deletion mutant ไปทดสอบคา
ความตานทานตอคอปเปอรเทยบกบ wild type ผลพบวา copA deletion mutant และ actP1 deletion mutant
จะมคาลดลง 2 เทา และ 4 เทาตามล าดบ (ตาราง 26)
ภาพ 17 แสดงขนตอนการท า mutation ยนทสนใจโดยใช suicide vector

126
ตาราง 26 คาความตานทานตอคอปเปอร (MIC) ของเชอ
Strains Characteristics MIC copper (mM)
AB1521 MDRAB, CRAB clinical isolate
10
AB1521ΔactP1 actP1 (efflux pump) interrupted derivative of AB1521, Kanr
2.5
AB1521ΔcopA copA (Multi-copper oxidase) interrupted derivative of AB1521, Kanr
5
* MDRAB: Multi-drug resistance A. baumannii, CRAB: Carbapenem resistance A. baumannii, Kanr:
Kanamycin resistance

127
ขอวจารณ (Discussion)
1. การตรวจหา copper resistance determinants ใน A. baumannii และการศกษาการแสดงออก
ของ copper associated genes ตอโลหะหนกคอปเปอร
ผล whole-genome sequenced ของ A. baumannii พบวาเชอ A. baumannii พบวา ม copper
responsive operon สองระบบ คอ cue operon และ copRS operon โดย cue operon ประกอบดวยยนท
encode โปรตน CueR โปรตน Copper ATPase และ heavy metal chaperon สวน copRS operon ประกอบดวย
ยนท encode โปรตน CopB โปรตน CopA โปรตน CopR โปรตน CopS โปรตน Copper ATPase โปรตน CopC
และโปรตน CopD (ภาพ 18) โดย Cue system เปนระบบทพบในแบคทเรยแกรมลบหลายสปชสเชน E. coli,
Pseudomonas aeruginosa, Pseudomonas putida, Pseudomonas fluorescens, Salmonella ( Sal.)
enterica และพบใน alphaproteobacteria เชน Agrobacterium tumefaciens และ Rhodobacter sphaeroides
(Rensing & Grass, 2003; Rademacher and Masepohl; 2012) ระบบนจะพบวามโปรตน CueR ซงจดเปน MerR
family of transcriptional activators ท าหนาทเปน copper-sensing transcriptional regulators (Rademacher
and Masepohl; 2012) สวนระบบทสองคอ copRS operon พบเปนระบบทใช two-component regulatory
system ทประกอบดวย CopR ท าหนาทเปน sensor kinase และ CopS ท าหนาทเปน response regulator ท า
หนาทรบสญญานจากภายนอกเซล เมอ CopS ถก activate โดยขบวนการ phosphorylation จะชกน าการแสดงออก
ของยนอนๆใน copRS operon โดยระบบนพบในแบคทเรยแกรมลบหลายสปชสเชน E. coli, P. fluorescens, P.
putida, Pseudomonas syringae และ Helicobacter pylori รวมทงแบคทเรยแกรมบวกเชน Corynebacterium
glutamicum (Cooksey, 1994; Thaden et al., 2010; Rademacher and Masepohl; 2012) ซงการศกษาใน E.
coli พบวา Cue system และ copRS operon (CusCFBA –like-systems) เปนระบบควบคมสมดลคอปเปอรใน
สภาวะ aerobic conditions ( Grass & Rensing, 2001 ; Outten et al., 2001 ) และ A. baumannii เปนแบคทเรย
ทเจรญในสภาวะมออกซเจนเทานน

128
ภาพ 18 ภาพแสดง copper responsive operon ในเชอ A. baumannii สองระบบ คอ cue operon
และ copRS operon
การตรวจหายนจาก copper responsive operon ไดแก cueR และ copR gene ดวยเทคนค PCR ในเชอ
A. baumannii ทแยกไดผปวยกบเชอทแยกจากธรรมชาต ผลการศกษาพบวาไมมความแตกตางกนของเชอทแยกได
ผปวยกบเชอทแยกจากธรรมชาต โดยพบยน cueR แสดงถงการม Cue system ในทกสายพนธของ A. baumannii สวน
การตรวจหา ยน copR แสดงถงการม CopRS system พบเพยง 18.6% ในเชอทแยกไดผปวยและ 16.6 % ในเชอทแยก
จากธรรมชาตสอดคลองกบขอมลใน GeneBank (http://www.ncbi.nlm.nih.gov/genbank) คอ cueR system เปน
ระบบทพบในเชอ A. baumannii เปนสวนมากซงตรวจพบทกสายพนธทมขอมลใน GeneBank แต copR system จะ
พบในบางสายพนธเทานน จากขอมลดงกลาวแสดงวา cueR system เปนระบบทมความจ าเปนในการควบคมสมดลของ
คอปเปอรภายในเซลลของเชอ A. baumannii แต copR system นนอาจเปนเพยงอกระบบทชวยใหเชอทนตอ
สภาวะแวดลอมทมคอปเปอรไดมากขน โดยอาจไดรบยนนมาจากแบคทเรยสปชสอนโดยขบวนการ horizontal gene
transfer

129
2. การศกษาการแสดงออกของ copper resistance determinants ตอโลหะหนกคอปเปอร
ผลการศกษาการแสดงออกของ copper resistance determinants ตอโลหะหนกคอปเปอรใน A. baumannii สาย
พนธ 1521 และ ATCC 19606 ทม copper responsive operon แบบ Cue system ชกน าการแสดงออกของยน
cueO และ actP1 โดยยน cueO จะ encode ให chaperon โปรตน เมอเปรยบเทยบกบยน cueO ในแบคทเรย
E.coli จะ encode ให periplasmic multicopper oxidase ซงท าหนาท oxidizes Cu (I) ไปเปน Cu (II) และ
reduces dioxygen ไปเปนน าผานขบวนการ four single-electron transfer steps (Djoko et al. 2010; Outten et
al. 2001; Roberts et al. 2002) สวนยน actP1 จะ encode ให ion-translocating ATPases ซงท าหนาทขนสง
monovalent Cu ออกจากไซโตรพลาสซม (Nies and Silver; 1995) นอกจากนพบมการแสดงออกของระบบ PcoAB
ตอบสนองตอคอปเปอรใน A. baumannii ทงสองสายพนธโดยยน PcoAB เปน (plasmid-borne copper resistance)
system ทพบครงแรกใน conjugative plasmid pRJ1004 ทแยกจาก E. coli ทอาศยอยในล าไสหมทเลยงดวยอาหาร
ทม copper sulfate (Bondarczuk & Piotrowska-Seget, 2013) แตในการศกษาใน A. baumannii พบวาระบบ
PcoAB นมคณสมบตเปน intrinsic gene พบในโครโมโซมของ A. baumannii ทกสายพนธโดย Choudhary และ
Sar ไดท าการศกษาการแสดงออกของ metal resistance genes ยน pcoA โดยวธ Real-time PCR ในเชอ P.
aeruginosa strain J007 พบวาการแสดงออกของยน copA จะมความส าคญในชวงทเชอสมผสกบคอปเปอรใน
ปรมาณต า ผลการศกษาการแสดงออกของ copper resistance determinants ตอโลหะหนกคอปเปอรใน A.
baumannii สายพนธ 1521 ทมเฉพาะ copper responsive operon แบบ copRS operon ชกน าการแสดงออกของ
ยน copA copB actP2 (ATPase2) copC copD ซงหนาทของยนเหลานยงไมทราบแนชดแตเมอเปรยบเทยบ Peptide
sequences ของ copper resistance determinants จากเชอ E.coli และ P. aeruginosa คาดวา copA จะท าหนาท
คลาย multicopper oxidase และ actP2 จะ encode ให ion-translocating ATPases
3. การศกษาหนาทของยนทเกยวของกบการควบคมสมดลคอปเปอร
การศกษาหนาทของยนทเกยวของกบการควบคมสมดลคอปเปอรเลอกศกษาสองยนคอยน copA
encode ให Multicopper Oxidase จาก CopRS operon และยน actP1 ยน encodeให copper ATPase จาก Cue
operon ผลคาความตานทานตอคอปเปอรพบวา copA deletion mutant และ actP1 deletion mutant มคา MIC

130
ลดลงเมอเทยบกบ wild type เชนเดยวกบการศกษากอนหนานเมอท าการ mutate ยน mco หรอยนท copA ท
encode copper ATPase ในแบคทเรยชนดตางๆ เชน E.coli หรอ Staphylococcus aureus จะพบวาแบคทเรยจะ
ม copper sensitivity ลดลง (Outten et al., 2001; Sitthisak et al., 2005; 2007)

131
สรปและขอเสนอแนะ (Conclusion and Recommendation)
สรปผลงานวจย
จากการศกษา whole genome sequence ของเชอ A. baumannii ใน GeneBank พบวามระบบควบคมสมดลของ
คอปเปอร (cop operon) ทงหมด 3 ระบบคอระบบ cueR system ระบบ copRS system และ pCo-homolog
system ซงแตละระบบจะควบคมการแสดงออกของยนทแตกตางกน จากการศกษาครงนผลพบวาเชอ A. baumannii
ทแยกจากผปวย และแยกไดจากธรรมชาต ตรวจพบระบบ cueR ทกสายพนธ (100%) พบระบบ copRS ในเชอทแยก
ไดผปวย 18.6% และ ในเชอทแยกจากธรรมชาต 16.6 % โดยสวนใหญเชอทมระบบควบคมสมดลของคอปเปอร (cop
operon) ทง 2 ระบบจะมคาความตานทานตอคอปเปอรสง (MIC เทากบ 10 mM) พบ 100% ในเชอทแยกไดผปวย
และ 50% ในเชอทแยกจากธรรมชาต การแสดงออกของ copper resistance determinants ในทงสองระบบถกชก
น าโดยโลหะคอปเปอรและเมอท าการ mutate ยน mco หรอยนท copA จะพบวาแบคทเรยจะม copper sensitivity
ลดลง
ขอเสนอแนะ
การศกษาครงนน ามาซงขอมลเบองตนเกยวกบบทบาท หนาทของระบบควบคมสมดลของคอปเปอรในเชอ A.
baumannii โดยควรมการศกษาเพมเตมเกยวกบหนาทของ copper resistance determinants เกยวของกบความ
รนแรงในการเกดโรคเชน biofilm formation และ การทนตอ oxidative stress ในเซล นอกจากนจากการทพบ cueR
system ในทกสายพนธของ A. baumannii อาจศกษาเพมเตมเพอหาสารทมคณสมบตยบยงการท างานของพบ cueR
system เพอน ามาใชเปน antimicrobial agent ตอไป

132
เอกสารอางอง (reference)
1. Baumann, P. (1968). Isolation of Acinetobacter baumannii from Soil and Water. Journal of
Bacteriology, 96(1), 39-42.
2. Bondarczuk, K. & Piotrowska-Seget, Z. (2013). Molecular basis of active copper resistance
mechanisms in Gram-negative bacteria. Cell Biology and Toxicology, 29(6), 397-405.
3. Cooksey, A. D. (1994). Molecular mechanisms of copper resistance and accumulation in
bacteria. FEMS Microbiology Reviews, 14(4), 381-6.
4. Djoko, K. Y., Chong, L. X., Wedd, A. G., & Xiao, Z. (2010). Reaction mechanisms of the
multicopper oxidase CueO from Escherichia coli support its functional role as a cuprous
oxidase. Journal of the American Chemical Society, 132(6), 2005–2015.
5. Dollwet, H. H. A., & Sorenson, J. R. J. (1985). Historic uses of copper compounds in medicine.
Trace Elements in Medicine, 2, 80-87.
6. Erik, L., & Michael, J. P. (2015). Copper tolerance and virulence in bacteria. Metallomics, 7,
957-964.
7. Grass, G., & Rensing, C. (2001). CueO is a multi-copper oxidase that confers copper tolerance
in Escherichia coli. Biochemical and Biophysical Research Communications, 286, 902-908.
8. Halliwell, B., & Gutteridge, J.M. (1984). Oxygen toxicity, oxygen radicals, transition metals and
disease. Biochemical Journal, 219, 1-14.
9. Hassan, K. A., Pederick, V. G., Elbourne, L. D., Paulsen, I. T., Paton, J. C., McDevitt, C. A., &
Eijkelkamp, B. A, (2017). Zinc stress induces copper depletion in Acinetobacter baumannii.
BMC Microbiology, 17, 59.
10. Henry, R., Crane, B., Powell, D., Deveson Lucas, D., Li, Z., Aranda, J., Harrison, P., Nation, R.L.,
Adler, B., Harper, M., Boyce, J.D., & Li, J. (2015). The transcriptomic response of

133
Acinetobacter baumannii to colistin and doripenem alone and in combination in an in vitro
pharmacokinetics/pharmacodynamics model. Journal of Antimicrobial Chemotherapy, 70,
1303-1313.
11. Kinga, B., & Zofia, P. S. (2013). Molecular basis of active copper resistance mechanisms in
Gram-negative bacteria. Cell Biology and Toxicology, 29, 397-405.
12. Kyle W. B., & Eric, P. S. (2014). Metal limitation and toxicity at the interface between host and
pathogen. FEMS Microbiology Reviews, 38, 1235-1249.
13. Marie, I. S., Chen, D., Dennis, J. T., & Darwin, K. H. (2012). Copper in Microbial Pathogenesis:
Meddling with the Metal. Cell Host & Microbe, 11, 106-115.
14. Nies, D.H., & Silver, S. (1955). Ion efflux systems involved in bacterial metal resistances.
Journal of Industrial Microbiology and Biotechnology, 14(2), 186-99.
15. Outten F. W., Huffman D. L., Hale J. A., & O’Halloran T. V. ( 2001). The independent cue and
cus systems confer copper tolerance during aerobic and anaerobic growth in Escherichia coli.
Journal of Biological Chemistry, 276, 30670–30677.
16. Peña, M. M., Lee, J., & Thiele, D. J. (1999). A delicate balance: homeostatic control of copper
uptake and distribution. The Journal of Nutrition, 129, 1251-1260.
17. Rademacher, C, & Masepohl, B. (2012) Copper-responsive gene regulation in bacteria.
Microbiology, 158(Pt 10), 2451-64.
18. Rensing, C., & Grass, G. (2003). Escherichia coli mechanisms of copper homeostasis in a
changing environment. FEMS Microbiology Reviews, 27, 197-213.
19. Roberts, S. A., Weichsel, A., Grass, G., Thakali, K., Hazzard, J. T., Tollin, G., Rensing, C, &
Montfort, W.R. (2002). Crystal structure and electron transfer kinetics of CueO, a multicopper
oxidase required for copper homeostasis in Escherichia coli. Proceedings of the National
Academy of Sciences, 99, 2766–71.

134
20. Rungruanghiranya, S., Somboonwit, C., & Kanchanapoom, T. (2005). Acinetobacter Infection in
the Intensive Care Unit. Journal of Infectious Diseases and Antimicrobial Agents, 22, 77-92.
21. Singh, K., Senadheera, D. B., Lévesque, C. M., & Cvitkovitch, D. G. (2015). The copYAZ operon
functions in copper efflux, biofilm formation, genetic transformation, and stress tolerance in
Streptococcus mutans. Journal of Bacteriology, 197, 2545-2557.
22. Sitthisak, S., Howieson, K., Amezola, C., & Jayaswal, R.K. (2005). Characterization of a
multicopper oxidase gene from Staphylococcus aureus. Applied and Environmental
Microbiology, 71(9), 5650-3.
23. Solioz, M., & Stoyanov, J. V. (2002). How cells deal with copper: a lesson from bacteria. In:
Anke, M., Mller, R., Schafer, U., Stoepple, M., eds. Macro and Trace Elements. pp. 485-492.
24. Solioz, M., Stoyanov, J. V. (2003). Copper homeostasis in Enterococcus hirae. FEMS
Microbiology Reviews, 27, 183–196.
25. Sitthisak S., Knutsson, L., Webb, J.W., & Jayaswal, R. K. (2007). Molecular characterization of
the copper transport system in Staphylococcus aureus, Microbiology, 153, 4274-83.
26. Thaden, J. T., Lory, S., & Gardner T. S. (2010). Quorum-sensing regulation of a copper toxicity
system in Pseudomonas aeruginosa. Journal of Bacteriology, 192, 2557–2568.
27. Williams, C. L., Neu, H. M., Gilbreath, J. J., Michel, S. L., Zurawski, D. V., & Merrell, D. S. (2016).
Copper resistance of the emerging pathogen Acinetobacter baumannii. Applied and
Environmental Microbiology, 82, 6174-6188.

