
UJI VIABILITAS BIJI POHON KAYU PUTIH SUCI (Vitex ...perpustakaan.fmipa.unpak.ac.id/file/UJI...
20
UJI VIABILITAS BIJI POHON KAYU PUTIH SUCI (Vitex leucoxylon L.F. forma leucoxylon) PADA BERBAGAI PERLAKUAN CAHAYA DAN KARAKTER SIMPAN Brihaspati garba S.Si 1 , Dr. Prasetyorini, MS 2 , Dr. Dian Latifah, S.P.,M.App.Sc 3 Program Studi Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Pakuan Abstrak Penelitian tanaman yang berpotensi sebagai tanaman obat meningkat di seluruh dunia karena potensi penggunaannya dalam sistem pengobatan tradisional untuk mengobati berbagai penyakit. Genus Vitex termasuk sekitar 270 spesies yang diketahui dari pohon dan semak- semak di daerah tropis dan sub-tropis. Tanaman Vitex leucoxylon merupakan tanaman yang sangat berpotensi sebagai tanaman obat, tetapi belum banyak penelititan dan pembudidayaan yang dilakukan mengenai viabilitas dan karakter simpan dari tanaman tersebut. Tujuan penelitian ini untuk mengetahui pengaruh cahaya terhadap viabilitas biji V. leucoxylon dan karakter simpan biji V. leucoxylon. Penelitian dilakukan di Rumah Kaca dan Bank Biji Subbidang Registrasi dan Pembibitan, Bidang Konservasi Ex-situ Pusat Konservasi Tumbuhan Kebun Raya Bogor pada bulan April sampai dengan Juli 2016. Penelitian dilakukan dengan 2 tahap, tahap pertama adalah penyimpanan dan penyemaian biji V. Leucoxylon dan tahap kedua adalah perkecambahan biji dengan perlakuan cahaya. Kata kunci : Viabilitas, Vitex leucoxylon, penyimpanan dan penyemaian biji, perlakuan cahaya PENDAHULUAN Penelitian tanaman yang berpotensi sebagai tanaman obat meningkat di seluruh dunia karena potensi penggunaannya dalam sistem pengobatan tradisional untuk mengobati berbagai penyakit. Berbagai tanaman yang berpotensi sebagai tanaman obat telah diidentifikasi dan digunakan untuk mempelajari khasiat, keselamatan dan kemanjuran penggunaannya. Hasil menyoroti potensi besar tanaman obat di bidang farmakologi. Genus Vitex termasuk sekitar 270 spesies yang diketahui dari pohon dan semak-semak di daerah tropis dan sub-tropis, meskipun beberapa spesies juga ditemukan di daerah beriklim. Tanaman Vitex leucoxylon merupakan tanaman yang sangat berpotensi sebagai tanaman obat. Tanaman ini berasal dari India dan merupakan tanaman yang sering dimanfaatkan sebagai tanaman obat di India. Tanaman dari famili Lamiaceae ini memiliki potensi untuk menyembuhkan berbagai macam penyakit seperti diabetes, radang penyakit, gangguan hati dan penyakit yang dimediasi oleh radikal bebas (Meena et al, 2011).Di Indonesia sendiri belum banyak pembudidayaan tanaman V. leucoxylon karena belum diketahui mengenai karakter biji serta belum banyak pembudidaya tanaman V. leucoxylon yang membudidayakannya dan memanfaatkannya sebagai obat tradisional (Sutjipto dan Widiyastuti, 2009).
Transcript of UJI VIABILITAS BIJI POHON KAYU PUTIH SUCI (Vitex ...perpustakaan.fmipa.unpak.ac.id/file/UJI...

UJI VIABILITAS BIJI POHON KAYU PUTIH SUCI (Vitex leucoxylon L.F. forma
leucoxylon) PADA BERBAGAI PERLAKUAN CAHAYA DAN KARAKTER SIMPAN
Brihaspati garba S.Si1, Dr. Prasetyorini, MS2, Dr. Dian Latifah, S.P.,M.App.Sc3
Program Studi Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam
Universitas Pakuan
Abstrak
Penelitian tanaman yang berpotensi sebagai tanaman obat meningkat di seluruh dunia
karena potensi penggunaannya dalam sistem pengobatan tradisional untuk mengobati berbagai
penyakit. Genus Vitex termasuk sekitar 270 spesies yang diketahui dari pohon dan semak-
semak di daerah tropis dan sub-tropis. Tanaman Vitex leucoxylon merupakan tanaman yang
sangat berpotensi sebagai tanaman obat, tetapi belum banyak penelititan dan pembudidayaan
yang dilakukan mengenai viabilitas dan karakter simpan dari tanaman tersebut. Tujuan
penelitian ini untuk mengetahui pengaruh cahaya terhadap viabilitas biji V. leucoxylon dan
karakter simpan biji V. leucoxylon. Penelitian dilakukan di Rumah Kaca dan Bank Biji
Subbidang Registrasi dan Pembibitan, Bidang Konservasi Ex-situ Pusat Konservasi Tumbuhan
Kebun Raya Bogor pada bulan April sampai dengan Juli 2016. Penelitian dilakukan dengan 2
tahap, tahap pertama adalah penyimpanan dan penyemaian biji V. Leucoxylon dan tahap kedua
adalah perkecambahan biji dengan perlakuan cahaya.
Kata kunci : Viabilitas, Vitex leucoxylon, penyimpanan dan penyemaian biji, perlakuan cahaya
PENDAHULUAN
Penelitian tanaman yang berpotensi
sebagai tanaman obat meningkat di seluruh
dunia karena potensi penggunaannya dalam
sistem pengobatan tradisional untuk
mengobati berbagai penyakit. Berbagai
tanaman yang berpotensi sebagai tanaman
obat telah diidentifikasi dan digunakan
untuk mempelajari khasiat, keselamatan
dan kemanjuran penggunaannya. Hasil
menyoroti potensi besar tanaman obat di
bidang farmakologi. Genus Vitex termasuk
sekitar 270 spesies yang diketahui dari
pohon dan semak-semak di daerah tropis
dan sub-tropis, meskipun beberapa spesies
juga ditemukan di daerah beriklim.
Tanaman Vitex leucoxylon merupakan
tanaman yang sangat berpotensi sebagai
tanaman obat. Tanaman ini berasal dari
India dan merupakan tanaman yang sering
dimanfaatkan sebagai tanaman obat di
India. Tanaman dari famili Lamiaceae ini
memiliki potensi untuk menyembuhkan
berbagai macam penyakit seperti diabetes,
radang penyakit, gangguan hati dan
penyakit yang dimediasi oleh radikal bebas
(Meena et al, 2011).Di Indonesia sendiri
belum banyak pembudidayaan tanaman V.
leucoxylon karena belum diketahui
mengenai karakter biji serta belum banyak
pembudidaya tanaman V. leucoxylon yang
membudidayakannya dan
memanfaatkannya sebagai obat tradisional
(Sutjipto dan Widiyastuti, 2009).

Biji merupakan sumber dari
kehidupan tumbuhan. Pohon yang telah
berusia ratusan tahun pun berawal dari
sebuah biji yang dikecambahkan. Begitu
pentingnya peranan sebuah biji dalam
mendukung keberlangsungan kehidupan
jenisnya. Kita mengenal ada dua macam
jenis biji, yaitu biji ortodok dan biji
rekalsitran (Nurwardani, 2008).
Pengaruh intensitas cahaya yang
rendah terhadap hasil berbagai komoditi
sudah banyak dilaporkan. Naungan 50%
pada genotipe padi yang sensitif
menyebabkan jumlah gabah/malai kecil
serta persentase gabah hampa yang tinggi,
sehingga produksi biji rendah (Sopandie et
al. 2003). Intensitas cahaya rendah pada
saat pembungaan padi dapat menurunkan
karbohidrat yang terbentuk, sehingga
menyebabkan meningkatnya gabah hampa
(Chaturvedi et al. 1994).
Sampai saat ini belum banyak
diketahui lama waktu yang diperlukan
untuk menyimpan biji di kebun raya. Selain
itu, lebih dari 50.000 jenis tumbuhan atau
20% dari total tumbuhan di dunia
menghasilkan biji yang tidak dapat
mempertahankan viabilitasnya selama
dalam penyimpanan (rekalsitran), dan
masih banyak ketidakpastian tentang jenis
tumbuhan mana yang rekalsitran dan mana
yang bukan (Irawanto, 2009).
Pengetahuan mengenai viabilitas biji
masih sangat terbatas dan penelitian yang
banyak dilakukan baru terbatas pada
tanaman budidaya. Penelitian tentang biji
tanaman liar/tumbuhan hutan sangat
diperlukan, terutama untuk menentukan
cara yang tepat dalam mempertahankan
viabilitas dan laju kerusakan biji selama
penyimpanan (Irawanto, 2009).
Berdasarkan hal tersebut penelitian ini
dilakukan untuk mengetahui viabilitas biji
tanaman hutan yang berpotensi sebagai
tanaman obat di Kebun Raya Bogor untuk
mempertahankan kelangsungan plasma
nutfah dalam konservasi ex-situ.
BAHAN DAN METODE PENELITIAN
Waktu dan Lokasi Penelitian
Penelitian dilaksanakan dari bulan
April sampai dengan bulan Juli 2016 di
Rumah Kaca dan Bank Biji Subbidang
Registrasi dan Pembibitan, Bidang
Konservasi Ex-situ Pusat Konservasi
Tumbuhan Kebun Raya Bogor.
Bahan
Bahan yang digunakan adalah biji
dari buah V. leucoxylon yang berasal dari
koleksi tumbuhan Kebun Raya Bogor
dengan No. Vax XI.I. 46, asal koleksi India
yang dipanen langsung di Kebun Raya
Bogor pada bulan April 2016. Bahan lain
yang digunakan air, dithane, dan moss.

Alat
Alat yang digunakan dalam
penelitian ini chest freezer -21 oC,
desikator, timbangan analitik digital, lux
meter, oven, kalkulator, timer, ayakan,
polybag ukuran 10cm x 15cm, sarung
tangan, plastik air sealed, bak plastik,
aluminium foil sealed, log book, label
mikolin, kamera, digital caliper, RHS
colour chart, box kayu dengan ukuran
40cm x 40cm, lampu neon (36 W/840 putih
dingin Philips Lifemax), lampu pijar (100
W), 3-mm filter plexiglas merah,3-mm
filter plexiglass merah jauh.
Persiapan Penelitian
Pemanenan Buah
Buah V. leucoxylon dipanen dengan
cara memungut buah yang telah masak
secara fisiologis dan terjatuh dari pohonnya
dapat dilihat pada Gambar 1. Teknik
pemanenan buah ini dikarenakan pohon
yang terlalu tinggi sehingga tidak dapat
memanen buah menggunakan galah untuk
memanennya. Pemanenan buah dengan
metode ini diharuskan dengan sangat teliti,
karena buah yang dipanen harus dalam
keadaan utuh dan tidak busuk untuk
menghasilkan kualitas biji yang bagus
untuk diproses ke tahap selanjutnya.
Pemanenan buah dilakukan selama 1
minggu.
Gambar 1. Pengumpulan Buah V.
leucoxylon
A. Proses Pemanenan Buah B. Hasil
Panen.
Karakterisasi Buah dan Biji
Setelah proses pemanenan buah, buah
V. leucoxylon langsung diproses untuk
karakterisasi buah dan biji. Buah yang
sudah terkumpul lalu dipilih 30 buah untuk
proses karakterisasi buah dan biji.
Karakterisasi buah dan biji Vitex leucoxylon
dimulai dari pengamatan warna buah,
pengukuran dimensi buah meliputi
diameter buah dan biji, dan panjang buah
dan biji. Pengamatan warna buah
menggunakan Royal Horticultural Society
(RHS) Colour Chart dapat dilihat pada
Gambar 2. Sedangkan pengukuran panjang
dan diameter buah dan biji menggunakan
Digital Calliper NANKAI 150mm dapat
dilihat pada Gambar 3.
Gambar 2. RHS Colour Chart
A B

Gambar 3. Pengukuran Panjang dan
Diameter Buah dan Biji
A. Digital Calliper NANKAI 150mm. B.
Pengukuran Diameter Buah. C.
Pengukuran Diameter Biji.
Ekstraksi Buah
Proses ekstraksi buah dilakukan
setelah proses karakterisasi buah dan biji.
Pengekstraksian buah menggunakan
ayakan bambu untuk memisahkan daging
buah dari biji dapat dilihat pada Gambar 4.
Biji Vitex leucoxylon yang sudah terpisah
dari kulit dan daging buahnya lalu dicuci
bersih sampai tidak ada daging buah yang
menempel pada biji. Biji harus bersih dari
daging buahnya karena biji yang masih
menempel dengan daging buahnya dapat
menenyebabkan tumbuhnya jamur pada
saat proses penyimpanan. Setelah biji
dicuci bersih lalu biji dikering anginkan,
setelah biji kering lalu biji dapat diproses
ketahap pengukuran kadar air.
Gambar 4. Proses Ekstraksi Buah V.
leucoxylon
A. Ekstraksi Buah. B. Pengeringan Buah
Dibawah Sinar Matahari
Pengukuran Kadar Air Awal
Menurut Internasional Seed Testing
Asosiation (ISTA)(2010), pengukuran
kadar air awal dengan metode
langsung/oven dilakukan tiga kali sebagai
ulangan. Cawan petri yang sudah
dikeringkan di oven pada suhu 105oC
selama 15-30 menit lalu dimasukan ke
dalam desikator selama 15-30 menit.
Setelah itu, cawan petri ditimbang. Cawan
kemudian diisi dengan biji yang sudah
dikeringkan kemudian ditimbang
menggunakan timbangan analitik digital.
Biji dan cawan petri lalu dikeringkan di
dalam oven pada suhu 105oC selama 18 jam
(ambil waktu pukul 13:00-08:00 WIB). Jika
suhu 300oC maka pengeringan hanya
dilakukan selama 2-4 jam. Setelah kering
biji dan cawan petri kembali ditimbang
untuk penentuan kadar air awal.