135
กตตกรรมประกาศ
ผวจยขอขอบคณทนอดหนนการวจยส านกงานกองทนสนบสนนการวจย (สกว) ทใหการสนบสนนโครงการวจยนจนเสรจ
สนสมบรณ ผวจยขอขอบคณ นางสาวรววรรณ แสงศร นางสาวณฎฐน ลมเจยมรงษ นางสาวนารนทร ภกดโต นางสาว
กอบกล เทยนเทศ นางสาวกรพนภรณ มณนอย นางสาวพทยา พลวชย นายนครนทร ใจบญ นางสาวณฐชยา
บณฑตพฒนโภคน นสตปรญญาตร คณะวทยาศาสตรการแพทย มหาวทยาลยนเรศวร ทชวยในการท าวจย นางสาว
ภทรพร คงไทย นางสาวอดมลกษณ เหลองทองค า นสตปรญญาโท คณะวทยาศาสตรการแพทย มหาวทยาลย
นเรศวร นายระพ ธรรมมภกด นสตปรญญาเอก คณะวทยาศาสตรการแพทย ดร. อญชล ฐานวสย คณะวทยาศาสตร
การแพทย มหาวทยาลยนเรศวร รศ.ดร. นรศรา จนทราทตย ภาควชาจลชววทยาและอมมวโนโลย คณะ
เวชศาสตรเขตรอน มหาวทยาลยมหดล นายธวชชย กตต คณะแพทยตะวนออก วทยาลยเชยงราย และ Mr. Prabhu
Raj Joshi จาก Central Department of Microbiology, Tribhuvan University ทรวมด าเนนการวจย ผวจย
ขอขอบคณเจาหนาทจากโรงพยาบาลในประเทศไทยทงสโรงพยาบาลและโรงพยาบาล Tribhuvan University
ประเทศเนปาล ทใหความรวมมอในการเกบตวอยาง และสดทายนผวจยขอขอบคณ คณะวทยาศาสตรการแพทย
มหาวทยาลยนเรศวรและภาควชา จลชววทยาและปรสตวทยา คณะวทยาศาสตรการแพทย มหาวทยาลยนเรศวร ทให
การสนบสนนอปกรณเครองมอและสถานทในการท าวจย

136
Output ทไดจากโครงการ
1. เปนหวขอวทยานพนธของนสตบณทตศกษาระดบปรญญาโท 2 เรองและระดบปรญญาเอก 1 เรอง ไดแก
1.1 การศกษาคณสมบตทางอณชววทยาของคลาสด บตา-แลคทาเมส และการถายทอดยน ในเชอ
Acinetobacter baumannii
1.2 ลกษณะทางอณชววทยาของ extensively drug-resistant Acinetobacter baumannii ทแยกไดจาก
โรงพยาบาลในประเทศไทย
1.3 Molecular characterization of copper homeostasis systems in Acinetobacter baumannii
2. เขารวมประชมแสดงผลงานแบบ poster presentation ในงานประชมวชาการระดบชาตและนานาชาต
2.1 Rapee Thummeepak, Aunchalee Thanwisai, Narisara Chantratita and Sutthirat Sitthisak.
Characterization of copper resistance determinants among Acinetobacter baumannii clinical
isolates. The Asia Pacific Foundation for Infectious Diseases (APFID) และงาน ประชมวชาการ
ISAAR (International Symposium on Antimicrobial Agents and Resistance) & ICIC (International
Interscience Conference on Infection and Chemotherapy) ในวนท 14-16 กนยายน 2560 ณ เมองป
ซาน ประเทศเกาหล
2.2 Udomluk Leungtongkam, Rapee Thummeepak, Kannipa Tasanapak and Sutthirat Sitthisak.
Horizontal gene transfer of antibiotic resistance genes among Acinetobacter baumannii.
2.3 Phattaraporn Kongthai, Udomluk Leungtongkam, Rapee Thummeepak, Aunchalee
Thanwisai, Narisara Chantratita, Sutthirat Sitthisak. Molecular epidemiology of extensively-
drug resistance Acinetobacter baumannii (XDR-AB) isolated from four hospitals in Thailand.
น าเสนอผลงานทางวชาการ poster presentation ในการสมนาวทยาแหงชาต ครงท 23 ภายใตการประชม
วชาการเครอขายระดบโลกครงท 9 ระหวางวนท วนท 7-11 สงหาคม 2560 ณ โรงแรมดเอมเพลส จงหวด
เชยงใหม

137
3. ผลงานตพมพระดบนานาชาต 3 เรอง
3.1 Rapee Thummeepak, Phattaraporn Kongthai, Udomluk Leungtongkam, Sutthirat Sitthisak.
2016. Distribution of virulence genes involved in biofilm formation in multi-drug resistant
Acinetobacter baumannii clinical isolates. International Microbiology. 19(2):121-129.
doi:10.2436/20.1501.01.270
3.2 Prabhu Raj Joshi, Mahesh Acharya, Trishna Kakshapati, Udomluk Leungtongkam, Rapee
Thummeepak and Sutthirat Sitthisak. Co-existence of bla OXA-23 and bla NDM-1 genes
of Acinetobacter baumannii isolated from Nepal: antimicrobial resistance and clinical
significance. Antimicrobial Resistance & Infection Control. 2017; 6:21
3.3 Leungtongkam U, Thummeepak R, Wongprachan S, Thongsuk P, Kitti T, Ketwong K,
Runcharoen C, Chantratita N, Sitthisak S. Dissemination of blaOXA-23, blaOXA-24,
blaOXA-58, and blaNDM-1 Genes of Acinetobacter baumannii Isolates from Four Tertiary
Hospitals in Thailand. Microb Drug Resist. 2017 Jun 8. doi: 10.1089/mdr.2016.0248.
4. อยระหวางเตรยมผลงานตพมพระดบนานาชาตเพมอก 3 เรอง
4.1 Udomluk Leungtongkam, Rapee Thummeepak, Kannipa Tasanapak and Sutthirat Sitthisak.
Acquisition and transfer of antibiotic resistance genes and integrons of Acinetobacter
baumannii by conjugation.
4.2 Phattaraporn Kongthai, Udomluk Leungtongkam, Rapee Thummeepak, Thawatchai kitti ,
Aunchalee Thanwisai, Narisara Chantratita, Sutthirat Sitthisak. Molecular epidemiology and
virulence gene profiles of extensively drug-resistant Acinetobacter baumannii from Thailand.
4.3 Rapee Thummeepak, Aunchalee Thanwisai. Narissara Chantratita and Sutthirat Sitthisak.
Molecular characterization of copper resistance determinants of Acinetobacter baumannii.

138
ภาคผนวก
1. ผลงานตพมพ
2. การเชอมโยงกบตางประเทศ
2.1 เตรยมสงนสตบณทตศกษาระดบปรญญาเอกไปท าวจยท university of warwick ในหวขอ
Characterization of copper homeostasis genes of Acinetobacter baumannii.
2.2 ท างานวจยรวมกบนกวจยจากประเทศเนปาลเกยวกบ Acinetobacter baumannii
3. รางวลทไดรบ
OUTSTANDING ABSTRACT AWARDS จากงานประชมISAAR (International Symposium on
Antimicrobial Agents and Resistance) & ICIC (International Interscience Conference on
Infection and Chemotherapy) ในวนท 14-16 กนยายน 2560 ณ เมองปซาน ประเทศเกาหล

RESEARCH ARTICLEInternational Microbiology (2016) 19:121-129doi:10.2436/20.1501.01.270 ISSN (print): 1139-6709. e-ISSN: 1618-1095www.im.microbios.org
Distribution of virulence genes involvedin biofilm formation in multi-drug resistantAcinetobacter baumannii clinical isolates
Rapee Thummeepak,1 Phattaraporn Kongthai,1 Udomluk Leungtongkam,1 Sutthirat Sitthisak1,2
1Department of Microbiology and Parasitology, Faculty of Medical Sciences, Naresuan University, Phitsanulok, Thailand. 2Centre of Excellence in Medical Biotechnology, Faculty of Medical Science,
Naresuan University, Phitsanulok, Thailand
Received 27 April 2016 · Accepted 27 May 2016
Summary. Acinetobacter baumannii is an opportunistic bacterial pathogen that is the major cause of hospital-acquired infec-tions. It has been shown that A. baumannii with high biofilm formation increases the risk of acquiring infection. In this study, the prevalence of virulence genes involved in biofilm formation was determined in 225 A. baumannii clinical isolates from three hospitals in Thailand. Most of the isolates were multidrug-resistant A. baumannii strains (86.2%). Among all isolates, 76.9% (173/225) showed biofilm formation ability. The association between biofilm forming ability and gentamicin resistance was found (P < 0.05). The presence of virulence genes, epsA, bap, ompA, bfmS and blaPER-1 genes, was investigated by PCR. The prevalence of ompA, bfmS, bap, blaPER-1 and epsA genes among the isolated strains was 84.4%, 84%, 48%, 30.2%, respectively. Biofilm formation related genes, ompA and bap were associated with multidrug-resistant A. baumannii strains. The result of this study revealed that a high prevalence of biofilm-forming phenotypes among A. baumannii strains obtained from different hospitals. Effective strategies to prevent infection due to A. baumannii that produce biofilms are therefore needed. [Int Microbiol 19(2):121-129 (2016)]
Keywords: Acinetobacter baumannii · biofilms · virulence genes
*Corresponding author: Sutthirat SitthisakE-mail: [email protected]
Introduction
Acinetobacter baumannii is a Gram-negative bacterium that causes a variety of diseases. It exists especially in health care settings such as hospital environments. The emergence of multidrug-resistant A. baumannii strains is considered as a major and immediate threat to public health worldwide. One of the major factors involved in bacterial resistance to antimi-
crobials, chronic infections or survival in varying environ-ments is the ability to form biofilms. There are a variety of virulence determinants involved in biofilm formation of A. baumannii. This bacterium produces a molecule called the biofilm-associated protein (BAP), which is encoded by the bap gene [21]. BAP contributes to the initiation of biofilm production after A. baumannii attaches to a particular surface [14,19]. The outer membrane protein (OmpA), encoded by the ompA gene, is an adhesion molecule that functions during the attachment to human epithelial cells and induces biofilm formation [10]. Acinetobacter baumannii produces a polysac-charide export outer membrane protein, called exopolysac-

Int. MIcrobIol. Vol. 19, 2016 THUMMEEPAK ET AL.122
charide or EPS, which is encoded by epsA. EPS accumulates on the cell surface and provides protection to the cells against the harsh external environment [27,31]. Production of EPS is involved in the aggregation of bacteria which is associated with biofilm formation in many bacteria [33]. In addition, Lee et al. [16] have shown that the ability of clinical isolates of A. baumannii to form biofilm and to adhere to respiratory epithe-lial cells is enhanced by the presence and expression of the blaPER-1 gene. Recently, a two-component system (BfmS/BfmR) has been identified which is needed for biofilm forma-tion on polystyrene surfaces [32]. The A. baumannii 17978 type strain, with an inactivated bfmS, showed a reduction in biofilm formation [17]. To date, the mechanisms by which virulence determinants contribute to biofilm formation and antibiotic resistance still remain unclear. The aim of this study was to determine the association of biofilm formation, antibi-otic resistance phenotype and virulence genes in A. bauman-nii clinical isolates.
Materials and methods
Bacterial isolation and identification of Acinetobacter baumannii. A total of 225 individual clinical isolates was collected from 3 tertiary hospitals in 3 different provinces in Thailand over the 12 month period from November 2013 to October 2014. All A. baumannii isolates were col-lected from multiple collection sites, including sputum, urine, pus, blood, pleural fluid, ascetic fluid and wound. All isolates were identified as A. bau-
mannii by using biochemical tests, detecting of 16S rRNA gene [22] and the intrinsic blaOXA-51 gene [1,5]. Acinetobacter baumannii ATCC 19606 was used as a control.
Determination of antimicrobial susceptibilities. The antibiotic susceptibility was analyzed by the disc diffusion method (Oxoid). The concen-trations of the antibiotics in the discs (expressed in µg) were: amikacin (30), cefotaxime (30), ceftazidime (30), ceftriaxone (30), cefepime (30), ciprofloxa-cin (5), gentamicin (10), imipenem (10), meropenem (10), trimethoprim/sulfa-methoxazole (1.25/23.75), tetracycline (30), cefoperazone/sulbactam (105), and piperacillin/tazobactam (100/10). The Petri dishes were incubated at 35 °C for 24 h. The zones of inhibition determined whether the microorganism was susceptible, intermediately resistant, or resistant to each antibiotic. The results were interpreted according to the CLSI [6].
Detection of biofilm formation. Quantitative microtiter plate as-says for biofilm formation were performed as described by Brossard and Campagnari [4] with some modification. One hundred µl of 108 CFU/ml of A. baumannii and an equal volum of 2× Luria Bertain (LB) broth supple-mented with 20% glucose were added to each well in 96-well polystyrene microtiter plates (Nunc, Denmark). The plates were incubated overnight at 37 °C. After incubation, the cultures were gently removed. The wells were washed three times with phosphate buffered saline. The adherent cells were fixed with absolute methanol for 10 min and stained with 0.4 % crystal violet for 15 min, and washed three times with sterile distilled water and then air-dried. Afterward, the plates were filled with 250 μl of 33 % acetic acid for 15 min. The absorbance at OD595 nm was determined. All experiments were performed in three independent assays each repeated in triplicate. The mean optical density at 595 nm (OD595) of the non-biofilm producer E. coli DH5α was used as the OD cut-off value (ODc). The OD results of all tested strains were divided into the following four groups: (I) OD ≤ ODc = non biofilm Producer; (II) ODc < OD ≤ 2× ODc = weak biofilm Producer; (III) 2× ODc < OD ≤ 4 × ODc = moderate biofilm Producer; and (IV) 4× ODc < OD = strong biofilm producer [35].
Table 1. List of primers for detection of virulence genes used in this study
Target gene Primer sequence Tm (oC) References
epsA AGCAAGTGGTTATCCAATCGACCAGACTCACCCATTACAT 50 [31]
ompA CGCTTCTGCTGGTGCTGAATCGTGCAGTAGCGTTAGGGTA 50 [31]
blaPER-1
ATGAATGTCATTATAAAAGCAATTTGGGCTTAGGGCAAGAAA 50 [16]
bap TACTTCCAATCCAATGCTAGGGAGGGTACCAATGCAGTTATCCACTTCCAATGATCAGCAACCAAACCGCTAC 65 [12]
bfmS TTGCTCGAACTTCCAATTTATTATACTTATGCAGGTGCTTTTTTATTGGTC 53 [17]
16S rDNA AGAGTTTGATCCTGGCTCAGACGGCTACCTTGTTACGACTT 58 [22]
blaOXA-51
TAATGCTTTGATCGGCCTTGTGGATTGCACTTCATCTTGG 52 [5]

Int. MIcrobIol. Vol. 19, 2016 123VIRULENCE GENES OF A. BAUMANNII
Detection of virulence genes by PCR. The presence of epsA, bap, ompA, bfmS and blaPER-1 genes was detected with primers as shown in Table 1. DNA was extracted from all the isolates by boiling. Each PCR was performed in triplicate in a thermocycler with a PCR condition as described previously [12,16,17,31]. PCR products were analyzed by electrophoresis in 1% agarose gel containing 0.5 µg/ml ethidium bromide.
Statistical analyses. All statistical analyses were performed using Stata (Stata12.0 Corporation, USA). Non-parametric Kruskal–Wallis test and Dunn’s test were performed to compare the median value among multiple groups. The difference of biofilm biomass between two groups was compared by using Mann–Whitney U test. Fisher’s exact test was used to access differ-ences between frequencies. P-values < 0.05 were considered to be statisti-cally significant.
Results
Biofilm formation by clinical Acinetobacter baumannii isolates. All A. baumannii isolates were tested for the ability to form biofilms. The mean OD595 value for the negative control Escherichia coli DH5α was 0.221 ± 0.072 and this value was used as the optical density cut-off value (ODc). The classification of biofilm based on ODc re-
vealed that OD595 ≤ 0.221 = non-biofilm; 0.221 < OD595 ≤ 0.442 = weak biofilm; 0.442 < OD595 ≤ 0.884 = moderate bio-film; OD595 > 0.884 = strong biofilm. Among all isolates, 23.1% were non-biofilm producers, while the majority were biofilm producers (76.9%). The number of weak biofilm pro-ducers was 46 (20.4%), with 74 moderate biofilm producers (32.9 %) and 53 strong biofilm producers (23.6%). The me-dian OD595 and interquartile range (IQR) value for non-bio-film formers was 0.148 (0.091, 0.187), for weak biofilm form-ers, 0.332 (0.297, 0.404), moderate biofilm formers, 0.620 (0.494, 0.720) and strong biofilm formers, 1.170 (0.991, 1.430). We found that 173 of 225 isolates (76.9 %) were more capable of forming biofilms than the DH5α strain with a me-dian biofilm biomass of 0.624 (0.432, 0.949).
Distributions of biofilm-formers in various sources and wards. All 225 A. baumannii isolates were obtained from sputum (81.8 %), pus (7.5 %), urine (4.9 %) and other specimens (obtained from skin, blood, coccyx, cath-eter or pleural fluid) (5.8%). The proportion of strong biofilm producers of the other specimens was 46.2%, of urine, 45.5%,
Int M
icro
biol
Fig. 1. Characterization of biofilm production in 225 clinical isolates of Acinetobacter baumannii. (A and C) Relative composition of each biofilm formation level from different sources and wards. Each individual bar represents the proportion that contains different biofilm status. (B and D) The median of biofilm biomass (OD595) in clinical isolates from different sources and wards. Each data point is representative of the mean OD595 of independent triplicates of each individual isolates. The line bar (black) represents the median of OD595. ICU, Intensive Care Unit; MED, Medicine; SUR, Surgical; and Other: monk ward, coronary care unit, trauma ward, pediatric ward and outpatient department. Asterisks (*) indicate differences that are statistically significant; Kruskal–Wallis test followed by Dunn’s multiple comparison post-test; P < 0.05.

Int. MIcrobIol. Vol. 19, 2016 THUMMEEPAK ET AL.124
sputum, 32.6%, and pus, 11.8% (Fig. 1A). The comparison of biofilm biomass (OD595) among the various sources of speci-mens by the Kruskal–Wallis test showed significant differ-ences among the groups (P < 0.05). The median (IQR) of isolates obtained from urine [0.799 (0.489, 1.300)] and other specimens [0.720 (0.416, 1.330)] were higher than pus [0.186 (0.065, 0.423)] with P-values less than 0.05 (Dunn’s test) (Fig. 1B).
The prevalence of strong biofilm producers was similar among the strains recovered from different wards, ranging from 18.8% to 27.3% (Fig. 1C). However, analysis of the bio-film forming capacity of the isolates obtained from various wards revealed a statistically significant difference in OD595
between medical and surgical wards (P < 0.05; Kruskal–Wal-lis and Dunn’s tests) (Fig. 1D).
Association of biofilm-forming capability with antibiotic resistance phenotype. All isolates were tested for their antibiotic susceptibility toward 13 antibiotics. The majority of isolates were resistant to ciprofloxacin (84.4%). The A. baumannii isolated strains were also resistant to amikacin (54.2%), cefotaxime (76.9%), ceftazidime (82.2%), ceftriaxone (81.3%), cefepime (67.6%), gentamicin (68%), imipenem (79.6%), meropenem (78.7%), trime-thoprim/sulfamethoxazole (54.2%), tetracycline (62.7%) ce-foperazone/sulbactam (20.4%) and piperacillin/tazobactam (79.1%). All isolates were defined as being multidrug resis-tant A. baumannii (MDRAB) when there was resistance to
more than three antibiotic classes. The incidence of MDRAB was 86.2 % (194/225). Among 194 of MDRAB isolates, 150 (77.3 %) were biofilm-forming strains while 74.2 % (23/31) of non-MDRAB also produced biofilms (P = 0.654; Fisher’s exact test). This finding was supported by analysis of the me-dian OD595 among three drug resistance patterns (Fig. 2). As illustrated in Fig. 2B, the median (IQR) of non-MDRAB, MDRAB and XDRAB were 0.500 (0.157, 0.990), 0.461 (0.245, 0.794) and 0.605 (0.444, 0.770), respectively. There was no significant difference among the groups (P = 0.536; Kruskal–Wallis test).
The association between biofilm forming ability and indi-vidual drug resistance of A. baumannii was evaluated. The resistance rates of most antibiotics were found to be similar in both biofilm-forming and non-biofilm forming groups with a P-value ranging from 0.191 to 1.000. Of the 153 gentamicin resis tant isolates, 125 (81.7%) strains were biofilm producers while only 48 of 72 (66.7%) of gentamicin susceptible strains were biofilm producers (P = 0.017; Fish er’s exact test). This result was also confirmed by using Mann–Whitney U-test to compare the me dian (IQR) of OD595 between drug resistant and susceptible groups. In A. baumannii that resistance to gentamicin had a significantly higher ability to build biofilms when compared with gentamicin sensitive groups (P < 0.05) (Table 2). In contrast, the tetracycline susceptible isolates tended to form greater biofilm biomass than resistant strains (P < 0.001; Mann–Whitney U-test) (Table 2). However, the incidence rate of biofilm former in tetracycline susceptible
Int M
icro
biol
Fig. 2. Biofilm produced (OD595) on polystyrene by 225 isolates of Acinetobacter baumannii with different drug susceptibility patterns. (A) Biofilm formation of non MDR, MDR and XDR strains are represented by blue, green and orang bars, respectively. Data shown are the means of a triplicate plus standard deviation. (B) The open circle dot represents the mean of independent triplicate of each isolate. The line bar (black) indicates the median OD595 of each group with differences antibiotic resistance pattern. The dashed lines correspond to the cut-off value (ODc).

Int. MIcrobIol. Vol. 19, 2016 125VIRULENCE GENES OF A. BAUMANNII
strains was similar to that in resistant strains (82.1 vs. 73.8%) (P = 0.191; Fish er’s exact test). For other antibiotics, no sta-tistical correlation was observed (Table 2).
Relationship of biofilm production and the presence of different virulence genes. Polymerase chain reaction (PCR) was utilized to investigate the presence of the epsA, bap, ompA, bfmS and blaPER-1 genes in all A. bauman-nii isolates. The electrophoresis analysis showed that the ampli-con sizes of epsA, bap, ompA, bfmS and blaPER-1 genes were 451, 531, 927, 1225 and 1428 bp, respectively (data not shown). The most common virulence genes identified were ompA (84.4%) and bfmS (84%). The prevalence of genes bap, blaPER-1 and epsA genes among the isolated strains was 48%, 30.2% and 22.2%, respectively. Among the 225 A. baumannii isolates, all 5 virulence genes were present in 9 isolates (4%). The associa-tion between biofilm formed on the microtiter plate and the presence of virulence genes was also tested, using Mann–Whit-ney U test. The presence or absence of epsA, bfmS and blaPER-1 was not associated with the biofilm biomass (P > 0.05; Fig. 3C–E). The strains lacking bap or ompA genes form stronger biofilms than isolates carrying bap or ompA (P < 0.05; Fig. 3A,B). However, the frequency of epsA, bfmS, blaPER-1, bap and ompA was no significant difference between biofilm pro-ducers and non-biofilm producers with a P-value more than 0.05 (0.253, 0.281, 0.393, 0.117 and 0.199, respectively, Fish-er's exact test). We also examined the differences among vari-ous virulence gene patterns in their ability to produce biofilm. The number of virulence genes contributed to the trend of de-creased biofilm biomass, but this was not statistically signifi-cant (P > 0.05; Kruskal–Wallis test) (Fig. 3F).
Correlation between virulence genes and anti-biotic resistance patterns. The association between the presence of virulence genes and MDR status was evalu-ated. There is no statistical relationship between MDRAB and any of A. baumannii harbored epsA, bfmS and blaPER-1 genes (Table 3). The genes encoding BAP were present at a higher frequency in MDRAB than in non-MDRAB strains (P < 0.05) (Table 3). Gene ompA was present in 169 of 194 (87.1%) MDRAB isolates versus only 21 of 31 (67.74%) of non-MDRAB strains (P < 0.05). The correlation between the pres-ence of ompA and resistance to thirteen antimicrobials was also evaluated. The strains carrying ompA were found at a higher prevalence of resistance at least one drug from five antimicronial categories including aminoglycosides, ce-phems, fluoroquinolones, carbapenems and penicillins + β-lactamase inhibitors than the strains without this gene (P < 0.05;
(Table 4). The number of virulence genes present in the iso-lates was a statistically significant predictor of multiple drug resistance phenotype (risk ratio, 1.16; 95% CI, 1.11 to 2.42; P = 0.011).
Discussion
Biofilm production in clinical Acinetobacter baumannii. The ability of bacteria to form biofilm is regard-ed as an important virulence factor which plays a significant role in the bacteria’s persistence and antibiotic resistance [20]. In our work, we determined the biofilm formation, antibiotic suscepti-bility patterns and virulence genes among 225 clinical isolates. We found that more than seventy percent of studied A. bauman-nii showed biofilm formation ability. Our findings agree closely with those previously reported in [8] which showed that 75% of clinical A. baumannii isolates were positive for biofilm produc-tion, although different criteria were used to interpret biofilm status.
Correlation of biofilm among specimen types and wards. Clinical isolates recovered from urine and other sources (skin tissue, blood, coccyx, catheter and pleural fluids) were shown to have a significantly higher ability to form biofilms compared to those recovered from a pus source. This result agrees with a previous study which re-ported that A. baumannii obtained from urine specimens pro-duced biofilms with a greater biomass [8]. Urinary tract pathogens may have abilities to adhere and form biofilms in flowing environments, resulting in persistent infections. Our analysis also found significant differences in biofilm produc-ing capacity among isolates from various hospital wards. The ability to form a biofilm on an abiotic of clinical isolates pro-vides biofilm associated infection due to the attachment and colonization on medical device surfaces, such as urinary catheters [7].
Biofilm and antibiotic resistance. Acinetobacter baumannii is a major global health problem. In the past de-cade, high prevalence rates of MDRAB clinical isolates have been reported worldwide, ranging from 21–95% [15,18,34]. Similarly to other reports, we observed a high prevalence rate of MDRAB in this study. Previous studies reported that the MDR phenotype of pathogens as well as A. baumannii was linked to biofilm producing ability [13,28]. In contrast, our results indicate that the MDR and XDR phenotype has no as-sociation with biofilm producing ability. The ability of bacte-

Int. MIcrobIol. Vol. 19, 2016 THUMMEEPAK ET AL.126
ria to form biofilm may be associated with antibiotic resis-tance at the level of the individual. For example, Naparstek et al. [23] studied the biofilm production in Klebsiella pneu-moniae and they concluded that high-level gentamicin resis-tant strains show greater biofilm biomass compared with populations which have low-level resistance (median value of 0.15 versus 0.07, respectively) [23]. In 2016, Duarte et al. ob-
served that A. baumannii isolates resistant to gentamicin and tobramycin were more frequently able to form biofilms than susceptible strains [8]. In our study we found that strains pos-itive for biofilm formation were more frequently resistant to gentamicin.
In addition, the biofilm biomass of gentamicin resistance isolates was greater than susceptible groups (Table 2). Similar
Table 3. Relationship between virulence genes and antibiotic susceptibility patterns in all tested Acinetobacter baumannii isolates
The present of virulence genesAll A. baumannii isolates n = 225 (%)
P-valueaNon MDRAB (n = 31) MDRAB (n = 194)
bap 8 (25.81) 100 (51.55) 0.011*
ompA 21 (67.74) 169 (87.11) 0.013*
epsA 7 (22.58) 43 (22.16) 1.000
bfmS 26 (83.87) 163 (84.02) 1.000
blaPER-1 8 (25.81) 60 (30.93) 0.676aP-values represent the comparison between non MDRAB and MDRAB groups (Fisher’s exact test; P < 0.05). An asterisk (*) indicates the statistical significance (P-value < 0.05).
Table 2. Correlation between biofilm biomass (OD595) and drug resistance phenotype in all Acinetobacter baumannii isolates
Antibiotics susceptibilityMedian (IQR) of OD595
P-valueaSusceptible Resistant
amikacin 0.440 (0.252, 0.790) 0.496 (0.246, 0.890) 0.208
cefotaxime 0.481 (0.209, 0.921) 0.470 (0.277, 0.790) 0.942
ceftazidime 0.494 (0.211, 0.980) 0.470 (0.248, 0.790) 0.651
ceftriaxone 0.531 (0.252, 0.970) 0.465 (0.246, 0.790) 0.386
cefepime 0.515 (0.210, 0.830) 0.463 (0.258, 0.803) 0.893
ciprofloxacin 0.489 (0.157, 0.970) 0.471 (0.252, 0.790) 0.926
gentamicin 0.400 (0.154, 0.769) 0.515 (0.297, 0.860) 0.012*
imipenem 0.541 (0.279, 0.900) 0.461 (0.241, 0.790) 0.386
meropenem 0.510 (0.278, 0.860) 0.465 (0.241, 0.799) 0.648
TMX/SXT 0.521 (0.297, 0.901) 0.445 (0.237, 0.720) 0.166
tetracycline 0.692 (0.348, 1.055) 0.444 (0.213, 0.669) <0.001*
cefoperazone/sulbactam 0.461 (0.244, 0.814) 0.553 (0.268, 0.890) 0.303
piperacilin/tazobactam 0.462 (0.252, 0.790) 0.473 (0.246, 0.820) 0.928
aP-values represent the comparison of median OD595 of bacterial strains between two groups (Mann–Whitney U-test). An asterisk (*) indicates the significance (P-value < 0.05).

Int. MIcrobIol. Vol. 19, 2016 127VIRULENCE GENES OF A. BAUMANNII
results have also been reported for Pseudomonas aeruginosa, where strains characterized as gentamicin resistant showed a significant increase in biofilm production when compared to susceptible strains [30]. Our analysis of biofilm forming abil-ity with gentamicin resistance phenotypes has provided posi-tive statistical association findings.
We proposed that this phenotypic correlation may be due to biofilm-associated and gentamicin resistance determinants are co-located on the same plasmid or genomic island. How-ever, we found only an association between negative biofilm forming ability and tetracycline resistance phenotype. This finding differs from previous study in which a negative cor-relation between biofilm forming ability and antibiotic resis-tance to each of 20 antibiotics was reported [24]. The mecha-nism of this association was not clear but the expression of blaTEM-1 was reported to block biofilm formation via the bacte-rial adhesion interfering [11].
Biofilm and virulence genes. We found that A. bau-mannii isolates harboring virulence genes did not promote
biofilm forming ability on polystyrene, while the presence of bap or ompA showed an inverse correlation. Although many reports have demonstrated that biofilm associated genes, in-cluding bap, ompA, epsA, bfmS and blaPER-1, were responsible for the biofilm development of only certain selected A. bau-mannii strains [10,14,17,19,21,27,31,32], these reports did not fully characterize their functions in a diverse range of other strains and on different surfaces. Moreover, blaPER-1 and ompA were not over-expressed in biofilm cells as previously analyzed indicating that these genes are not fully required for biofilm production in some strains [25,26]. This suggests that other key factors or strain-dependent variations contribute to biofilm forming phenotypes in diverse biotic or abiotic sur-faces. [3,9].
Virulence gene and MDR phenotype. Although, the OmpA of A. baumannii was found to be essential for the development of biofilms and attachment to human epithelial cells [10]. Its involvement in antimicrobial resistance pheno-type was also reported [29]. In agreement with our results, the
Int M
icro
biol
Fig. 3. The correlation between in vitro biofilm formation and virulence genes. (A–E) Biofilm forming capacity (OD595) of bacteria harboring and not harboring individual virulence genes. Asterisks (*) indicate differences that are statistically significant; Mann–Whitney U-test. (F) OD595 among various virulence gene patterns P0; the absent of gene and P1–P5; the present of one to five genes, respectively. Each data point is the mean OD595 of independent triplicates of each individual isolates. The line bar (black) represents the median OD595 of each group.

Int. MIcrobIol. Vol. 19, 2016 THUMMEEPAK ET AL.128
association between MDR phenotype and the presence of bap and ompA genes was found. It is possible that due to OmpA being a β-barrel porin, antibiotics may be transferred from the periplasm through the outer membrane and then couples with inner membrane efflux pumps. We also found that A. bauman-nii carrying ompA were associated with individual drug resis-tant phenotype (e.g., cefotaxime, ciprofloxacin, and imipen-em). This finding is consistent with a previous report which indicated that, in A. baumannii ATCC 17978, OmpA was in-volved in resistance to chloramphenicol, aztreonam and nali-dixic acid [29].
We conclude that in this study we found a high prevalence of MDRAB and no difference in its ability to form biofilms when compared with non-MDRAB. Our data indicate that non-MDRAB strains have the ability to form biofilms, and biofilm formation might help these strains adapt or persist
during infections. The presence of tested virulence genes does not seem to be related to biofilm formation of A. baumannii on a plastic surface. Interestingly, two of those genes, espe-cially ompA, was associated with antibiotic resistant pheno-types. The transcriptional or translational analysis of viru-lence genes can provide good data to confirm their association with biofilm or antibiotic resistance phenotypes which must be further analyzed.
Acknowledgements. Our thanks to Roy Morien of the Naresuan Uni-versity Language Centre for his editing assistance and advice on English ex-pression in this document. This work has been supported by grants from the Thailand Research Fund and Naresuan University (RSA5780015) to S. Sit-thisak and The Royal Golden Jubilee PhD Program to R. Thummeepak (PHD/ 0031/2558).
Competing interests. None declared.
Table 4. The correlation between the presence of ompA and antibiotic resistance phenotype in all tested Acinetobacter baumannii isolates
Antibiotics resistanceAll A. baumannii isolates n = 225 (%)
P-value a
ompA– (n=35) ompA+ (n=190)
AMInoglycosIdes
amikacin 17 (48.57) 105 (55.26) 0.468
gentamicin 18 (51.43) 135 (71.05) 0.030*
cepheMs
cefotaxime 22 (62.56) 151 (79.47) 0.048*
ceftazidime 23 (65.71) 162 (85.26) 0.014*
ceftriaxone 22 (62.57) 161 (84.74) 0.004*
cefepime 21 (60.00) 131 (68.95) 0.328
FluoroquInolones
ciprofloxacin 25 (71.43) 165 (86.84) 0.038*
carbapenems
imipenem 20 (57.14) 159 (83.68) 0.001*
meropenem 21 (60.00) 156 (82.11) 0.006*
FolAte pAthwAy InhIbItors
TMX/SXT 19 (54.29) 103 (54.21) 1.000
penIcIllIns + β-lactamase inhibitors
cefoperazone/sulbactam 2 (5.71) 44 (23.16) 0.021*
piperacilin/tazobactam 23 (65.71) 155 (81.58) 0.042*
aP-values were analyzed using Fisher's exact test. Asterisks (*) assign statistical significance results (P < 0.05) between the ompA-negative and the ompA-positive groups.