Cara penghitungan kadar air biji yaitu
dengan rumus (Bank Biji Kebun Raya):
A
C
B
A
B

Penentuan Bobot 1000 butir Biji
Menurut Direktorat Perbenihan
Tanaman Hutan (2002), pengukuran bobot
1000 butir benih dilakukan dengan cara
menimbang bobot dan contoh kerja masing-
masing 100 butir biji secara acak (dalam
gram). Dalam kegiatan penelitian ini
penentuan bobot 1000 butir dihitung
dengan cara menimbang 100 butir biji
kemudian hasilnya diekstrapolasi dan
dikalikan 10 sehingga akan diperoleh hasil
bobot 1000 butir benih.
Pengujian 100 Biji untuk Penentuan
Karakter Simpan Biji
Menentukan kadar air (ISTA, 2007)
dari 10 biji individual, melaksanakan tes
perkecambahan awal, pada suhu optimum
untuk spesies, pada dua sampel dari 13 biji.
32 biji kering dicampur dengan berat yang
sama dari silika gel dalam wadah tertutup
yang sesuai, disimpan dalam inkubator
pada 15oC (untuk spesies beriklim) atau
25oC (untuk spesies tropis). Silika gel
diganti dan menimbang berat biji setiap 1-
3 hari, tergantung pada ukuran benih.
Setiap berat biji menurun, periode ini dapat
diperpanjang. Ketika biji mencapai berat
konstan pada kesetimbangan, biasanya
setelah 2-3 minggu untuk menentukan
eRH. Sampel harus dikeringkan untuk
melengkapi 15% Erh. Setelah itu diambil 6
biji untuk menentukan kadar air setelah
desikasi. Pada saat yang sama, tempatkan
32 biji kontrol pada kelembaban yang
tinggi dalam wadah tertutup, menggunakan
vermiculite basah, untuk menjaga kondisi
lembab. Simpan di bawah suhu yang sama
sebagai sampel kering (15oC atau 25oC)
untuk jumlah waktu yang sama. Setelah
periode ini, pindahkan biji dari
penyimpanan lembab untuk menentukan
Erh, dan ambil 6 biji untuk menentukan
kadar air seperti di atas. Imbibisi sisa biji
kering dan melakukan tes perkecambahan
pada dua sampel dari 13 biji, melakukan tes
perkecambahan pada dua sampel dari 13
biji dari sampel kontrol. Plot kemajuan
perkecambahan kurva (% perkecambahan x
masa inkubasi) untuk perkecambahan awal
biji segar, perkecambahan setelah
pengeringan dan perkecambahan setelah
penyimpanan lembab.
Penentuan kadar air simpan pengujian
100 biji dihitung dengan rumus (Bank
Biji Kebun Raya):
Pengujian Daya Simpan Untuk
Penentuan Karakter Simpan Biji Vitex
leucoxylon
Uji perkecambahan secara berkala
untuk mengetahui karakter simpan dari biji
V. leucoxylon. Penyemaian biji dilakukan
dalam periode: minggu ke 0, minggu ke 2,
minggu ke 4, dan minggu ke 6. Biji
disimpan dalam kemasan aluminium foil
sealed dan disimpan pada 4 perlakuan
simpan yaitu penyimpanan dalam chest

freezer -21oC, penyimpanan pada ruang AC
dengan suhu 18oC dengan kelembaban 70%
tanpa media, penyimpanan pada ruang AC
dengan suhu pengujian 18oC dengan
kelembaban 70% menggunakan media
moss, dan penyimpanan pada suhu ruang
26-28oC.
Penyemaian Biji
Penyemaian biji V. leucoxylon
dengan kadar air 13,4 % untuk mengetahui
metode perkecambahan yang terbaik.
Sebelum penyemaian biji di imbibisi
selama 24 jam,. Biji yang telah diimbibisi
kemudian direndam Dithane selama 5
menit untuk mecegah pertumbuhan jamur
pada saat biji disemai. Media
perkecambahan yang digunakan adalah
moss. Media disterilisasi dengan cara
menyiraminya dengan air panas suhu 100o
C. Biji V. leucoxcylon ditanam setelah
media dingin sejumlah 10 biji/polybag.
Perlakuan Cahaya
Percobaan ini meneliti efek dari
perbedaan kualitas spektral cahaya pada
perkecambahan biji V. leucoxylon. Semua
biji sebelum penanaman sudah di imbibisi
terlebih dahulu. Sumber cahaya putih
adalah lampu neon (36 W / 840 putih dingin
Philips Lifemax). Cahaya spektrum merah
dipasok oleh satu lampu neon didasari
dengan 3-mm filter plexiglas merah tebal
yang berfungsi sebagai filter cahaya pada
jarak 1 m (Borthwick et al, 1954). Sumber
cahaya spektrum merah jauh terdiri dari
satu lampu pijar (100 W) dan lampu neon
disaring melalui 3-mm filter plexiglass
merah jauh. Jarak antara lampu pijar adalah
60cm (Borthwick et al, 1954). Spektrum
ditularkan melalui setiap filter lalu
diperiksa menggunakan spektrofotometer
untuk mengkonfirmasi transmisi cahaya
merah (M) pada 600 nm dan merah jauh
(MJ) pada 730 nm. Benih disinari dalam
kotak kayu kedap cahaya dengan ukuran 40
cm x 40 cm. Selain perlakuan cahaya pada
laboratorium, perlakuan cahaya juga di
lakukan pada rumah kaca dengan
perbandingan naungan. Perbandingan
naungan yang dilakukan pada perlakuan
cahaya di rumah kaca yaitu perlakuan
cahaya tanpa naungan paranet, perlakuan
cahaya dengan naungan paranet 55%, dan
perlakuan cahaya dengan naungan paranet
65%.
Tabel 1. Hasil Pengukuran Intensitas
Cahaya di Lingkungan Penelitian

Metode Penelitian
Penelitian dilakukan dengan 2 tahap,
tahap pertama adalah penyimpanan dan
penyemaian biji V. leucoxylon. Biji
disimpan menggunakan aluminium foil
sealed, biji disimpan dalam 4 periode
penyimpanan (0,2,4,6 minggu) dengan 4
perlakuan penyimpanan yaitu :
1. Penyimpanan dalam ruang simpan
dengan suhu 18oC menggunakan
media moss dengan kelembaban 70%.
2. Penyimpanan dalam ruang simpan
dengan suhu 18oC tanpa menggunakan
media dengan kelembaban 70%.
3. Penyimpanan dalam Chest freezer
dengan suhu -21o C.
4. Penyimpanan pada suhu ruangan 26-
28o C.
Penelitian tahap kedua adalah
perkecambahan biji dengan perlakuan
cahaya. Penelitian tahap kedua ini adalah
dengan 7 perlakuan cahaya yaitu :
Perlakuan perkecambahan rumah kaca:
1. Rumah kaca tanpa naungan.
2. Rumah kaca dengan naungan paranet
55%.
3. Rumah kaca dengan naungan paranet
65%.
Perlakuan perkecambahan spektrum
cahaya:
1. Laboratorium dengan cahaya spektrum
merah.
2. Laboratorium dengan cahaya spektrum
merah jauh.
3. Laboratorium gelap.
4. Laboratorium terbuka.
Pengamatan dilakukan setiap hari
dengan mengamati Daya Kecambah Total
(DKT), Koefisien Kecepatan
Perkecambahan (KCP), Koefisien
Keserempakan Perkecambahan (KSP),
Hari Pertama Berkecambah (HPB), Hari
Terakhir berkecambah (HTB), P50.
1. Daya Kecambah Total dihitung dengan
rumus (DKT) (Bank Biji Kebun Raya):
DK = 𝑛
𝑁 x 100
DK = Daya kecambah; n = benih yang
berkecambah; N = benih yang
dikecambahkan.
2. Koefisien Kecepatan Perkecambahan
dihitung dengan rumus (KCP) (Bank
Biji Kebun Raya):
X =∑𝑛
∑(𝑡x𝑛) x 100
X = koefisien kecepatan berkecambah, n =
benih yang dikecambahkan, t = hari saat
benih berkecambah.
3. Koefisien Keserempakan
Perkecambahan dihitung dengan
rumus (KSP) (Bank Biji Kebun Raya): ∑𝑛
∑{(𝑇−𝑡)2 x n )} x 100 dimana T =
∑(𝑡 x 𝑛)
∑𝑛
∑𝑛 = jumlah total biji berkecambah; t x n =
n benih yang berkecambah pada hari ke-t.
4. Perkecambahan mencapai 50% (P50)
adalah waktu yang diperlukan biji
untuk dapat berkecambah sebanyak
50% dari hari setelah tanam (HST).
5. Hari Pertama Berkecambah (HPB)
dihitung waktu awal biji berkecambah
dari hari setelah tanam (HST).
6. Hari Terakhir Berkecambah (HTB)
adalah waktu terakhir biji berkecambah
yang diperlukan biji berkecambah dari
hari setelah tanam (HST).

Analisis Data
Pengaruh perlakuan diuji dengan uji
Duncan pada taraf 95% analisis statistik
RAL untuk perlakuan cahaya dan analisis
statistik RAL Faktorial untuk perlakuan
periode simpan dan perlakuan
penyimpanan yang menghitung Daya
Kecambah Total (DKT), Koefisien
Kecepatan Perkecambahan (KCP), dan
Koefisien Keserempakan Perkecambahan
(KSP).
Parameter
1. Kadar air
2. Viabilitas biji
3. Daya Kecambah Total (DKT),
Koefisien Kecepatan Perkecambahan
(KCP), Koefisien Keserempakan
Perkecambahan (KSP)
4. Perkecambahan Mencapai 50% (P50)
5. Hari Pertama Berkecambah (HPB) dan
Hari Terakhir Berkecambah (HTB)
6. Pengaruh naungan terhadap
perkecambahan biji
7. Pengaruh spektrum cahaya terhadap
perkecambahan biji
HASIL DAN PEMBAHASAN
Karakter Buah dan Biji V. leucoxylon
Karakter Buah V. leucoxylon
Buah V. leucoxylon koleksi Kebun
Raya Bogor (KRB) berbentuk obovoid,
berwarna ungu-hitam (Greyed-Puple group
186A) (RHS Colour Chart, 2007), dan
berdaging dapat dilihat pada Gambar 5.
Buah berukuran panjang dengan rata-rata 9
mm ± Std. Dev 0,4418, dengan berat buah
rata-rata 0,4146 gr± Std. Dev 0,0220
Gambar 5. Karakterisasi Buah V.
leucoxylon
A. Warna Buah Berdasarkan RHS Colour
Chart. B. Panjang Buah C. Daging Buah
Karakter Biji V. leucoxylon
Biji V. leucoxylon berbentuk bulat
telur, terletak di tengah buah dengan
diselimuti daging buah. Biji V. leucoxylon
memiliki panjang rata-rata 7 mm ± Std. Dev
0,3946 dapat dilihat pada Gambar 6, berat
biji dengan rata-rata 0,1325g ± Std. Dev
0,0220 , dan tiap buah menghasilkan 1 biji
setiap buahnya.
Gambar 6. Morfologi Biji V.leucoxylon
A. Kulit Biji. B. Endosperm. C. Embrio
Biji V. leucoxylon memiliki
endosperm yang tebal, embrio terletak
ditengah-tengah biji dapat dilihat pada
Gambar 6, kulit bijinya keras dan bersifat
semipermiabel terhadap air.
Penentuan Bobot 1000 Butir Biji V.
leucoxylon
Hasil penimbangan 100 butir biji
diketahui seberat 12,992g. Hasil
penimbangan tersebut kemudian
A
B C

diekstrapolasikan untuk 1000 butir biji,
sehingga diketahui penentuan bobot 1000
butir biji V. leucoxylon seberat 129,92 g.
Pengujian Kadar Air Simpan Biji V.
leucoxylon
Kadar air biji V. leucoxylon sangat
berbeda nyata selama penyimpanan 0-6
MSP. Penyimpanan biji V. leucoxylon
dilakukan dengan 4 perlakuan simpan.
Kadar air 0-2 MSP cenderung mengalami
penurunan, tetapi pada perlakuan simpan
pada penyimpanan ruang simpan dengan
suhu 18oC menggunakan media moss kadar
air biji V. leucoxylon terlihat naik. Hal ini
menunjukkan bahwa penyimpanan
menggunakan media moss menyebabkan
biji dapat berimbibisi. Selama
penyimpanan 4-6 minggu ada
kecenderungan biji mengalami imbibisi
hingga mencapai kondisi full imbibition dan
bermetabolisme hingga siap berkecambah
dan tidak dapat disimpan lagi. Dari kadar
air yang di dapat diduga biji V. leucoxylon
memiliki sifat intermediate.
Tabel 2. Kadar Air Simpan Biji V.
leucoxylon
Pola Perkecambahan Biji V. leucoxylon
Stadia perkecambahan biji V.
leucoxylon terjadi pada waktu yang
beragam dapat dilihat pada Gambar 7.
Stadia 1 radikula tumbuh dan memecah
testa. Stadia 2 radikula memanjang dan
hipokotil tumbuh. Stadia 3 hipokotil
memanjang dan disertai plumula, akar
primer dan sekunder memanjang. Stadia 4
plumula, epikotil, kotiledon dan daun
pertama. Stadia 4 daun kedua. Stadia 5 bibit
hasil perkecambahan.
Gambar 7. Pola Perkecambahan V.
leucoxylon
A. Stadia 2 radikula memanjang dan
hipokotil tumbuh (21 hari) B. Stadia 3
hipokotil memanjang dan disertai plumula,
akar primer dan sekunder memanjang (24
hari). C. Stadia 4 plumula, epikotil,
kotiledon dan daun pertama (31 hari). D.
Stadia 4 daun kedua (34 hari).