Int. MIcrobIol. Vol. 19, 2016 129VIRULENCE GENES OF A. BAUMANNII
References
1. Akers KS, Chaney C, Barsoumian A, et al. (2010) Aminoglycoside resis-tance and susceptibility testing errors in Acinetobacter baumannii-calco-aceticus complex. J Clin Microbiol 48:1132-1138
2. Badave GK, Kulkarni D (2015) Biofilm producing multidrug resistant Acinetobacter baumannii: An emerging challenge. J Clin Diagn Res 9:DC08-10
3. Bitrian M, Solari CM, González RH, Nudel CB (2012) Identification of virulence markers in clinically relevant strains of Acinetobacter geno-species. Int Microbiol 15:79-88
4. Brossard KA, Campagnari AA (2012) The Acinetobacter baumannii biofilm-associated protein plays a role in adherence to human epithelial cells. Infect Immun 80:228-233
5. Brown S, Young HK, Amyes SG (2005) Characterization of OXA-51, a novel class D carbapenemase found in genetically unrelated clinical strains of Acinetobacter baumannii from Argentina. Clin Microbiol In-fect 11:15-23
6. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-first Informational Supple-ment M100-S21. CLSI, Wayne, PA, USA, 2011
7. Djeribi R, Bouchloukh W, Jouenne T, Menaa B (2012) Characterization of bacterial biofilms formed on urinary catheters. Am J Infect Control 40:854-859
8. Duarte A, Ferreira S, Almeida S, Domingues FC (2016) Clinical isolates of Acinetobacter baumannii from a Portuguese hospital: PFGE charac-terization, antibiotic susceptibility and biofilm-forming ability. Comp Immunol Microbiol Infect Dis 45:29-33
9. Eijkelkamp BA, Stroeher UH, Hassan KA, Elbourne LD, Paulsen IT, Brown MH (2013) H-NS plays a role in expression of Acinetobacter baumannii virulence features. Infect Immun 81:2574-2583
10. Gaddy JA, Tomaras AP, Actis LA (2009) The Acinetobacter baumannii 19606 OmpA protein plays a role in biofilm formation on abiotic sur-faces and in the interaction of this pathogen with eukaryotic cells. Infect Immun 77:3150-3160
11. Gallant CV, Daniels C, Leung JM, Ghosh AS, Young KD, Kotra LP, Bur-rows LL (2005) Common beta-lactamases inhibit bacterial biofilm for-mation. Mol Microbiol 58:1012-1024
12. Goh HMS, Beatson SA, Totsika M, et al. (2013). Molecular analysis of the Acinetobacter baumannii biofilm-associated protein. Appl Environ Microbiol l79:6535-6543
13. Gurung J, Khyriem AB, Banik A, Lyngdoh WV, Choudhury B, Bhat-tacharyya P (2013) Association of biofilm production with multidrug re-sistance among clinical isolates of Acinetobacter baumannii and Pseudo-monas aeruginosa from intensive care unit. Indian J Crit Care Med 17:214-218
14. Howard A, O’Donoghue M, Feeney A, Sleator RD (2012) Acinetobacter baumannii: An emerging opportunistic pathogen. Virulence 3:243-250
15. Inchai J, Liwsrisakun C, Theerakittikul T, Chaiwarith R, Khositsakulchai W, Pothirat C (2015) Risk factors of multidrug-resistant, extensively drug-resistant and pandrug-resistant Acinetobacter baumannii ventila-tor-associated pneumonia in a Medical Intensive Care Unit of University Hospital in Thailand. J Infect Chemother 21:570-574
16. Lee HW, Koh YM, Kim J, et al. (2008) Capacity of multidrug-resistant clinical isolates of Acinetobacter baumannii to form biofilm and adhere to epithelial cell surfaces. Clin Microbiol Infect 14:49-54
17. Liou ML, Soo PC, Ling SR, Kuo HY, Tang CY, Chang KC (2014) The sensor kinase BfmS mediates virulence in Acinetobacter baumannii. J Microbiol Immunol Infect 47:275-281
18. Liu Q, Li W, Du X, et al. (2015) Risk and prognostic factors for multi-drug-resistant Acinetobacter baumannii complex bacteremia: A retro-
spective study in a tertiary hospital of west China. PLoS ONE 10:e0130701
19. Loehfelm TW, Luke NR, Campagnari AA (2008) Identification and char-acterization of an Acinetobacter baumannii biofilm-associated protein. J Bacteriol 190:1036-1044
20. Longo F, Vuotto C, Donelli G (2014) Biofilm formation in Acinetobacter baumannii. New Microbiol 37:119-127
21. McConnell MJ, Actis L, Pachón J (2013) Acinetobacter baumannii: hu-man infections, factors contributing to pathogenesis and animal models. FEMS Microbiol Rev 37:130-155
22. Misbah S, Hassan H, Yusof MY, Hanifah YA, AbuBakar S (2005). Ge-nomic species identification of Acinetobacter of clinical isolates by 16S rDNA sequencing. Singapore Med J 46:461-464
23. Naparstek L, Carmeli Y, Navon-Venezia S, Banin E (2014) Biofilm for-mation and susceptibility to gentamicin and colistin of extremely drug-resistant KPC-producing Klebsiella pneumoniae. J Antimicrob Che-mother 69:1027-1034
24. Qi L, Li H, Zhang C, et al. (2016) Relationship between antibiotic resis-tance, biofilm formation, and biofilm-specific resistance in Acinetobacter baumannii. Front Microbiol 7:483 doi:10.3389/fmicb.2016.00483
25. Rao RS, Karthika RU, Singh SP, et al. (2008) Correlation between bio-film production and multiple drug resistance in imipenem resistant clini-cal isolates of Acinetobacter baumannii. Indian J Med Microbiol 26: 333-337
26. Rumbo-Feal S, Gómez MJ, Gayoso C, et al. (2013) Whole transcrip-tome analysis of Acinetobacter baumannii assessed by RNA-sequencing reveals different mRNA expression profiles in biofilm compared to planktonic cells. PLoS One 8:e72968
27. Russo TA, Luke NR, Beanan JM, et al. (2010) The K1 capsular polysac-(2010) The K1 capsular polysac-charide of Acinetobacter baumannii strain 307-0294 is a major virulence factor. Infect Immun 78:3993-4000
28. Sanchez CJ Jr, Mende K, Beckius ML, Akers KS, Romano DR, Wenke JC, Murray CK (2013) Biofilm formation by clinical isolates and the implications in chronic infections. BMC Infect Dis 13:47
doi:10.1186/1471-2334-13-4729. Smani Y, Fàbrega A, Roca I, Sánchez-Encinales V, Vila J, Pachón J
(2014) Role of OmpA in the multidrug resistance phenotype of Acineto-bacter baumannii. Antimicrob Agents Chemother 58:1806-1808
30. Suman E, Varghese S, Jose J (2005) Gentamicin resistance in biofilm producing Pseudomonas aerruginosa causing catheter associated uri-nary tract infections. Indian J Med Sci 59:214-216
31. Tayabali AF, Nguyen KC, Shwed PS, Crosthwait J, Coleman G, Seligy VL (2012) Comparison of the virulence potential of Acinetobacter strains from clinical and environmental sources. PLoS One 7:e37024
32. Tomaras AP, Flagler MJ, Dorsey CW, Gaddy JA, Actis LA (2008) Char-acterization of a two component regulatory system from Acinetobacter baumannii that controls biofilm formation and cellular morphology. Mi-crobiology 154:3398-3409
33. Vu B, Chen M, Crawford R, Ivanova E (2009) Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 14:2535-2554
34. Wright MS, Iovleva A, Jacobs MR, Bonomo RA, Adams MD (2016) Genome dynamics of multidrug-resistant Acinetobacter baumannii dur-ing infection and treatment. Genome Med 8:1-12 doi:10.1186/s13073-016-0279-y
35. Zhang D, Xia J, Xu Y, Gong M, Zhou Y, Xie L, Fang X (2016) Biological features of biofilm-forming ability of Acinetobacter baumannii strains derived from 121 elderly patients with hospital-acquired pneumonia. Clin Exp Med 16:73-80

RESEARCH Open Access
Co-existence of blaOXA-23 and blaNDM-1genes of Acinetobacter baumannii isolatedfrom Nepal: antimicrobial resistance andclinical significancePrabhu Raj Joshi1, Mahesh Acharya1, Trishna Kakshapati2, Udomluk Leungtongkam3, Rapee Thummeepak3
and Sutthirat Sitthisak3,4*
Abstract
Background: Molecular analysis of carbapenem-resistant genes in Acinetobacter baumannii, an emerging pathogen,is less commonly reported from Nepal. In this study we determined the antibiotic susceptibility profile and geneticmechanism of carbapenem resistance in clinical isolates of A. baumannii.
Methods: A. baumannii were isolated from various clinical specimens and identified based on Gram staining,biochemical tests, and PCR amplification of organism specific 16S rRNA and blaOXA-51 genes. The antibioticsusceptibility testing was performed using disc diffusion and E-test method. Multiplex PCR assays were used todetect the following β-lactamase genes: four class D carbapenem hydrolyzing oxacillinases (blaOXA-51, blaOXA-23,blaOXA-24 and blaOXA-58). Uniplex PCRs were used to detect three class B metallo-β-lactamases genes (blaIMP, blaVIMand blaNDM-1), class C cephalosporin resistance genes (blaADC), aminoglycoside resistance gene (aphA6), and ISAba1of all isolates. Insertion sequence ISAba125 among NDM-1 positive strains was detected. Clonal relatedness of allisolates were analyzed using repetitive sequence-based PCR (rep-PCR).
Results: Of total 44 analyzed isolates, 97.7% (n = 43) were carbapenem-resistant A. baumannii (CR-AB) and 97.7% (n = 43)were multidrug resistant A. baumannii (MDR-AB). One isolate was detected to be extremely drug resistant A. baumannii(XDR-AB). All the isolates were fully susceptible to colistin (MICs < 2 μg/ml). The blaOXA-23 gene was detected in all isolates,while blaNDM-1 was detected in 6 isolates (13.6%). Insertion sequence, ISAba1 was detected in all of blaOXA-23 positiveisolates. ISAba125 was detected in all blaNDM-1 positive strains. The blaADC and aphA6 genes were detected in 90.1 and 40.1%, respectively. The rep-PCR of all isolates represented 7 different genotypes.
Conclusion: We found high prevalence of CR-AB and MDR-AB with blaOXA-23 gene in a tertiary care hospital in Nepal.Systemic network surveillance should be established for monitoring and controlling the spread of these resistant strains.
Keywords: Acinetobacter baumannii, Carbapenem resistance, blaOXA-23 and blaNDM-1 carbapenemase genes
* Correspondence: [email protected] of Microbiology and Parasitology, Faculty of Medical Science,Naresuan University, Phitsanulok, Thailand4Centre of Excellence in Medical Biotechnology, Faculty of Medical Science,Naresuan University, Phitsanulok, ThailandFull list of author information is available at the end of the article
© The Author(s). 2017 Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, andreproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link tothe Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver(http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
Joshi et al. Antimicrobial Resistance and Infection Control (2017) 6:21 DOI 10.1186/s13756-017-0180-5

BackgroundAcinetobacter baumannii, an emerging pathogen ofhealthcare centers, shows intrinsic as well as acquireddrug-resistance mechanisms [1]. Multidrug-resistant A.baumannii can be resistant to all of the currently avail-able antibiotics, and in its deadliest form these are onlysusceptible to potentially toxic polymyxins and colistins,leaving limited options for treatment [2]. Infections withcarbapenem- and colistin-resistant A. baumannii areemerging globally [3].Carbapenem resistance in A. baumannii encompasses
production of class B, C and class D carbapenemase, de-creased membrane permeability, altered penicillin-binding proteins, and overexpression of efflux pumps [4,5]. Most commonly, Acinetobacter spp. develop carba-penem resistance by production of OXA-type carbape-nemase and metallo-β-lactamases (MBLs) [6, 7]; blaOXA-
23-like, blaOXA-40-like, blaOXA-58-like and blaOXA-51-likecarbapenemases are broadly reported, where blaOXA-51-like β-lactamases, intrinsic to A. baumannii, is used forspecies identification [8–10]. Among multiple MBLgenes, blaIMP and blaVIM types (chromosomal or plasmidencoded) encode carbapenemase in A. baumannii [9]. A.baumannii harboring plasmid encoded New Delhimetallo-β-lactamase-1 (NDM-1), a novel carbapenemasegene, is reported from many countries [11, 12]. Inaddition, detection of class C β-lactamase genes (blaADC)which mediated cephalosporin resistances and aminogly-coside resistant genes (aphA6) has increased in recentyears in A. baumannii clinical isolates [13, 14].A. baumannii remains a critical problem in many
healthcare settings throughout the world despite the imple-mentation of infection control practices. There are limiteddata on carbapenem-resistant A. baumannii in Nepal. Theobjective of this study was to determine antibiotic suscepti-bility profile, antibiotic resistance genes and genetic mech-anism of carbapenem resistance of A. baumannii in clinicalisolates at a tertiary care hospital, Nepal.
MethodsBacterial isolation and identificationA. baumannii isolates were collected from inpatientunits of a tertiary hospital, Nepal. Forty-four non-duplicate isolates were collected (24 male and 20 female;age range between 24 to 80 years) over 9 months periods(October 2014 to June 2015). All isolates were identifiedby classical biochemical methods and confirmed by PCRmethod for detecting 16S rRNA gene and blaOXA-51
gene [15, 16]. Isolates were identified as A. baumanniiby PCR result of positive for both PCRs.
Antibiotic susceptibility testingThe antibiotic susceptibility of amikacin (30), cefotax-ime (30), ceftazidime (30), ceftriaxone (30), cefepime
(30), ciprofloxacin (5), gentamicin (10), imipenem(10), meropenem (10), trimethoprim/sulfamethoxazole(1.25/23.75), tetracycline (30), and piperacillin/tazo-bactam (100/10) (Oxoid) was determined on MuellerHinton Agar (High Media, India) according to theantibiotic disk diffusion method [17]. The plates wereincubated at 37 °C for 24 h. The zones of inhibitionwere determined whether the microorganism was sus-ceptible, intermediately resistant, or resistant to eachantibiotic according to Clinical and LaboratoryStandards Institute (CLSI) guidelines. E-test was per-formed to determine the Minimum inhibitory concen-tration (MIC) of ceftazidime, imipenem, tigecyclineand colistin (High Media, India) according to manu-facturer instructions and interpreted as per CLSIguidelines except for tigecycline. Multidrung-resistantA. baumannii (MDR-AB) was defined when A. bau-mannii resistant to multiple antibiotics, often definedas three or more antibiotic classes. Extensively drugresistant A. baumannii (XDR-AB) was defined whenA. baumannii was resistant to all antimicrobial agentsexcept polymyxins (colistin) [18].
PCR amplification of antibiotic resistance genesPCR assays to detect blaOXA-23, blaOXA-24, blaOXA-51,blaOXA-58, blaIMP, blaVIM, blaNDM, blaADC and ahpA6genes were performed using primers as describe previ-ously (Table 1). The amplification reaction was per-formed using A. baumannii cell lysate as DNAtemplate. Each PCR was performed in triplicate in athermocycler with a PCR condition as described previ-ously [14, 16, 19–21]. All PCR assays used 16S rRNAor blaOXA-51 genes as the internal control. The ISAba1of blaOXA-23 gene was detected using combination ofprimers ISAba1-F/ISAba1-R and ISAba1-F/blaOXA-23-R (Table 1) [22]. The ISAba125 of blaNDM-1 gene weredetermined in all blaNDM-1 positive strains using com-bination of primers ISA125-F/ISA125-R and ISA125-F/blaNDM-R (Table 1). PCR products of the blaNDM-1
genes were purified and sequenced. BLAST was usedto compare the sequences of blaNDM-1 genes againstthe GenBank Database. PCR products were analyzedby electrophoresis in 1% agarose gel containing0.5 μg/ml ethidium bromides.
IPM-EDTA combined disk testAll blaNDM-1 positive strains were tested for MBLproduction by IPM-EDTA combined disk test. Thetest was performed as previously described [23]. After24 h incubation, the difference of inhibition zonediameter between IPM-EDTA disk and IPM diskalone (≥7 mm) was considered the positive criteriafor the presence of MBL.
Joshi et al. Antimicrobial Resistance and Infection Control (2017) 6:21 Page 2 of 7