Viabilitas Biji V. leucoxylon Pada
Perlakuan Cahaya di Rumah Kaca
Daya Kecambah Total (DKT) Perlakuan
Cahaya di Rumah Kaca (RK)
Hasil analisis uji viabilitas biji V.
leucoxylon dengan pengamatan daya
kecambah total, terlihat adanya pengaruh
cahaya terhadap perkecambahan biji.
Adapun hasil analisis uji viabilitas biji
dapat dilihat pada Tabel 3.

Tabel 3. Hasil Analisis Statistik Perlakuan
Cahaya Pada Rumah Kaca Terhadap Daya
Kecambah Total (DKT)
Keterangan: angka-angka dalam kolom yang sama dan diikuti
dengan huruf yang sama menunjukkan berbeda tidak nyata pada
taraf 95%.
Perlakuan dengan Daya Kecambah
Total (DKT) tertinggi terdapat pada
perlakuan cahaya di rumah kaca dengan
naungan paranet 55% dan daya kecambah
total terendah terdapat pada perlakuan
cahaya di rumah kaca tanpa naungan
paranet. Dari hasil analisis statistik bahwa
pengaruh perlakuan cahaya dengan
naungan 55% dan naungan 65% tidak
berbeda nyata dengan perlakuan kontrol
(tanpa naungan), kecenderungan daya
kecambah terbaik didapat pada perlakuan
cahaya dengan naungan paranet 55%.
Penelitian ini merujuk dengan
penelitian Sudomo (2009) pengaruh
naungan terhadap pertumbuhan dan mutu
bibit Manglid (Manglieta glauca bi) bahwa
naungan 55% dan naungan 65% dengan
intensitas cahaya yang kurang tetap
menghasilkan perkecambahan jika
dibandingkan dengan kontrol (naungan
0%). Intensitas yang terlalu tinggi akan
menyebabkan transpirasi terlalu besar
sedangkan intensitas yang terlalu rendah
akan menghambat fotosintesa sehingga
menghambat pertumbuhan tanaman.
Koefisien Kecepatan Perkecambahan
(KCP) Perlakuan Cahaya RK
Hasil analisis uji viabilitas biji V.
leucoxylon dengan pengamatan koefisien
kecepatan perkecambahan, terlihat tidak
adanya pengaruh cahaya terhadap
perkecambahan biji. Kuantitas cahaya
berpengaruh terhadap kecepatan
perkecambahan biji V. leucoxylon dapat
dilihat pada Tabel 4.
Tabel 4. Hasil Analisis Statistik Perlakuan
Cahaya Pada Rumah Kaca Terhadap
Koefisien Kecepatan Perkecambahan
(KCP)
Keterangan: angka-angka dalam kolom yang sama dan diikuti
dengan huruf yang sama menunjukkan berbeda tidak nyata pada taraf 95%.
Perlakuan cahaya dengan koefisien
kecepatan perkecambahan terbaik terdapat
pada perlakuan tanpa menggunakan
naungan paranet. Marjenah (2001)
melaporkan bahwa perkecambahan biji
lebih cepat pada tempat terbuka dari pada
tempat ternaung sehingga tanaman yang
ditanam pada tempat terbuka cenderung
berkecambah dengan cepat namun dalam
analisis statistik menunjukkan tidak
berbeda nyata. Akan tetapi pada titik jenuh
cahaya, tanaman tidak mampu menambah
hasil fotosintesis walaupun jumlah cahaya
bertambah. Tingginya suhu udara akan
meningkatkan laju transpirasi, hal ini antara

lain dapat ditandai dengan turunnya
kelembaban udara relatif. Apabila
berlangsung cukup lama, hal ini dapat
menyebabkan keseimbangan air tanaman
terganggu dan dapat menurunkan
perkecambahan biji tanaman.
Koefisien Keserempakan
Perkecambahan Perlakuan Cahaya RK
Hasil analisis koefisien
keserempakan perkecambahan biji V.
leucoxylon pada perlakuan cahaya di rumah
kaca dapat dilihat pada Tabel 5.
Tabel 5. Hasil Analisis Statistik Perlakuan
Cahaya Pada Rumah Kaca Terhadap
Koefisien Keserempakan Perkecambahan
(KSP)
Keterangan: angka-angka dalam kolom yang sama dan diikuti
dengan huruf yang sama menunjukkan berbeda tidak nyata pada
taraf 95%.
Berdasarkan data pada Tabel 5, dapat
dilihat bahwa nilai koefisien keserempakan
perkecambahan biji V. leucoxylon paling
tinggi pada perlakuan cahaya di rumah kaca
dalam naungan paranet 55%. Sedangkan
nilai KSP terendah ada pada perlakuan
cahaya rumah kaca tanpa naungan paranet.
Dari hasil tersebut diketahui bahwa
keserempakan perkecambahan tidak
bergantung dari kualitas cahaya yang
diterima, melainkan bergantung pada
kuantitas cahaya yang diterima oleh biji V.
leucoxylon dan keserempakan
perkecambahan diduga karena adanya
faktor luar seperti kelembaban dan suhu.
Hari Pertama Berkecambah (HPB) dan
Hari Terakhir Berkecambah (HTB)
Perlakuan Cahaya RK
Hasil analisis uji viabilitas biji V.
leucoxylon terlihat adanya pengaruh
perlakuan cahaya terhadap hari pertama
berkecambah dapat dilihat pada Tabel 6.
Tabel 6. Hari Pertama Berkecambah (HPB)
Perlakuan cahaya Rumah Kaca
Pada perlakuan cahaya rumah kaca
tanpa naungan paranet biji berkecambah
lebih cepat jika dibandingkan dengan
perlakuan dengan naungan paranet 55%
dan 65%. Daniel et al. (1997) menyatakan
bahwa terhambatnya perkecambahan
tanaman karena spektrum cahaya matahari
yang kurang merangsang aktivitas hormon
giberelin dalam proses perkecambahan biji,
terutama pada intensitas cahaya yang
rendah. Hasil analisis statistik viabilitas biji
V. leucoxylon terlihat adanya pengaruh
terhadap hari terakhir berkecambah dapat
dilihat pada Tabel 7.
Tabel 7. Hari Terakhir Berkecambah
(HTB) Perlakuan cahaya RK

Pada naungan paranet 65% terlihat
hari berkecambah terakhir lebih cepat jika
dibandingkan dengan naungan paranet
55%. Tingginya suhu udara akan
meningkatkan laju transpirasi, hal ini antara
lain dapat ditandai dengan turunnya
kelembaban udara relatif. Apabila
berlangsung cukup lama, hal ini dapat
menyebabkan keseimbangan air pada biji
terganggu dan dapat menurunkan
perkecambahan biji (Sudomo, 2009).
Viabilitas Biji V. leucoxylon Pada
Perlakuan Spektrum Cahaya di
Laboratorium
Hasil penelitian uji viabilitas biji V.
leucoxylon perlakuan cahaya di
laboratorium dengan perlakuan cahaya
spektrum merah dan cahaya spektrum
merah jauh serta perlakuan cahaya dalam
kondisi gelap dan terbuka tidak
menunjukkan adanya perkecambahan pada
perlakuan ini. Hal ini diduga bahwa biji V.
leucoxylon membutuhkan kuantitas
intensitas cahaya yang cukup untuk
melakukan perkecambahan melainkan
kualitas intensitas cahaya.
Pada umumnya cahaya yang
diperlukan oleh setiap jenis tanaman
berbeda-beda. Setiap tanaman atau jenis
pohon mempunyai toleransi yang berlainan
terhadap cahaya matahari. Ada tanaman
yang tumbuh baik di tempat terbuka,
sebaliknya ada beberapa tanaman yang
dapat tumbuh dengan baik pada tempat
teduh/bernaungan. Ada pula tanaman yang
memerlukan intensitas cahaya yang
berbeda sepanjang periode hidupnya. Pada
waktu masih muda memerlukan cahaya
dengan intensitas rendah dan menjelang
sapihan mulai memerlukan cahaya dengan
intensitas tinggi (Soekotjo,1976 dalam
Sudomo, 2009).
Karakter Simpan Biji V. leucoxylon
Uji 100 Biji Untuk Menentukan
Karakter Simpan Biji V. leucoxylon
Hasil dari uji 100 biji dengan desikasi
perlakuan silica gel dan vermiculite,
menunjukkan bahwa biji V. leucoxylon
bersifat intermediate berdasarkan dari
kadar air yang didapat dan perkecambahan
dari desikasi perlakuan silica gel dan
vermiculite dapat dilihat pada Tabel 8.
Tabel 8. Uji 100 Biji Dengan Perlakuan
Silica Gel dan Vermiculite
Menurut Gold dan Hay (2014), uji
100 biji dengan menggunakan Vermiculite
dapat mempertahankan kadar air biji sesuai
dengan kadar air aslinya. Karakter biji
rekalsitran tidak dapat bertahan dalam
pengeringan dan periode simpan yang
lama, pernyataan ini sesuai dengan hasil
yang didapatkan dari penelitian ini.
Viabilitas Biji V. leucoxylon Pada
Perlakuan Periode Simpan dan
Perlakuan Penyimpanan
Daya Kecambah Total (DKT)
Hasil analisis statistik uji viabilitas
biji V. leucoxylon dengan pengamatan daya
kecambah total terlihat tidak adanya
interaksi pada masa simpan dan perlakuan
simpan terhadap viabilitas biji dapat dilihat
pada Tabel 9.
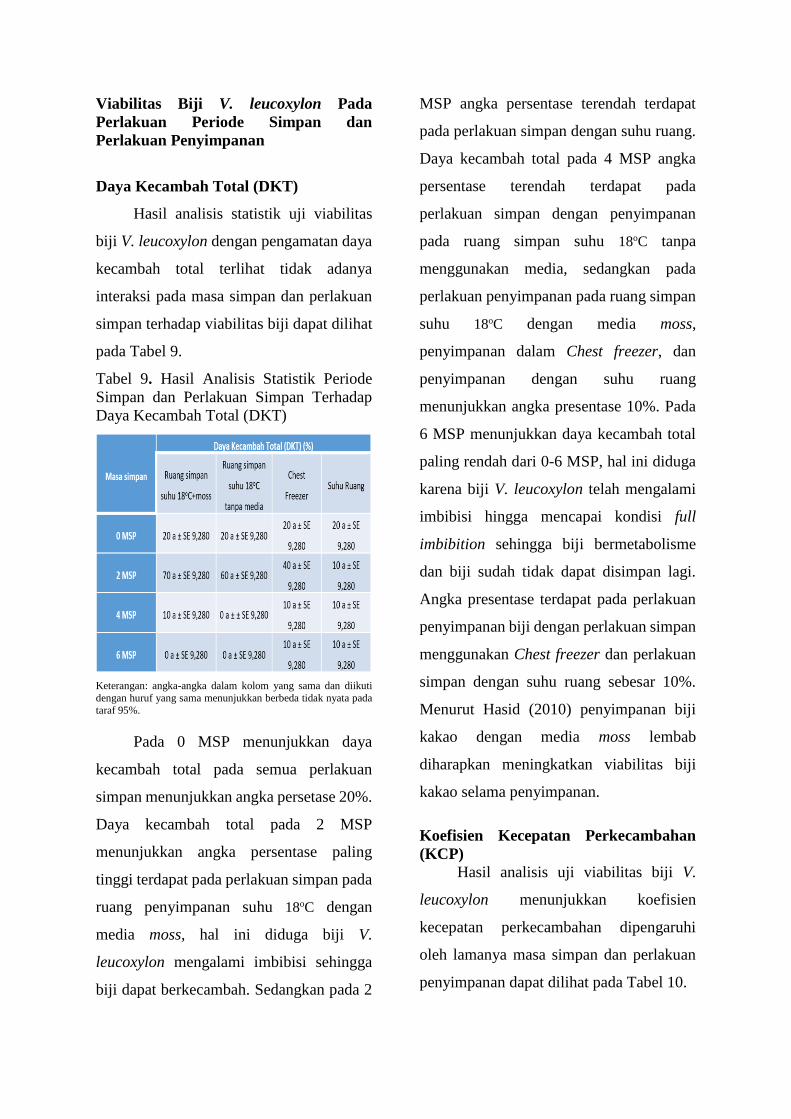
Tabel 9. Hasil Analisis Statistik Periode
Simpan dan Perlakuan Simpan Terhadap
Daya Kecambah Total (DKT)
Keterangan: angka-angka dalam kolom yang sama dan diikuti
dengan huruf yang sama menunjukkan berbeda tidak nyata pada
taraf 95%.
Pada 0 MSP menunjukkan daya
kecambah total pada semua perlakuan
simpan menunjukkan angka persetase 20%.
Daya kecambah total pada 2 MSP
menunjukkan angka persentase paling
tinggi terdapat pada perlakuan simpan pada
ruang penyimpanan suhu 18oC dengan
media moss, hal ini diduga biji V.
leucoxylon mengalami imbibisi sehingga
biji dapat berkecambah. Sedangkan pada 2
MSP angka persentase terendah terdapat
pada perlakuan simpan dengan suhu ruang.
Daya kecambah total pada 4 MSP angka
persentase terendah terdapat pada
perlakuan simpan dengan penyimpanan
pada ruang simpan suhu 18oC tanpa
menggunakan media, sedangkan pada
perlakuan penyimpanan pada ruang simpan
suhu 18oC dengan media moss,
penyimpanan dalam Chest freezer, dan
penyimpanan dengan suhu ruang
menunjukkan angka presentase 10%. Pada
6 MSP menunjukkan daya kecambah total
paling rendah dari 0-6 MSP, hal ini diduga
karena biji V. leucoxylon telah mengalami
imbibisi hingga mencapai kondisi full
imbibition sehingga biji bermetabolisme
dan biji sudah tidak dapat disimpan lagi.
Angka presentase terdapat pada perlakuan
penyimpanan biji dengan perlakuan simpan
menggunakan Chest freezer dan perlakuan
simpan dengan suhu ruang sebesar 10%.
Menurut Hasid (2010) penyimpanan biji
kakao dengan media moss lembab
diharapkan meningkatkan viabilitas biji
kakao selama penyimpanan.