Repetitive element PCR-mediated DNA fingerprinting(rep-PCR)Genomic DNA of each isolates was extracted from theovernight cultures using GF-1 bacterial DNA extractionkit (Vivantis, Malaysia). Rep-PCR was performed byusing genomic DNA as a template for PCR amplificationwith the ERIC-2 primer (Table 1) using condition as de-scribe previously [24, 25]. PCR-banding patterns andrep-PCR types were analyzed and interpreted as previ-ously described [25].
ResultsDemographic characteristic of patientsDemographic characteristics of the inpatients with A.baumannii infection were analyzed; 24 (54.5%) weremale and 20 (45.5%) were female. Most of the specimenswere from ICU wards (n = 27, 61.4%) (Fig. 1). Isolateswere collected from sputum (n = 26, 59.1%), tracheal as-pirates (n = 9, 20.4%), catheter tip, (n = 4, 9.1%), pus (n =4, 9.1%) and urine (n = 1, 2.3%) (Fig. 1).
Antibiotic susceptibilityOf the 44 isolates, resistance was found against cipro-floxacin (n = 43, 97.7%), cefotaxime (n = 43, 97.7%), cef-tazidime (n = 42, 95.4%), ceftriaxone (n = 41, 93.2%),cefepime (n = 39, 88.6%), amikacin (n = 19, 43.2%), gen-tamicin (n = 23, 52.3%), trimethoprim/sulfamethoxazole(n = 41, 93.2%), tetracycline (n = 21, 47.7%) and piperacil-lin/tazobactam (n = 43, 97.7%). Only one isolate ofA.baumannii was susceptible to all tested antibiotics.Most isolates (97.7%, n = 43) were carbapenem resistantA. baumannii (CR-AB); all CR-AB were MDR-AB. Oneisolate was detected to be XDR-AB. All the isolates werefully susceptible to colistin (MICs < 2 μg/ml) and MIC oftigecycline was determined to be <2.5 μg/ml (Table 2).
Antibiotic resistance genes and IS element in A.baumanniiAminoglycoside resistance gene, aphA6 and cephalo-sporin resistance genes, blaADC were detected in 40.1%(18/44) and 90.1% (40/44), respectively. The blaOXA-23
was present in all isolates. Other class D β-lactamase
Table 1 List of primer for detection of genes used in this study
Target genes Primer name Sequence 5’-3’ Size/ Annealing temp. References
16S rRNA 16S rRNA-F AGAGTTTGATCCTGGCTCAG 1500/58 [15]
16S rRNA-R ACGGCTACCTTGTTACGACTT
blaOXA-23 blaOXA-23-F GATCGGATTGGAGAACCAGA 501/52 [16]
blaOXA-23-R ATTTCTGACCGCATTTCCAT
blaOXA-51 blaOXA-51-F TAATGCTTTGATCGGCCTTG 353/52 [16]
blaOXA-51-R TGGATTGCACTTCATCTTGG
blaOXA-24 blaOXA-24-F GGTTAGTTGGCCCCCTTAAA 246/52 [16]
blaOXA-24-R AGTTGAGCGAAAAGGGGATT
blaOXA-58 blaOXA-58-F AAGTATTGGGGCTTGTGCTG 599/52 [16]
blaOXA-58-R CCCCTCTGCGCTCTACATAC
blaIMP blaIMP –F GGAATAGAGTGGCTTAAYTCTC 232/52 [20]
blaIMP –R GGTTTAAYAAAACAACCACC
blaVIM blaVIM –F GATGGTGTTTGGTCGCATA 390/52 [20]
blaVIM –R CGAATGCGCAGCACCAG
blaNDM blaNDM–F GGTTTGGCGATCTGGTTTTC 621/52 [21]
blaNDM–R CGGAATGGCTCATCACGATC
blaADC blaADC-F TAAACACCACATATGTTCCG 663/56 [19]
blaADC-F ACTTACTTCAACTCGCGACG
aphA6 aphA6-F ATGGAATTGCCCAATATTATTC 736/55 [14]
aphA6-R TCAATTCAATTCATCAAGTTTTA
ISAba1 ISAba1-F CATTGGCATTAAACTGAGGAGAAA 451/52 [22]
ISAba1-R TTGGAAATGGGGAAAACGAA
ISAba125 ISA125-F TGTTGAAGCGATCCGTTGTT 755/57 This study
ISA125-R GTGCGACAGTTTCAAAAGCCA
Rep-PCR ERIC-2 AAGTAAGTGACTGGGGTGAGCG variable length/45 [24]
Joshi et al. Antimicrobial Resistance and Infection Control (2017) 6:21 Page 3 of 7

genes, including blaOXA-24 and blaOXA-58, markers ofcarbapenem resistance in A. baumannii, were not de-tected in analyzed isolates. ISAba1 was detected in all ofblaOXA-23 positive isolates (100%). Of total analyzed iso-lates, 6 (13.6%) also harbored blaNDM-1 gene in additionto blaOXA-23 and blaOXA-51. All NDM-1 positive strainsexhibited insertion sequence ISAba125 detecting withprimers ISA125-F/ISA125-R. All isolates also detected aband of 1.6 kb in a PCR using ISA125-F/blaNDM-Rprimers. Metallo-β-lactamase (MBL) genes, includingblaVIM and blaIMP, were not detected in all isolates. Thesequences of the blaNDM-1 gene yielded 99-100% se-quence identity to the blaNDM-1 gene from Acinetobacterlwoffii strain WJ10621 plasmid pNDM-BJ01 (Accession:JQ001791) obtained from the GenBank Database.
MBL productionSix A. baumannii isolates harbored blaNDM-1 gene weredetected for MBL production. All of blaNDM-1 positivestrains were positive for MBL production. MBL positivestrains showed resistance to fluoroquinolones and β-lactam.
Epidemiological typingClonal relationship among isolates were studied usingrep-PCR typing. The fingerprinting represented 7 differ-ent DNA patterns consisting of 2 to 5 DNA fragment
sizes. The amplicons size for ERIC-2 PCR was 500–4000 bp. The genotype was named A-G as shown inFig. 2. The high prevalence genotype was type C (n = 14;31.8%) and D (n = 12; 27.3%). Genotype A, B, C and Dwere disseminated in all isolated ward (ICU, generalward and post-operative ward). Among 44 isolates, oneisolate of type F (2.3%) and G (2.3%) was found. Type Fwas obtained from a catheter tip specimen from the ICUward. Type G was obtained from sputum of a patientfrom a general ward. All NDM-1 positive strains exhib-ited genotype A (n = 1), B (n = 1), C (n = 3) and D(n = 1).
DiscussionsA. baumannii harboring blaOXA-51-like gene has beenidentified as a marker for species identification. An in-trinsic blaOXA-51-like gene detected in all isolates in thisstudy supports the use of this gene as a surrogatemarker of A. baumannii identification [8–10]. Highprevalence of cephalosporin resistance genes, blaADC
(90.1%) was found in this study. In addition, we found ahigh rate of cepharosporin resistant antibiotics (cefotax-ime, ceftazidime, ceftriaxone) using the disk diffusionmethod. These data indicated that cephalosporins nolonger work to treat A. baumannii isolated from Nepal.Carbapenem resistance in A. baumannii is a majorconcern and is most often associated with class D
Fig. 1 Distribution of A. baumannii carrying carbapenemase genes in different specimen types a and wards b
Table 2 The carbapenemases gene patterns, rep-PCR types and MIC determination of A. baumannii isolated from difference wards
Sites β-lactamase genepatterns
No. ofisolates
Rep-PCRTypes (n)
MIC (μg/ml) range
CAZ IMP TG CL
Intensive care unit blaOXA-51/ OXA-23 25 A (2), B (6), C (8), D (6), E (2), F (1) 4– > 256 1– > 32 1.6–3.9 0.13–2
blaOXA-51/ OXA-23/ NDM-1 2 B (1), C (1) >256 >32 1.7–3.2 0.61–0.79
General ward blaOXA-51/ OXA-23 9 A (1), B (1), C (1), D (5), G (1) >256 >24–>32 2–3.4 0.32–0.88
Post-operative ward blaOXA-51/ OXA-23 4 A (1), B (1), C (2) >256 >32 2.3–3.3 0.54–0.78
blaOXA-51/ OXA-23/ NDM-1 4 A (1), C (2) D (1) >256 >32 2.1–3.2 0.23–0.51
Abbreviations: CAZ ceftazidime, IPM imipenem, TG tigecycline, CL colistin
Joshi et al. Antimicrobial Resistance and Infection Control (2017) 6:21 Page 4 of 7

β-lactamases and MBLs. The full susceptibility of all CR-AB to colistin in this study indicates that colistin is stillan option of drug for the treatment of infections causedby A. baumannii in Nepal hospital.OXA-type carbapenemases are predominant in A.
baumannii [6, 7]. In agreement with this finding, highprevalence of blaOXA-23 carrying A. baumannii strainshas been reported in Nepalese patients [26]. The ac-quired blaOXA-23 is the dominant genetic determinant inAsia. The blaOXA-23 gene located on plasmid can betransferred between A. baumannii through conjugation.Thus, antibiotic resistant bacteria have been rapidly in-creasing worldwide [27]. The blaOXA-24 and blaOXA-58
were not detected in any isolates from this study. TheblaOXA-24/40 and blaOXA-58 genes were common in A.baumannii isolated from Europe [2, 28]. Recently,blaOXA-143 and blaOXA-235, which are novel class D β-lactamase genes in A. baumannii have been identified.To date, these determinants were detected only in Brazil,Mexico and the USA [29, 30]. ISAba1 was detected inwidespread clones of A. baumannii worldwide. Ourstudy found ISAba1 upstream of blaOXA-23 in all A. bau-mannii isolates. A correlation between A. baumanniiclusters carrying the ISAba1/blaOXA-23 gene and in-creased minimal inhibitory concentrations for carbapen-ems was reported [31]. One isolate (AB-13) that wasrecovered from catheter tips of long-stay hospital pa-tients showed an extreme drug resistance pattern (Add-itional file 1: Table S1). This isolate represented blaOXA-
23, blaADC and aphA6 genes. Further molecular study todetect other antibiotic resistance genes is needed to ex-plain what factors correlated with extreme drug resist-ance. We also found one isolate (AB-25) harboring
blaOXA-23, blaADC and aphA6 genes was sensitive to alltested drugs (Additional file 1: Table S1). This may be dueto the lack of promoter or mutation of ISAba1 or blaOXA-23
gene. Further study is needed to warrant the conclusion.The blaNDM-1 carrying A. baumannii has recently been
emerged in many countries, including Germany, Spain,Israel, Egypt, Switzerland, Libya, India, Pakistan andNepal [11, 26, 32, 33]. The blaNDM-1 gene has been iden-tified as a chimeric gene constructed by the fusion of theaminoglycoside-resistance gene aphA6 with a mannose-binding lectin gene. This event most likely occurs in Aci-netobacter spp., indicating that these bacteria are likelythe origin of this gene [34]. In this study, we identified13.6% of A. baumannii carrying blaNDM-1 gene. Previousstudy has identified high prevalence (24.6%) of the A. bau-mannii harbored the blaNDM-1 gene in Nepal in 2013–2014 [26]. Taking into consideration the relationship be-tween India, China and Nepal, the spread of blaNDM-1 islikely to occur rapidly, mostly through A. baumannii ra-ther than Enterobacteriaceae. A. baumannii able to trans-fer the blaNDM-1 gene via conjugation to the recipients andTn125 appears to be the main vehicle for dissemination ofthe blaNDM-1 genes in A. baumannii [35]. Poirel et al.reported that the blaNDM-1 gene was located within thecomposite transposon Tn125 bracketed by two copies of astrong promoter of blaNDM-1 gene called ISAba125 [11].This report was correlated with our finding that foundISAba125 in 100% of NDM-1 producing A. baumannii.The previous study reported that the most of A. bau-
mannii isolates harboring blaNDM-1 belonged to ST85and ST25 [35–37]. In Libyan hospital, Libya, the mainclone of imipenem-resistant NDM-1-producing A. bau-mannii belonged to ST2 [33]. We used rep-PCR typing
Fig. 2 Rep-PCR-based DNA fingerprint patterns of A. baumannii isolates. The lanes marked M contain molecular markers. Each lane representsgenotype patterns of A–G
Joshi et al. Antimicrobial Resistance and Infection Control (2017) 6:21 Page 5 of 7

to determine the clonal relationship in NDM-1 produ-cing A. baumannii. Our study highlighted that most ofNDM-1-producing A. baumannii isolates belonged to 4genotypes using rep-PCR. Rep-PCR is a method thatgenerates DNA fingerprints to discriminate betweenbacterial strains, and has been used to characterize A.baumannii isolates from hospitalized patients [38]. Ourrep-PCR typing represented a high genetic diversity(A-G) among A. baumannii isolates from Nepal. Someclonally related groups (A, B, C and D) were observed inthe all wards represented the disseminated of theseclones in the hospital. Four genotypes (A, B, C, and D)of co-existence of blaOXA-23 and blaNDM-1 A. baumanniiisolates were found. In addition, dissemination of thesefour genotypes into different wards also confirms as amajor epidemic. Since rep-PCR is less discriminatory formolecular typing of bacterial strains, further study usingmulti-locus sequence typing could be useful for epi-demiological investigations.
ConclusionAntibiotic resistance in A. baumannii is considered tobe a major future challenge in Nepal. Beyond OXA-typecarbapenemase, there is no doubt the emergence andspreads of NDM-1 encoding A. baumannii–a superbug–will further limit chemotherapeutic options and threatenthe public health of Nepal. The mechanism of hospitaladaptiveness beyond antibiotic resistance will be moredemanded in order to fully understand and combatMDR and XDR A. baumannii.
Additional file
Additional file 1: Table S1. Type of clinical specimen, ward, antibioticsusceptibility patterns, rep-PCR types, resistance genes and MIC of 44 A.baumannii isolates. (DOCX 23 kb)
AbbreviationsCR-AB: Carbapenem-resistant Acinetobacter baumannii; ICU: Intensive careunit; IPM-EDTA: Imipenem-ethylenediaminetetraacetic acid; MBL: Metallo-beta-lactamase; MDR-AB: Multidrug-resistant Acinetobacter baumannii;MIC: Minimum inhibitory concentration; NDM: New Delhi metallo-beta-lactamase; PCR: Polymerase chain reaction; XDR-AB: Extremely drug resistantAcinetobacter baumannii
AcknowledgementsWe thank Dr. Basant Pant, Neurosurgeon, Annapurna Neurological Instituteand Allied Sciences for providing laboratory facility and moral support. Wealso acknowledge Nepal Health Research Council for approving this study.
FundingThis study was partial funding by Thailand Research Fund (RSA5780015).
Availability of data and materialsPlease contact author for data requests.
Authors’ contributionsPRJ and MA designed the study, collected data, analyzed the data andprepared the manuscript, TK supervised the study, UL and RT collected data,
SS, analyzed the data, supervised the study and prepared the manuscript. Allauthors read and approved the manuscript.
Competing interestsThe authors declare that they have no competing interests.
Consent for publicationNot applicable.
Ethics approval and consent to participateEthical approval was obtained from the Ethical Review Board of Nepal HealthResearch Council (NHRC) (Reg. 27/2015). Informed consent was taken fromall the patients or patients’ guardians. The research was in compliance withthe Helsinki Declaration.
Author details1Central Department of Microbiology, Tribhuvan University, Kirtipur,Kathmandu, Nepal. 2Annapurna Neurological Institute and Allied Sciences,Maitighar, Kathmandu, Nepal. 3Department of Microbiology and Parasitology,Faculty of Medical Science, Naresuan University, Phitsanulok, Thailand.4Centre of Excellence in Medical Biotechnology, Faculty of Medical Science,Naresuan University, Phitsanulok, Thailand.
Received: 11 October 2016 Accepted: 3 February 2017
References1. Yamamoto M, Nagao M, Matsumura Y, Matsushima A, Ito Y, Takakura S, et al.
Interspecies dissemination of a novel class 1 integron carrying blaIMP-19 amongAcinetobacter species in Japan. J Antimicrob Chemother. 2011;66:2480–3.
2. Villalon P, Valdezate S, Medina-Pascual MJ, Carrasco G, Vindel A, Saez-NietoJA. Epidemiology of the Acinetobacter-derived cephalosporinase,carbapenem-hydrolysingoxacillinase and metallo-beta-lactamase genes, andof common insertion sequences, in epidemic clones of Acinetobacterbaumannii from Spain. J Antimicrob Chemother. 2013;68:550–3.
3. Agodi A, Voulgari E, Barchitta M, Quattrocchi A, Bellocchi P, Poulou A,et al. Spread of a carbapenem- and colistin-resistant Acinetobacterbaumannii ST2 clonal strain causing outbreaks in two Sicilian hospitals. JHosp Infect. 2014;86:260–6.
4. Heritier C, Poirel L, Lambert T, Nordmann P. Contribution of acquiredcarbapenem-hydrolyzing oxacillinases to carbapenem resistance inAcinetobacter baumannii. Antimicrob Agents Chemother. 2005;49:3198–202.
5. Quale J, Bratu S, Landman D, Heddurshetti R. Molecular epidemiology andmechanisms of carbapenem resistance in Acinetobacter baumannii endemicin New York City. Clin Infect Dis Off Publ Infect Dis Soc Am. 2003;37:214–20.
6. Amudhan MS, Sekar U, Kamalanathan A, Balaraman S. blaIMP and blaVIMmediated carbapenem resistance in Pseudomonas and Acinetobacter speciesin India. J Infect Develop Ctries. 2012;6:757–62.
7. Thomson JM, Bonomo RA. The threat of antibiotic resistance in Gramnegative pathogenic bacteria: beta-lactams in peril! Curr Opin Microbiol.2005;8:518–24.
8. Cicek AC, Saral A, Iraz M, Ceylan A, Duzgun AO, Peleg AY, et al. OXA- andGES-type beta-lactamases predominate in extensively drug-resistantAcinetobacter baumannii isolates from a Turkish University Hospital. ClinMicrobiol Infect. 2014;20:410–5.
9. Tsakris A, Ikonomidis A, Pournaras S, Tzouvelekis LS, Sofianou D, Legakis NJ,et al. VIM-1 metallo-beta-lactamase in Acinetobacter baumannii. Emerg InfectDis. 2006;12:981–3.
10. Kusradze I, Diene SM, Goderdzishvili M, Rolain JM. Molecular detection ofOXA carbapenemase genes in multidrug-resistant Acinetobacter baumanniiisolates from Iraq and Georgia. Int J Antimicrob Agents. 2011;38:164–8.
11. Poirel L, Bonnin RA, Boulanger A, Schrenzel J, Kaase M, Nordmann P. Tn125-related acquisition of blaNDM-like genes in Acinetobacter baumannii.Antimicrob Agents Chemother. 2012;56:1087–9.
12. Abbas M, Cherkaoui A, Fankhauser C, Schrenzel J, Harbarth S. Epidemiologyand clinical implications of carbapenemase-producing bacteria inSwitzerland. Rev Med Suisse. 2012;8:882–4.
13. Sarhaddi N, Soleimanpour S, Farsiani H, Mosavat A, Dolatabadi S, SalimizandH, et al. Elevated prevalence of multidrug-resistant Acinetobacter baumanniiwith extensive genetic diversity in the largest burn centre of northeast Iran.J Glob Antimicrob Resist. 2016;8:60–6.
Joshi et al. Antimicrobial Resistance and Infection Control (2017) 6:21 Page 6 of 7