Koefisien Kecepatan Perkecambahan
(KCP)
Hasil analisis uji viabilitas biji V.
leucoxylon menunjukkan koefisien
kecepatan perkecambahan dipengaruhi
oleh lamanya masa simpan dan perlakuan
penyimpanan dapat dilihat pada Tabel 10.
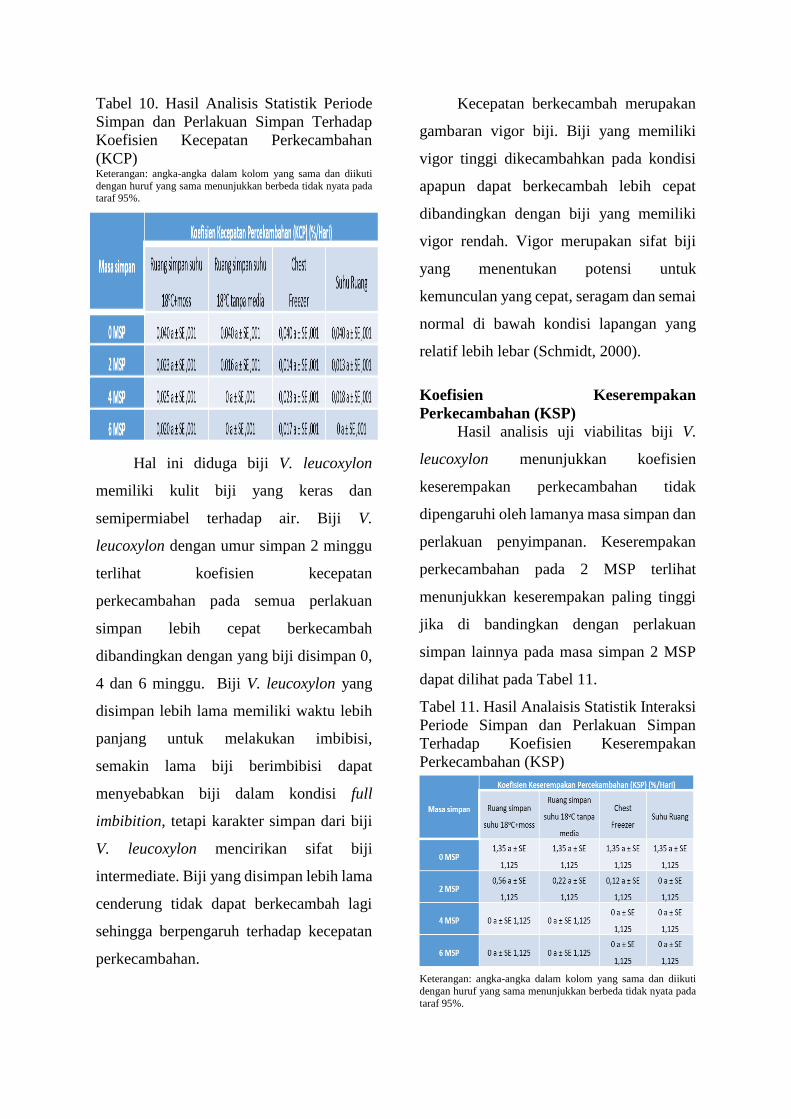
Tabel 10. Hasil Analisis Statistik Periode
Simpan dan Perlakuan Simpan Terhadap
Koefisien Kecepatan Perkecambahan
(KCP) Keterangan: angka-angka dalam kolom yang sama dan diikuti
dengan huruf yang sama menunjukkan berbeda tidak nyata pada taraf 95%.
Hal ini diduga biji V. leucoxylon
memiliki kulit biji yang keras dan
semipermiabel terhadap air. Biji V.
leucoxylon dengan umur simpan 2 minggu
terlihat koefisien kecepatan
perkecambahan pada semua perlakuan
simpan lebih cepat berkecambah
dibandingkan dengan yang biji disimpan 0,
4 dan 6 minggu. Biji V. leucoxylon yang
disimpan lebih lama memiliki waktu lebih
panjang untuk melakukan imbibisi,
semakin lama biji berimbibisi dapat
menyebabkan biji dalam kondisi full
imbibition, tetapi karakter simpan dari biji
V. leucoxylon mencirikan sifat biji
intermediate. Biji yang disimpan lebih lama
cenderung tidak dapat berkecambah lagi
sehingga berpengaruh terhadap kecepatan
perkecambahan.
Kecepatan berkecambah merupakan
gambaran vigor biji. Biji yang memiliki
vigor tinggi dikecambahkan pada kondisi
apapun dapat berkecambah lebih cepat
dibandingkan dengan biji yang memiliki
vigor rendah. Vigor merupakan sifat biji
yang menentukan potensi untuk
kemunculan yang cepat, seragam dan semai
normal di bawah kondisi lapangan yang
relatif lebih lebar (Schmidt, 2000).
Koefisien Keserempakan
Perkecambahan (KSP)
Hasil analisis uji viabilitas biji V.
leucoxylon menunjukkan koefisien
keserempakan perkecambahan tidak
dipengaruhi oleh lamanya masa simpan dan
perlakuan penyimpanan. Keserempakan
perkecambahan pada 2 MSP terlihat
menunjukkan keserempakan paling tinggi
jika di bandingkan dengan perlakuan
simpan lainnya pada masa simpan 2 MSP
dapat dilihat pada Tabel 11.
Tabel 11. Hasil Analaisis Statistik Interaksi
Periode Simpan dan Perlakuan Simpan
Terhadap Koefisien Keserempakan
Perkecambahan (KSP)
Keterangan: angka-angka dalam kolom yang sama dan diikuti
dengan huruf yang sama menunjukkan berbeda tidak nyata pada
taraf 95%.

Hal ini diduga perlakuan simpan
dengan penyimpanan pada ruang simpan
suhu 18oC dengan media moss
menyebabkan biji lebih cepat berimbibisi
sehingga biji pada masa simpan 2 MSP siap
untuk berkecambah dan menunjukan
keserempakan perkecambahan paling
tinggi diantara perlakuan simpan lainnya.
Koefisien keserempakan perkecambahan
pada perlakuan penyimpanan pada suhu
ruang menunjukkan angka terendah pada
masa simpan 2 MSP.
Masa simpan 4-6 MSP tidak
menunjukkan adanya koefisien
keserempakan perkecambahan pada biji V.
leucoxylon. Hal ini diduga biji sudah dalam
keadaan full imbibition sehingga biji harus
segera kecambahkan dan tidak dapat
disimpan lebih lama lagi. Yuniarti (2002)
menyatakan penyimpanan biji merbau
dalam ruang simpan dengan suhu 18oC
dengan media moss dapat mencegah
kemunduran viabilitas biji merbau. Pada
umumnya semakin lama biji disimpan
maka daya berkecambahnya semakin
menurun. Hal ini berkaitan dengan adanya
kemunduran kualitas benih dalam
penyimpanan. Kemunduran ini terjadi
karena selama dalam periode simpan, benih
tetap melakukan kegiatan respirasi.