14. Hujer KM, Hujer AM, Hulten EA, Bajaksouzian S, Adams JM, Donskey CJ, et al.Analysis of antibiotic resistance genes in multidrug-resistant Acinetobacter sp.isolates from military and civilian patients treated at the Walter Reed ArmyMedical Center. Antimicrob Agents Chemother. 2006;50:4114–23.
15. Misbah S, Hassan H, Yusof MY, Hanifah YS, Abu-Bakar S. Genomic speciesidentification of Acinetobacter of clinical isolates by 16S rDNA sequencing.Singapore. Med J. 2005;46:461–4.
16. Woodford N, Ellington MJ, Coelho JM, Turton JF, Ward ME, Brown S, et al.Multiplex PCR for genes encoding prevalent OXA carbapenemases inAcinetobacter spp. Int J Antimicrob Agents. 2006;27:351–3.
17. Clinical and Laboratory Standards Institute. Performance standards forantimicrobial susceptibility testing, document M100-24. Wayne: Clinical andLaboratory Standards Institute; 2014.
18. Magiorakos AP, Srinivasan A, Carey RB, Carmeli Y, Falagas ME, Giske CG,et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistantbacteria: an international expert proposal for interim standard definitions foracquired resistance. Clin Microbiol Infect. 2012;18:268–81.
19. Liu Y, Liu X. Detection of AmpC β-lactamases in Acinetobacter baumannii in theXuzhou region and analysis of drug resistance. Exp Ther Med. 2015;10:933–6.
20. Ellington MJ, Kistler J, Livermore DM, Woodford N. Multiplex PCR for rapiddetection of genes encoding acquired metallo-beta-lactamases. JAntimicrob Chemother. 2007;59:321–2.
21. Poirel L, Walsh TR, Cuvillier V, Nordmann P. Multiplex PCR for detection ofacquired carbapenemase genes. Diagn Microbiol Infect Dis. 2011;70:119–23.
22. Ruiz M, Marti S, Fernandez-Cuenca F, Pascual A, Vila J. Prevalence of IS(Aba1) in epidemiologically unrelated Acinetobacter baumannii clinicalisolates. FEMS Microbiol Lett. 2007;274:63–6.
23. Yong D, Lee K, Yum JH, Shin HB, Rossolini GM, Chong Y. Imipenem-EDTAdisk method for differentiation of metallo-beta-lactamase-producing clinicalisolates of Pseudomonas spp. and Acinetobacter spp. J Clin Microbiol. 2002;40:3798–801.
24. Vila J, Marcos MA, Jiminez de Anta MT. A comparative study of differentPCR-based DNA fingerprinting techniques for typing of the Acinetobactercalcoaceticus-A. baumannii complex. J Med Microbiol. 1996;44:482–9.
25. Grundmann HJ, Towner KJ, Dijkshoorn L, Gerner-Smidt P, Maher M, SeifertH, et al. Multicenter study using standardized protocols and reagents forevaluation of reproducibility of PCR-based fingerprinting of Acinetobacterspp. J Clin Microbiol. 1997;35:3071–7.
26. Shrestha S, Tadab T, Miyoshi-Akiyamac T, Oharad H, Shimadab K, Satoue K,et al. Molecular epidemiology of multidrug-resistant Acinetobacterbaumannii isolates in a university hospital in Nepal reveals the emergenceof a novel epidemic clonal lineage. Int J Antimicrob Agents. 2015;46:526–31.
27. Bertini A, Poirel L, Mugnier PD, Villa L, Nordmann P, Carattoli A.Characterization and PCR-based replicon typing of resistance plasmids inAcinetobacter baumannii. Antimicrob Agents Chemother. 2010;54:4168–77.
28. Cherkaoui A, Emonet S, Renzi G, Schrenzel J. Characteristics of multidrug-resistant Acinetobacter baumannii strains isolated in Geneva duringcolonization or infection. Ann Clin Microbiol Antimicrob. 2015;11:14–42.
29. Higgins PG, Pérez-Llarena FJ, Zander E, Fernández A, Bou G, Seifert H. OXA-235, a novel class D β-lactamase involved in resistance to carbapenems inAcinetobacter baumannii. Antimicrob Agents Chemother. 2013;57:2121–6.
30. Zander E, Bonnin RA, Seifert H, Higgins PG. Characterization of blaOXA-143variants in Acinetobacter baumannii and Acinetobacter pittii. AntimicrobAgents Chemother. 2014;58:2704–8.
31. Viana GF, Zago MC, Moreira RR, Zarpellon MN, Menegucci TC, Cardoso CL, et al.ISAba1/blaOXA-23: A serious obstacle to controlling the spread and treatment ofAcinetobacter baumannii strains. Am J Infect Control. 2016;44:593–5.
32. Decousser JW, Jansen C, Nordmann P, Emirian A, Bonnin RA, Anais L, et al.Outbreak of NDM-1-producing Acinetobacter baumannii in France, January toMay 2013. Eurosurveillance. 2013;18. doi:10.2807/1560-7917.ES2013.18.31.20547
33. Mathlouthi N, El Salabi AA, Ben Jomàa-Jemili M, Bakour S, Al-Bayssari C,Zorgani AA, et al. Early detection of metallo-β-lactamase NDM-1- and OXA-23 carbapenemase-producing Acinetobacter baumannii in Libyan hospitals.Int J Antimicrob Agents. 2016;48:46–50.
34. Toleman MA, Spencer J, Jones L, Walsh TR. blaNDM-1 is a chimera likelyconstructed in Acinetobacter baumannii. Antimicrob Agents Chemother.2012;56:2773–76.
35. Ramoul A, Loucif L, Bakour S, Amiri S, Dekhil M, Rolain JM. Co-occurrence ofblaNDM-1 with blaOXA-23 or blaOXA-58 in clinical multidrug-resistantAcinetobacter baumannii isolates in Algeria. J Glob Antimicrob Resist.2016;6:136–41.
36. Rafei R, Pailhoriès H, Hamze M, Eveillard M, Mallat H, Dabboussi F, et al. Molecularepidemiology of Acinetobacter baumannii in different hospitals in Tripoli,Lebanon using blaOXA-51-like sequence based typing. BMC Microbiol. 2015;15:11–7.
37. Bakour S, Touati A, Bachiri T, Sahli F, Tiouit D, Naim M, et al. First report of16S rRNA methylase ArmA-producing Acinetobacter baumannii and rapidspread of metallo-β-lactamase NDM-1 in Algerian hospitals. 2014. J InfectChemother. 2014;20:696–701.
38. Pasanen T, Koskela S, Mero S, Tarkka E, Tissari P, Vaara M, et al. Rapidmolecular characterization of Acinetobacter baumannii clones with rep-PCRand evaluation of carbapenemase genes by new multiplex PCR in hospitaldistrict of Helsinki and Uusimaa. Plos One. 2014;9:e85854.
• We accept pre-submission inquiries
• Our selector tool helps you to find the most relevant journal
• We provide round the clock customer support
• Convenient online submission
• Thorough peer review
• Inclusion in PubMed and all major indexing services
• Maximum visibility for your research
Submit your manuscript atwww.biomedcentral.com/submit
Submit your next manuscript to BioMed Central and we will help you at every step:
Joshi et al. Antimicrobial Resistance and Infection Control (2017) 6:21 Page 7 of 7

EPIDEMIOLOGY
Dissemination of blaOXA-23, blaOXA-24, blaOXA-58,and blaNDM-1 Genes of Acinetobacter baumanniiIsolates from Four Tertiary Hospitals in Thailand
Udomluk Leungtongkam,1 Rapee Thummeepak,1 Suchada Wongprachan,2
Pollawat Thongsuk,3 Thawatchai Kitti,4 Kwanjai Ketwong,5 Chakkaphan Runcharoen,6
Narisara Chantratita,6 and Sutthirat Sitthisak1,7
Acinetobacter baumannii is a major threat to public health due to the emergence and dissemination ofantibiotic-resistant strains. The purpose of this study was to determine the molecular epidemiology of antibiotic-resistant A. baumannii isolates collected from four tertiary hospitals in Thailand during the period November2013–February 2015. We screened 339 A. baumannii, nonrepetitive clinical isolates to determine drug sus-ceptibility. Among all isolates, we found that 7.9% was nondrug-resistant A. baumannii (NR-AB). Carbapenem-resistant A. baumannii (CR-AB) strains accounted for 84.9% of the total isolates, with extensively drug-resistantA. baumannii (XDR-AB) accounting for 7.9% of the total isolates. We further investigated class D carbape-nemase genes using multiplex-PCR amplification and class B metallo-b-lactamase genes, including blaIMP,blaVIM, and blaNDM-1 genes, using PCR and sequencing methods. We found that 300 (88.5%) isolates carriedacquired class D carbapenemase genes, including blaOXA-23 (82.6%), blaOXA-24 (0.3%), and blaOXA-58 (6.5%).The genes blaIMP and blaVIM were not detected in any isolates. The blaNDM-1 was detected in 31 isolates fromtwo hospitals (9.1%). All of the blaNDM-1-positive A. baumannii (NDM-AB) had ISAba125 sequences upstreamof the blaNDM-1 gene. A coexistence of three resistance genes—blaOXA-23-blaOXA-58-blaNDM-1—was found inone isolate. A repetitive element palindromic-PCR (REP-PCR) revealed that all A. baumannii isolates weregenetically diverse and could be divided into 33 genotypes. Only three genotypes were found to be predominantin all hospitals. Data from our study indicate the widespread emergence of multiple resistance determinants inA. baumannii isolates in Thailand, suggesting the need for more stringent infection control measures.
Keywords: carbapenemase, metallo-b-lactamase, Acinetobacter baumannii, blaOXA-23 gene, blaNDM-1 gene
Introduction
Acinetobacter baumannii is a Gram-negative bacte-rium usually associated with hospital-acquired infection.
In the past decades, the prevalence of carbapenem-resistantA. baumannii (CR-AB) and extensively drug-resistant A. bau-mannii (XDR-AB) has been increasing in hospitals worldwide.The major resistance mechanisms to carbapenems inA. baumannii are the production of carbapenem-hydrolyzing-oxacillinases belonging to molecular class D (OXA-typecarbapenemases) and also by other metallo-b-lactamases(MBLs) belonging to class B enzymes. Although in some
parts of the world, class B MBLs are an increasing prob-lem, their prevalence is still far from OXA b-lactamases.1
MBLs have potent carbapenemase activity which resistsb-lactamase inhibitors but is susceptible to inhibition bymetal ion chelators.2 Since the 1990s, several MBLs en-coded by mobile DNA elements have emerged in A. bau-mannii in many countries, of which blaNDM-1, a novel typeof MBL, is a major concern. New Delhi Metallo-beta-lactamase-1 (NDM-1) producing bacteria were first reportedin 2009.3 Since then, there have been many reports ofblaNDM-1-positive bacteria disseminated worldwide.4 Thefirst identification of NDM-1-producing bacteria in Thailand
1Department of Microbiology and Parasitology, Faculty of Medical Science, Naresuan University, Phitsanulok, Thailand.2Phichit Hospital, Amphoe Meuang, Phichit, Thailand.3Chiangrai Prachanukroh Hospital, Amphoe Meuang, Chiangrai, Thailand.4Department of Oriental Medicine, ChiangRai College, Chiangrai, Thailand.5Sawan Pracharak Hospital, Amphoe Meuang, Nakorn Sawan, Thailand.6Department of Microbiology and Immunology, Faculty of Tropical Medicine, Mahidol University, Bangkok, Thailand.7Centre of Excellence in Medical Biotechnology, Naresuan University, Phitsanulok, Thailand.
MICROBIAL DRUG RESISTANCEVolume 00, Number 00, 2017ª Mary Ann Liebert, Inc.DOI: 10.1089/mdr.2016.0248
1

have been detected in Escherichia coli, Klebsiella pneu-moniae, and Citrobacter freundii but not found in anyAcinetobacter spp.5 Class D carbapenemases are encodedby acquired antibiotic resistance genes blaOXA-23, blaOXA-24,and blaOXA-58, as well as intrinsic blaOXA-51.6,7 These genesare chromosomally encoded or carried on plasmids.6 Themost commonly acquired class D carbapenemase genes inAsia is blaOXA-23, which is highly disseminated among A.baumannii in Thailand.8,9
A. baumannii is the leading cause of nosocomial pathogenin Thailand, but the genes essential for antibiotic resistanceremain largely unknown. The need to conduct an epidemi-ology survey to identify the genetic mechanisms of antibi-otic resistance remains. Thus, in this study, our objectivewas to determine the prevalence of class B MBLs and classD carbapenemase genes in A. baumannii isolates, from fourtertiary hospitals in Thailand.
Materials and Methods
Bacterial isolates and geographiccharacteristics of hospitals
A total of 339 samples were collected from the inpatientunits of four tertiary hospitals in Thailand during the periodNovember 2013–February 2015. The specimens were iso-lated from the outpatient department (OPD) and varioushospital wards, including Medicine, Intensive Care Units(ICUs), Surgery, Monk, Very Important Person, Pediatrics,and Trauma. For the purpose of our analysis and discussion,the four hospitals are designated as HA, HB, HC, and HD(Table 1). HC is located in a tourism region and border areaof Thailand, and HD is surrounded by a cluster of indus-trial factories with a substantial population of transientforeign workers. Standard biochemical tests were carriedout on the samples collected from the hospitals to identifyA. baumannii, and this was confirmed by the PCR method todetect the 16S rRNA gene and the intrinsic blaOXA-51
gene.7,10,11 A. baumannii ATCC 19606 was used as control.
Antimicrobial susceptibility testing
The antibiotic susceptibility of all the isolates was deter-mined using the disc diffusion method. The results were in-terpreted according to Clinical and Laboratory StandardsInstitute guidelines.12,13 Disc diffusion tests were performedwith the following 15 antibiotics (Oxoid, Basingstoke, Eng-land): amikacin (30mg), cefepime (30mg), cefotaxime (30mg),cefoperazone/sulbactam (105mg), ceftazidime (30mg), cef-triaxone (30mg), ciprofloxacin (5mg), colistin (10mg), genta-micin (10mg), imipenem (10mg), meropenem (10mg),piperacillin/tazobactam (100/10mg), tetracycline (30mg), ti-gecycline (15mg), and trimethoprim/sulfamethoxazole (1.25/23.75mg). Multidrug-resistant A. baumannii (MDR-AB), CR-AB, and XDR-AB were classified as previously described byMagiorakos et al.14
Detection of antibiotic resistance genes
DNA was extracted from all isolates and subjected tomultiplex PCR to detect blaOXA-23, blaOXA-24, blaOXA-58,and intrinsic blaOXA-51 genes.7 PCR was used to screen forMBL genes: blaIMP, blaVIM, and blaNDM-1.15 The ISAba125of blaNDM-1 gene was determined in all blaNDM-1 positive
strains using PCR with a primer pair (SupplementaryTable S1; Supplementary Data are available online at www.liebertpub.com/mdr) designed by the Primer-BLAST program(www.ncbi.nlm.nih.gov/tools/primer-blast). The blaNDM-1 genefrom Acinetobacter lwoffii strain WJ10621 plasmid pNDM-BJ01 (GenBank Accession No. JQ001791) was used as acontrol. An upstream of the ISAba125 was determined usingprimers ISA125-F/blaNDM-R (Supplementary Table S1).PCR products of the blaNDM-1 genes were purified and se-quenced. BLAST was used to compare the sequences ofblaNDM-1 genes against the GenBank database (https://blast.ncbi.nlm.nih.gov BLAST). All PCR products were analyzedby electrophoresis in 1% agarose gel containing 0.5 mg/mlethidium bromide.
MBL production
All metallo-b-lactamase positives found by PCR werefurther analyzed for MBL production by the combined disktest. An increased inhibition zone with a diameter of ‡7 mmwith imipenem-EDTA disk compared to imipenem diskalone was considered as MBL positive.16
Repetitive element palindromic-PCR
The genetic diversity among A. baumannii isolates wasstudied using repetitive element palindromic-PCR (REP-PCR) typing. The REP-PCR analysis was performed inall isolates using the ERIC-2 primers (SupplementaryTable S1) according to the minor modification of previousstudy.17 The condition of ERIC-2 PCR was one cycle at95�C for 5 minutes, followed by 45 cycles at 94�C for 1minute, 45�C for 1 minute, and 72�C for 2 minutes. TheREP-PCR was analyzed by the banding patterns. The iso-lates were assigned to the same REP-PCR type if theirpatterns differed by fewer than two bands, according topreviously defined criteria.17,18 Using REP-PCR, a strainwas considered to be ‘‘epidemic’’ when it had one or twopredominant genotypes that isolated from various patients ineach hospital. In contrast, a strain was considered as ‘‘spo-radic’’ when it had many genotypes, and each genotype hada few isolates in each hospital.
Results
Isolation and identification of antibiotic-resistantA. baumannii
A. baumannii isolates were collected from sputum (82.0%),urine (4.1%), pus (9.1%), blood (1.8%), body fluid (pleuralfluid, ascitic fluid, and drain) (1.2%), wound (1.2%), andmedical devices (0.6%) (Table 1). Most of the A. baumanniiisolates in our study were isolated from the medical ward(46.3%) (Table 1). All isolates were positive by PCR for de-tecting the 16S rRNA gene and the intrinsic blaOXA-51 gene.The antimicrobial susceptibility of all A. baumannii isolates to15 antibiotics is shown in Fig. 1. Among the 339 isolates, 27isolates (7.9%) were susceptible to all the antibiotics tested.All isolates were sensitive to colistin and tigecycline. Morethan 70% of the A. baumannii isolates were resistant to cefo-taxime (CTX), ceftazidime (CAZ), ceftriaxone (CRO), andciprofloxacin (CIP). More than 65% of the A. baumannii iso-lates were resistant to imipenem and meropenem. The preva-lence of MDR-AB from the four hospitals was 90.3%. CR-AB
2 LEUNGTONGKAM ET AL.

Ta
ble
1.
Geo
gra
ph
ic
Ch
ara
cteristics
of
Fo
ur
Ho
spita
ls,
Specim
en
Ty
pes,
Iso
la
ted
Wa
rd
s,
an
dR
EP
-ty
pes
of
Clin
ica
lS
pecim
en
sU
sed
in
Th
is
Stu
dy
Hosp
ital
code
Loca
tion
(No.
of
bed
s)C
oll
ecti
on
per
iod
No.
of
isola
tes
Spec
imen
types
(No.
and
per
centa
ge
of
each
spec
imen
type)
Ward
s(N
o.
and
per
centa
ge
of
each
ward
)R
EP
-typ
es(N
o.
of
isola
tes)
HA
Cen
tral
(653)
Novem
ber
–D
ecem
ber
2013
62
Sputu
m(4
8,
77.4
)M
edic
ine
(30,
48.4
){R
1(5
),R
2(1
),R
4(6
),R
6(4
),R
7(1
),R
8(1
),R
9(1
),R
10(1
),R
11(2
),R
12(5
),R
15(2
),R
16(3
1),
R26(1
),R
27(1
)}U
rine
(6,
9.7
)IC
U(1
5,
24.2
)P
us
(1,
1.6
)S
urg
ery
(10,
16.1
)B
ody
fluid
(2,
3.2
)M
onk
(1,
1.6
)W
ound
(4,
6.5
)V
IP(2
,3.2
)M
edic
aldev
ices
(1,
1.6
)P
edia
tric
s(4
,6.5
)
HB
Low
erN
ort
her
n(4
05)
August
–S
epte
mber
2014
54
Sputu
m(2
9,
53.7
)M
edic
ine
(30,
55.6
){R
4(4
),R
6(1
),R
11(2
),R
12(1
5),
R14(1
),R
16(3
1)}
Uri
ne
(5,
9.2
)IC
U(4
,7.4
)P
us
(16,
29.6
)S
urg
ery
(17,
31.5
)B
ody
fluid
(1,
1.9
)V
IP(2
,3.7
)B
lood
(2,
3.7
)O
PD
(1,
1.8
)M
edic
aldev
ices
(1,
1.9
)
HC
Nort
her
n(7
56)
July
–O
ctober
2014
106
Sputu
m(1
06,
100)
Med
icin
e(4
4,
41.5
){R
1(9
),R
2(2
),R
4(5
),R
6(4
),R
8(1
),R
12(9
),R
13(1
),R
14(1
),R
16(3
0),
R18(1
),R
20(1
),R
21(5
),R
22(1
),R
24(2
9),
R25(1
),R
28(3
),R
30(1
),R
31(1
),R
33(1
)}
ICU
(31,
29.3
)S
urg
ery
(17,
16.0
)M
onk
(8,
7.6
)O
PD
(1,
0.9
)P
edia
tric
s(1
,0.9
)T
raum
a(4
,3.8
)
HD
Eas
t(5
61)
Dec
ember
–F
ebru
ary
2015
117
Sputu
m(9
5,
81.2
)M
edic
ine
(53,
45.3
){R
1(9
),R
3(4
),R
4(1
2),
R5(5
),R
8(3
),R
12(9
),R
16(6
5),
R17(1
),R
18(2
),R
19(4
),R
23(1
),R
29(1
),R
32(1
)}U
rine
(3,
2.6
)IC
U(2
7,
23.1
)P
us
(14,
12.0
)S
urg
ery
(20,
17.1
)B
lood
(4,
3.4
)M
onk
(3,
2.5
)B
ody
fluid
(1,
0.8
)V
IP(7
,6.0
)O
PD
(5,
4.3
)P
edia
tric
s(2
,1.7
)
All
hosp
ital
s(2
,375)
Novem
ber
2013–
Feb
ruar
y2015
339
Sputu
m(2
78,
82.0
)M
edic
ine
(157,
46.3
){R
1(2
3),
R2(3
),R
3(4
),R
4(2
7),
R5(5
),R
6(9
),R
7(1
),R
8(5
),R
9(1
),R
10(1
),R
11(4
),R
12(3
8),
R13(1
),R
14(2
),R
15(2
),R
16(1
57),
R17(1
),R
18(3
),R
19(4
),R
20(1
),R
21(5
),R
22(1
),R
23(1
),R
24(2
9),
R25(1
),R
26(1
),R
27(1
),R
28(3
),R
29(1
),R
30(1
),R
31(1
),R
32(1
),R
33(1
)}
Uri
ne
(14,
4.1
)IC
U(7
7,
22.7
)P
us
(31,
9.1
)S
urg
ery
(64,
18.9
)B
lood
(6,
1.8
)M
onk
(12,
3.5
)B
ody
fluid
(4,
1.2
)V
IP(1
1,
3.2
)W
ound
(4,
1.2
)O
PD
(7,
2.1
)M
edic
aldev
ices
(2,
0.6
)P
edia
tric
s(7
,2.1
)T
raum
a(4
,1.2
)
ICU
,in
tensi
ve
care
unit
;O
PD
,outp
atie
nt
dep
artm
ent;
VIP
,ver
yim
port
ant
per
son;
RE
P,
repet
itiv
eel
emen
tpal
indro
mic
.
3

FIG. 1. Antibiotic resistance patterns of 339 Acinetobacter baumannii isolates from patients of four hospitals in Thailand.AK, amikacin; FEP, cefepime; SCF, cefoperazone/sulbactam; CTX, cefotaxime; CAZ, ceftazidime; CRO, ceftriaxone, CIP,ciprofloxacin; CT, colistin; CN, gentamicin; IPM, imipenem; MEM, meropenem; PIP, piperacillin/tazobactam; TE, tetra-cycline; TGC, tigecycline; SXT, trimethoprim/sulfamethoxazole.
FIG. 2. Prevalence (%) of MDR-AB, CR-AB, and XDR-AB (A) and dissemination of blaOXA-23, blaOXA-24, blaOXA-58, andblaNDM-1 genes of Acinetobacter baumannii among four hospitals in Thailand (B). Other *(A) represented the percentage ofA. baumannii isolates, which are non-MDR (non-MDR classified as nonresistant or less resistant than the three antibiotic groups).Each bar (B) represented the percent of antibiotic resistance genes of A. baumannii isolates. No ABGs (B) represented percent ofA. baumannii isolates which did not harbor the blaOXA-23, blaOXA-24, blaOXA-58, and blaNDM-1 genes. CR-AB, carbapenem-resistant A. baumannii; MDR-AB, multidrug-resistant A. baumannii; XDR-AB, extensively drug-resistant A. baumannii.
4

was found in 84.9%, and XDR-AB was found in 7.9%. XDR-AB strains were found in each hospital with the prevalencerange of 5.7–12.9%. All XDR-AB were CR-AB (Fig. 2A).
Detection of class D carbapenemase and MBL genes
We found high prevalence of blaOXA-23 in all isolateswhich was 82.6% (280/339). The prevalence of blaOXA-58
and blaOXA-24 was 6.5% (22/339) and 0.3% (1/339), re-spectively (Supplementary Fig. S1A). The blaOXA-23 genewas detected in 79.1% of the CR-AB strains from the fourhospitals, while the blaOXA-24 gene was detected only in HAwith a prevalence of 1.6% of all isolates from HA (Fig. 2B).The genes blaIMP and blaVIM were not detected in any iso-lates. Among the 339 isolates, blaNDM-1 was detected in 31(9.1%) isolates from two hospitals [2.6% (9/339) HC and6.5% (22/339) HD]. In all the blaNDM-1 positive strains,ISAba125 was detected upstream of the blaNDM-1 gene(Supplementary Fig. S1B). A coexistence of two resistancegenes—blaOXA-23-blaNDM-1—was detected in HC (0.9%)and HD (18.8%) (Fig. 2B). A coexistence between the twopairs of resistance genes blaOXA-23-blaOXA-58 (1.9%) andblaOXA-58-blaNDM-1 (3.8%), and also among the three re-sistance genes, blaOXA-23-blaOXA-58-blaNDM-1 (0.9%), wasfound only in HC (Fig. 2B). The MBL production by thecombined disk test demonstrated that 100% of 31 isolates ofthe blaNDM-1 positive strain were MBL-producing isolates.
Epidemiological typing
The fingerprinting represented 33 different DNA pat-terns consisting of amplicon sizes ranging from 400 to5,000 bp. The genotype was named R1 to R33 (Table 1). As
shown in Fig. 3A, A. baumannii isolates from HA, HC, andHD represented 14, 19, and 13 genotypes, respectively,while isolates from HB could be typed only into six geno-types, which were R4, R6, R11, R12, R14, and R16. Wefound only three genotypes (R4, R12, R16) distributed inall the four hospitals (Fig. 3A). In this study, we dividedA. baumannii isolates into three different groups, whichwere nondrug-resistant A. baumannii (NR-AB) 27 isolates,NDM-AB 31 isolates, and XDR-AB 27 isolates. One isolatewith a phenotype of both NDM-AB and XDR-AB representedgenotype R1. REP-typing analysis of three different groups isshown in Fig. 3B. NR-AB isolates belonged to 12 genotypes,while XDR-AB was characterized by only four genotypes.The NDM-AB belonged to five genotypes, which were R1 toR5. REP-PCR genotypes R1 and R4 were presented in allthree groups. In addition, A. baumannii blaOXA-23 positiveisolates were divided into 26 genotypes. Only one isolatewhich harbored blaOXA-24 gene was genotype R16. Twenty-two A. baumannii isolates carried the blaOXA-58 gene andhad three genotypes, which were R1, R12, and R16.
Sequencing analysis
The sequences of the blaNDM-1 gene from hospitals HCand HD yielded 100% sequence identity to the blaNDM-1
gene from A. lwoffii strain WJ10621 plasmid pNDM-BJ01(Accession: JQ001791) obtained from the GenBank data-base. The nucleotide sequences of the blaNDM-1 genes of theisolates from these two hospitals were similar, and thesenucleotide sequences from each hospital were deposited inthe GenBank database under the following accession num-bers KX447524 and KX495193.
FIG. 3. Genetic diversity using REP-PCR of all Acinetobacter baumannii from Thailand. (A) Genetic diversity of isolates fromeach hospital. (B) REP-types of NR-AB, NDM-AB, and XDR-AB. Color in B denotes the three groups of A. baumannii used forREP-PCR. The number of isolates per set is indicated in bars. NDM-AB, blaNDM-1-positive A. baumannii; NR-AB, nondrug-resistant A. baumannii; XDR-AB, extensively drug-resistant A. baumannii; REP-PCR, repetitive element palindromic-PCR.
ANTIBIOTIC RESISTANCE OF ACINETOBACTER BAUMANNII 5

Discussion
A. baumannii infection is a major global concern ofnosocomial origin worldwide. In the present study, weconfirmed the potential impact of A. baumannii on respira-tory infections in Thailand. Eighty-two percent of thespecimens were isolated from sputum. This result is con-sistent with the data of A. baumannii isolates obtained fromHua Hin hospital, Thailand in 2014.19 The prevalence ofCR-AB has been increasing in Thailand and worldwide.Unfortunately, carbapenems, the first antibiotic used to treatthe A. baumannii infection, can no longer be used to treatCR-AB, due to its resistance.20 In our study, we observedthat most of the A. baumannii isolates (84.9%) are resistantto the carbapenem antibiotic group, including imipenem andmeropenem. High rates of MDR-AB (90.3%) and CR-AB(84.9%) were found in all A. baumannii isolates from all thefour hospitals. However, we found lower rates of XDR-AB(5.7–12.9%) than were previously reported in Thailand.21
Our study also showed that tigecycline and colistin are stilleffective drugs for the treatment of MDR-AB, since wefound no resistance to these antibiotics.
The mechanisms of carbapenem resistance in A. bau-mannii are mostly associated with class D b-lactamase,22
but the prevalence of class B MBL in A. baumannii has beenincreasingly reported in some parts of the world.1 TheblaOXA-58 gene is highly prevalent among A. baumanniiisolates in Mexico and Spain.23,24 The first reported pres-ence of this gene in Thailand was in 2013.25 Subsequentreports from Thailand in 2013 and 2015 indicated thepresence of CR-AB, which carries blaOXA-23, blaOXA-24, andblaOXA-58; and further the blaOXA-23 gene was found in 85–100% of A. baumannii isolates.8,9 In our study, we showedthat most of MDR-AB and CR-AB harbored the blaOXA-23
gene. The lowest occurrence of blaOXA-24 (1.6%) was foundin samples from HA, located in the central part of Thailand.A previous study which analyzed A. baumannii isolatesfrom hospitals in the central region of Thailand found highprevalence (50%) of blaOXA-24 among A. baumannii strainsproducing MBL.25
The first identification of New Delhi MBL-1 (blaNDM-1)in Thailand, in 2012, was in Enterobacteriaceae—two iso-lates of E. coli, two isolates of K. pneumoniae, and twoisolates of C. freundii.5 We identified the blaNDM-1 gene inA. baumannii in two hospitals in Thailand, one located inthe northern region and the other in the eastern region. Thefirst hospital HC is located in the northern tourism regionand border area adjoining Myanmar and the second hospitalHD in eastern Thailand, surrounded by a cluster of industrialfactories with many foreign workers. A horizontal genetransfer of antibiotic-resistant genes might occur due tothe movement of tourists and foreign workers.26 The pres-ence of the blaNDM-1 gene was found in 8.5% (9/106) ofA. baumannii isolates taken from HC and 18.8% (22/117) ofA. baumannii isolates from HD. These results are similar tothose from several previous studies, which variously re-ported that the blaNDM-1 gene was found in 1.7–11.4% ofA. baumannii isolates tested during 2013–2014 in Bangla-desh, Pakistan, and India.27–29 The blaNDM-1 gene is knownto be carried by chromosome and plasmids, which are themajor causes of the spread of antibiotic-resistant genesamong many bacteria, through horizontal gene transfer.30,31
ISAba125 (IS30 family) has been reported to relate to theacquisition of blaNDM-like genes in A. baumannii.32 In ourstudy, all of the blaNDM-1 positive isolates were positive forISAba125. The ISAba125 element, which has been locatedupstream of the blaNDM-1 gene, is a strong promoter ofthe expression of this gene.33 It is possible that ISAba125may be a vehicle for the dissemination of blaNDM-1 gene inA. baumannii.29
We found class D b-lactamase genes and blaNDM-1 co-existing in two hospitals in Thailand. This result was similarto previous studies in Japan and Switzerland, which foundthe coexistence of blaOXA-23 and blaNDM-1 genes.34,35 Inaddition, the coexistence of blaOXA-58 and blaNDM-1 geneswas found in Acinetobacter junii and Acinetobacter pit-tii.36,37 The coexistence of three resistant genes—blaOXA-23,blaOXA-69, and blaNDM-1—in A. baumannii isolated fromrectal swabs was also reported in Switzerland.38 Further-more, in the Czech Republic, Krizova et al. found thatA. baumannii ANC4097 harbored five genes encodingb-lactamase, including blaNDM-1, blaOXA-23, blaOXA-69,blaTEM-1, and blaAmp-C.39 In our study, we found only oneisolate (0.3%) from HC hospital which was positive forblaOXA-23, blaOXA-58, and blaNDM-1, from a total of 339A. baumannii isolates taken from four hospitals in Thailand.This study is the first report of blaNDM-1, blaOXA-23, andblaOXA-58 genes in A. baumannii in the same isolates.
The REP-PCR method has previously been used to dis-criminate A. baumannii isolates worldwide.17,18,40 Our REP-PCR typing revealed a high genetic diversity (R1–R33)among A. baumannii isolates. Our epidemiological studyhighlighted the fact that most of the isolates belonged togenotype R16 (157/339). This type was the major epidemicstrains in HA, HB, and HD. In contrast, we found genotypeR16 and genotype R24 as two major epidemic strains in HC.A previous study showed the presence of blaOXA-23 andblaOXA-58 genes in the same major REP-PCR clone ofA. baumannii.40 Our study displayed the blaOXA-23 positiveisolates (151/280) in all the four hospitals representing themajor genotype, R16. The major REP-PCR typing presentedin XDR-AB was genotype R4 (22/27). This strain was ep-idemic in all XDR-AB from four hospitals. Furthermore, wefound that all XDR-AB genotype R4 isolates were positivefor the blaOXA-23 gene. Most of A. baumannii harboredblaNDM-1 gene isolates (NDM-AB) collected from HC andHD hospitals were genotype R1 (17/31), and all of NDM-AB genotype R1 were classified as the epidemic strain. Incontrast, NR-AB strains had 12 genotypes and each of thegenotype had a few isolates. All these strains were consid-ered as sporadic strains.
In conclusion, our study indicated an increase in the preva-lence of antibiotic-resistant A. baumannii bacteria. This studyindicated the dissemination of class D b-lactamase genes andblaNDM-1 in many regions of Thailand. The blaOXA-23 harboredA. baumannii isolates are found most abundantly. A greatereffort for infection control is essential to prevent the dissemi-nation of the A. baumannii harboring class D b-lactamase andblaNDM-1 determinants in hospitals in Thailand.
Acknowledgments
The authors thank staff of four hospitals for providing theA. baumanni isolates. Many thanks to Mr. Roy Morien of
6 LEUNGTONGKAM ET AL.

the Naresuan University Language Centre for his Englishediting in this document. This study was funded by ThailandResearch Fund (RSA5780015). U.L. was supported by aGraduate student scholarship from Naresuan University.R.T. was supported by The Royal Golden Jubilee PhDProgram (PHD/0031/2558).
Disclosure Statement
No competing financial interests exist.
References
1. Mathlouthi, N., A.A. El Salabi, M. Ben Jomaa-Jemili, S.Bakour, C. Al-Bayssari, A.A. Zorgani, A. Kraiema, O.Elahmer, L. Okdah, J.M. Rolain, and C. Chouchani. 2016.Early detection of metallo-b-lactamase NDM-1- and OXA-23 carbapenemase-producing Acinetobacter baumannii inLibyan hospitals. Int. J. Antimicrob. Agents 48:46–50.
2. Queenan, A.M., and K. Bush. 2007. Carbapenemases: theversatile b-lactamases. Clin. Microbiol. Rev. 20:440–458.
3. Yong, D., M.A. Toleman, C.G. Giske, H.S. Cho, K.Sundman, K. Lee, and T.R. Walsh. 2009. Characterizationof a new metallo-beta-lactamase gene, bla(NDM-1), and anovel erythromycin esterase gene carried on a unique ge-netic structure in Klebsiella pneumoniae sequence type 14from India. Antimicrob. Agents. Chemother. 53:5046–5054.
4. Berrazeg, M., S. Diene, L. Medjahed, P. Parola, M. Drissi,D. Raoult, and J. Rolain. 2014. New Delhi Metallo-beta-lactamase around the world: an eReview using GoogleMaps. Euro. Surveill. 19: pii: 20809.
5. Rimrang, B., A. Chanawong, A. Lulitanond, C. Wilai-luckana, N. Charoensri, P. Sribenjalux, W. Phumsrikaew,L. Wonglakorn, A. Kerdsin, and P. Chetchotisakd. 2012.Emergence of NDM-1- and IMP-14a-producing En-terobacteriaceae in Thailand. J. Antimicrob. Chemother.67:2626–2630.
6. Peleg, A.Y., H. Seifert, and D.L. Paterson. 2008. Acineto-bacter baumannii: emergence of a successful pathogen.Clin. Microbiol. Rev. 21:538–582.
7. Woodford, N., M.J. Ellington, J.M. Coelho, J.F. Turton,M.E. Ward, S. Brown, S.G. Amyes, and D.M. Livermore.2006. Multiplex PCR for genes encoding prevalent OXAcarbapenemases in Acinetobacter spp. Int. J. Antimicrob.Agents 27:351–353.
8. Jantama, S.S., T. Kunawatanakul, S. Semaram, S. Sangsa-non, N. Teerawattanasuk, and K. Jantama. 2013. Detectionof carbapenemase and blaOXA-23-like gene in carbapenem-resistant Acinetobacter baumannii at SunpasitthiprasongHospital. I. J. P. S. 9:99–109.
9. Teo, J., T.P. Lim, L.Y. Hsu, T.Y. Tan, S. Sasikala, P.Y.Hon, A.L. Kwa, and A. Apisarnthanarak. 2015. Extensivelydrug-resistant Acinetobacter baumannii in a Thai hospital:a molecular epidemiologic analysis and identification ofbactericidal Polymyxin B-based combinations. Antimicrob.Resist. Infect. Control. 4:2.
10. Misbah, S., H. Hassan, M.Y. Yusof, Y.A. Hanifah, and S.AbuBakar. 2005. Genomic species identification of Acine-tobacter of clinical isolates by 16S rDNA sequencing.Singapore. Med. J. 46:461–464.
11. Turton, J.F., N. Woodford, J. Glover, S. Yarde, M.E.Kaufmann, and T.L. Pitt. 2006. Identification of Acineto-bacter baumannii by detection of the blaOXA-51-like carba-
penemase gene intrinsic to this species. J. Clin. Microbiol.44:2974–2976.
12. Clinical and Laboratory Standards Institute. 2014. Perfor-mance Standards for Antimicrobial Susceptibility Testing;Twenty-Fourth Informational Supplement. M100-S24.CLSI, Wayne, PA.
13. Piewngam, P., and P. Kiratisin. 2014. Comparative as-sessment of antimicrobial susceptibility testing for tigecy-cline and colistin against Acinetobacter baumannii clinicalisolates, including multidrug-resistant isolates. Int. J. An-timicrob. Agents 44:396–401.
14. Magiorakos, A.P., A. Srinivasan, R.B. Carey, Y. Carmeli,M.E. Falagas, C.G. Giske, S. Harbarth, J.F. Hindler, G.Kahlmeter, B. Olsson-Liljequist, D.L. Paterson, L.B. Rice,J. Stelling, M.J. Struelens, A. Vatopoulos, J.T. Weber, andD.L. Monnet. 2012. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an internationalexpert proposal for interim standard definitions for acquiredresistance. Clin. Microbiol. Infect. 18:268–281.
15. Poirel, L., T.R. Walsh, V. Cuvillier, and P. Nordmann.2011. Multiplex PCR for detection of acquired carbape-nemase genes. Diagn. Microbiol. Infect. Dis. 70:119–123.
16. Yong, D., K. Lee, J.H. Yum, H.B. Shin, G.M. Rossolini,and Y. Chong. 2002. Imipenem-EDTA disk method fordifferentiation of metallo-beta-lactamase-producing clini-cal isolates of Pseudomonas spp. and Acinetobacter spp. J.Clin. Microbiol. 40:3798–3801.
17. Reboli, A.C., E.D. Houston, J.S. Monteforte, C.A. Wood,and R.J. Hamill. 1994. Discrimination of epidemic andsporadic isolates of Acinetobacter baumannii. J. Clin. Mi-crobiol. 32:2635–2640.
18. D’Arezzo, S., A. Capone, N. Petrosillo, P. Visca; GRAB,M. Ballardini, S. Bartolini, E. Bordi, A. Di Stefano, M.Galie, R. Minniti, M. Meledandri, L. Pacciani, G. Parisi, G.Prignano, C. Santini, M. Valmarin, M. Venditti, and S.Ziantoni. 2009. Epidemic multidrug-resistant Acinetobacterbaumannii related to European clonal types I and II inRome (Italy). Clin. Microbiol. Infect. 15:347–357.
19. Santimaleeworagun, W., A. Thathong, W. Samret, P. Pre-echachuawong, W. Sae-lim, and T. Jitwasinkul. 2014.Identification and characterization of carbapenemase genesin clinical isolates of carbapenem-resistant Acinetobacterbaumannii from general hospital in Thailand. SoutheastAsian J. Trop. Med. Public. Health 45:874–880.
20. Viehman, J.A., M.H. Nguyen, and Y. Doi. 2014. Treatmentoptions for carbapenem-resistant and extensively drug-resistantAcinetobacter baumannii infections. Drugs. 74:1315–1355.
21. Lawan, A., D. Pornphan, U. Fuangfa, T. Sudaluck, S.Rudiwilai, and S. Kanokrat. 2009. Epidemiological char-acteristics of Acinetobacter baumannii infections at Phra-mongkutklao Hospital. J. Med. Assoc. Thai. 92:164–172.
22. Al Atrouni, A., M. Hamze, T. Jisr, C. Lemarie, M. Eveil-lard, M.L. Joly-Guillou, and M. Kempf. 2016. Wide spreadof OXA-23-producing carbapenem-resistant Acinetobacterbaumannii belonging to clonal complex II in differenthospitals in Lebanon. Int. J. Infect. Dis. 52:29–36.
23. Bocanegra-Ibarias, P., C. Pena-Lopez, A. Camacho-Ortiz,J. Llaca-Dıaz, J. Silva-Sanchez, H. Barrios, U. Garza-Ramos, A.M. Rodrıguez-Flores, and E. Garza-Gonzalez.2015. Genetic characterization of drug resistance and clonaldynamics of Acinetobacter baumannii in a hospital settingin Mexico. Int. J. Antimicrob. Agents. 45:309–313.
24. Villalon, P., S. Valdezate, M.J. Medina-Pascual, G. Carras-co, A. Vindel, and J.A. Saez-Nieto. 2013. Epidemiology of
ANTIBIOTIC RESISTANCE OF ACINETOBACTER BAUMANNII 7

the Acinetobacter-derived cephalosporinase, carbapenem-hydrolysing oxacillinase and metallo-b-lactamase genes,and of common insertion sequences, in epidemic clones ofAcinetobacter baumannii from Spain. J. Antimicrob. Che-mother. 68:550–553.
25. Jumroon, N., and P. Santanirand. 2013. Multiple combi-nation patterns of oxa-type carbapenem hydrolyzing andmetallo-b-lactamases encoding genes among clinicallyisolated Acinetobacter baumannii. Int. J. Pharm. Bio. Sci.4:908–917.
26. Bogaerts, P., R. Rezende de Castro, S. Roisin, A. Deplano,T.D. Huang, M. Hallin, O. Denis, and Y. Glupczynski.2012. Emergence of NDM-1-producing Acinetobacterbaumannii in Belgium. J. Antimicrob. Chemother. 67:1552–1553.
27. Farzana, R., S. Shamsuzzaman, and K.Z. Mamun. 2013.Isolation and molecular characterization of New Delhimetallo-beta-lactamase-1 producing superbug in Bangla-desh. J. Infect. Dev. Ctries. 7:161–168.
28. Hasan, B., K. Perveen, B. Olsen, and R. Zahra. 2014.Emergence of carbapenem-resistant Acinetobacter bau-mannii in hospitals in Pakistan. J. Med. Microbiol. 63:50–55.
29. Mishra, S., M.R. Sen, S. Upadhyay, and A. Bhattacharjee.2013. Genetic linkage of blaNDM among nosocomial iso-lates of Acinetobacter baumannii from a tertiary referralhospital in northern India. Int. J. Antimicrob. Agents 41:452–456.
30. Krahn, T., D. Wibberg, I. Maus, A. Winkler, S. Bontron, A.Sczyrba, P. Nordmann, A. Puhler, L. Poirel, and A. Schluter.2016. Intraspecies transfer of the chromosomal Acineto-bacter baumannii blaNDM-1 carbapenemase gene. Anti-microb. Agents. Chemother. 60:3032–3040.
31. Zhang, C., S. Qiu, Y. Wang, L. Qi, R. Hao, X. Liu, Y. Shi,X. Hu, D. An, Z. Li, P. Li, L. Wang, J. Cui, P. Wang, L.Huang, J.D. Klena, and H. Song. 2013. Higher isolation ofNDM-1 producing Acinetobacter baumannii from thesewage of the hospitals in Beijing. PLoS One 8:6.
32. Pfeifer, Y., G. Wilharm, E. Zander, T.A. Wichelhaus, S.Gottig, K.P. Hunfeld, H. Seifert, W. Witte, and P.G. Hig-gins. 2011. Molecular characterization of blaNDM-1 in anAcinetobacter baumannii strain isolated in Germany in2007. J. Antimicrob. Chemother. 66:1998–2001.
33. Wang, Y., C. Wu, Q. Zhang, J. Qi, H. Liu, Y. Wang, T. He,L. Ma, J. Lai, Z. Shen, Y. Liu, and J. Shen. 2012. Identi-
fication of New Delhi metallo-b-lactamase 1 in Acineto-bacter lwoffii of food animal origin. PLoS One 7:5.
34. Cherkaoui, A., S. Emonet, G. Renzi, and J. Schrenzel.2015. Characteristics of multidrug-resistant Acinetobacterbaumannii strains isolated in Geneva during colonization orinfection. Ann. Clin. Microbiol. Antimicrob. 14:42.
35. Nakazawa, Y., R. Ii, T. Tamura, T. Hoshina, K. Tamura, S.Kawano, T. Kato, F. Sato, T. Horino, M. Yoshida, S. Hori,M. Sanui, Y. Ishii, and K. Tateda. 2013. A case of NDM-1-producing Acinetobacter baumannii transferred from Indiato Japan. J. Infect. Chemother. 19:330–332.
36. Ang, G.Y., C.Y. Yu, Y.M. Cheong, W.F. Yin, and K.G.Chan. 2016. Emergence of ST119 Acinetobacter pittii co-harbouring NDM-1 and OXA-58 in Malaysia. Int. J. An-timicrob. Agents 47:168–169.
37. Regeen, H., D. Al-Sharafa-Kittaneh, R. Kattan, R. Al-Dawodi, H. Marzouqa, and M.Y. Hindiyeh. 2014. Firstreport of blaNDM and blaOXA-58 coexistence in Acineto-bacter junii. J. Clin. Microbiol. 52:3492–3493.
38. Bonnin, R.A., L. Poirel, T. Naas, M. Pirs, K. Seme, J.Schrenzel, and P. Nordmann. 2012. Dissemination ofNew Delhi metallo-b-lactamase-1-producing Acinetobacterbaumannii in Europe. Clin. Microbiol. Infect. 18:E362–E365.
39. Krizova, L., R.A. Bonnin, P. Nordmann, A. Nemec, and L.Poirel. 2012. Characterization of a multidrug-resistantAcinetobacter baumannii strain carrying the blaNDM-1 andblaOXA-23 carbapenemase genes from the Czech Republic.J. Antimicrob. Chemother. 67:1550–1552.
40. Pasanen, T., S. Koskela, S. Mero, E. Tarkka, P. Tissari, M.Vaara, and J. Kirveskari. 2014. Rapid molecular charac-terization of Acinetobacter baumannii clones with rep-PCRand evaluation of carbapenemase genes by new multiplexPCR in hospital district of Helsinki and Uusimaa. PLoSOne 9:1.
Address correspondence to:Sutthirat Sitthisak, PhD
Department of Microbiology and ParasitologyFaculty of Medical Science
Naresuan UniversityPhitsanulok 65000
Thailand
E-mail: [email protected]
8 LEUNGTONGKAM ET AL.




