Perkecambahan Mencapai 50% (P50)
Hasil analisis data menunjukkan lama
masa simpan biji dan perlakuan simpan biji
sangat berpengaruh terhadap daya
kecambah bjii mencapai 50% dapat dilihat
pada Tabel 12.
Tabel 12. Perkecambahan Mencapai 50%
(P50) Periode Simpan dan Perlakuan
Simpan
Perkecambahan biji V. leucoxylon
menunjukkan P50 hanya terdapat pada
masa simpan 2 MSP pada perlakuan simpan
pada ruang simpan suhu 18oC dengan
media moss dan perlakuan simpan pada
ruang simpan suhu 18oC tanpa media. Pada
masa simpan dengan perlakuan simpan
ruang simpan suhu 18oC dengan media
moss perkecambahan biji V. leucoxylon
mencapai 50% pada hari ke 47 setelah
tanam, sedangkan perlakuan simpan pada
ruang simpan suhu 18oC tanpa media
perkecambahan mencapai 50% pada hari ke
70. Hal ini diduga karena biji dengan
penyimpanan menggunakan media moss
biji dapat berimibibisi sehingga biji
bermetabolisme dan siap untuk
berkecambah. Byrd (1983) dalam Yuniarti
(2002) menyatakan bahwa hilangnya daya
berkecambah benih dalam penyimpanan
karena adanya respirasi yang cukup tinggi,
dengan menggunakan energi makanan yang
ada dalam sel-sel tetapi tidak mengandung
air yang cukup untuk memindahkan
jaringan simpanan makanan ke sel-sel yang
sedang melangsungkan respirasi tersebut.
Hari Pertama Berkecambah (HPB) dan
Hari Terakhir Berkecambah (HTB)
Hasil uji viabilitas biji V. leucoxylon
menunjukkan hari pertama berkecambah
dipengaruhi oleh lamanya masa simpan dan
perlakuan simpan dapat dilihat pada Tabel
13.
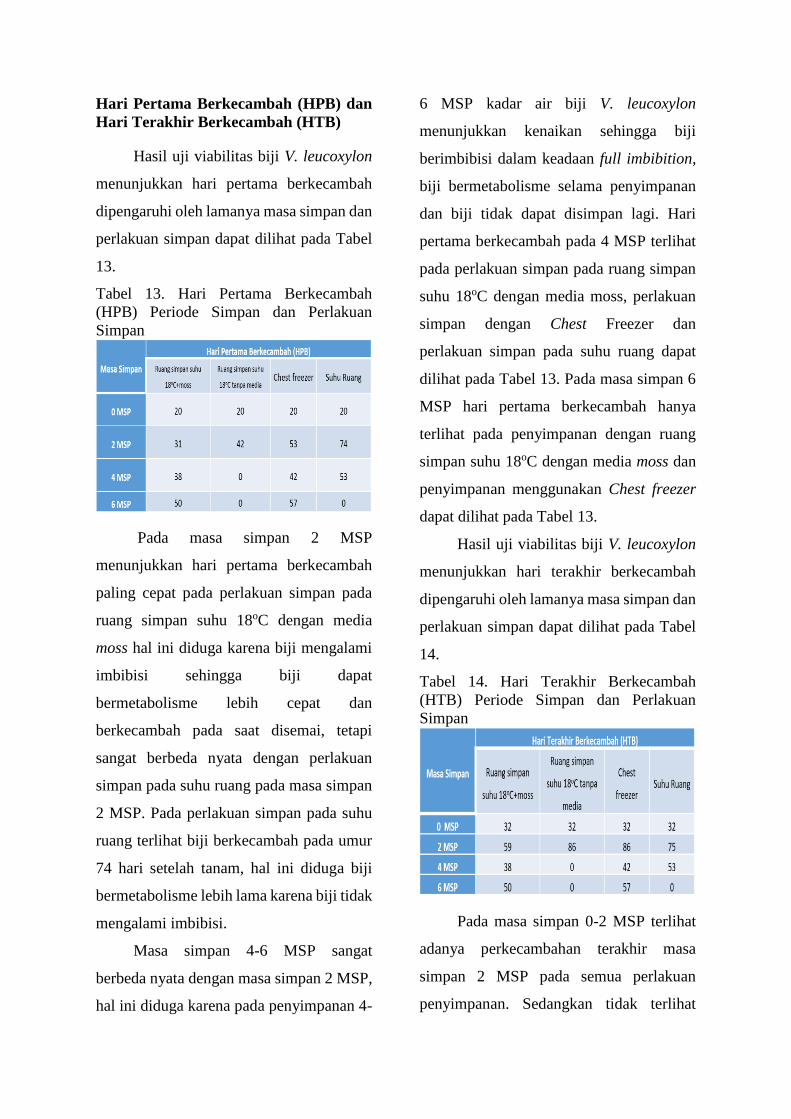
Tabel 13. Hari Pertama Berkecambah
(HPB) Periode Simpan dan Perlakuan
Simpan
Pada masa simpan 2 MSP
menunjukkan hari pertama berkecambah
paling cepat pada perlakuan simpan pada
ruang simpan suhu 18oC dengan media
moss hal ini diduga karena biji mengalami
imbibisi sehingga biji dapat
bermetabolisme lebih cepat dan
berkecambah pada saat disemai, tetapi
sangat berbeda nyata dengan perlakuan
simpan pada suhu ruang pada masa simpan
2 MSP. Pada perlakuan simpan pada suhu
ruang terlihat biji berkecambah pada umur
74 hari setelah tanam, hal ini diduga biji
bermetabolisme lebih lama karena biji tidak
mengalami imbibisi.
Masa simpan 4-6 MSP sangat
berbeda nyata dengan masa simpan 2 MSP,
hal ini diduga karena pada penyimpanan 4-
6 MSP kadar air biji V. leucoxylon
menunjukkan kenaikan sehingga biji
berimbibisi dalam keadaan full imbibition,
biji bermetabolisme selama penyimpanan
dan biji tidak dapat disimpan lagi. Hari
pertama berkecambah pada 4 MSP terlihat
pada perlakuan simpan pada ruang simpan
suhu 18oC dengan media moss, perlakuan
simpan dengan Chest Freezer dan
perlakuan simpan pada suhu ruang dapat
dilihat pada Tabel 13. Pada masa simpan 6
MSP hari pertama berkecambah hanya
terlihat pada penyimpanan dengan ruang
simpan suhu 18oC dengan media moss dan
penyimpanan menggunakan Chest freezer
dapat dilihat pada Tabel 13.
Hasil uji viabilitas biji V. leucoxylon
menunjukkan hari terakhir berkecambah
dipengaruhi oleh lamanya masa simpan dan
perlakuan simpan dapat dilihat pada Tabel
14.
Tabel 14. Hari Terakhir Berkecambah
(HTB) Periode Simpan dan Perlakuan
Simpan
Pada masa simpan 0-2 MSP terlihat
adanya perkecambahan terakhir masa
simpan 2 MSP pada semua perlakuan
penyimpanan. Sedangkan tidak terlihat

adanya perkecambahan pada hari terakhir
masa simpan 4-6 MSP, hal ini diduga
karena biji mengalami imibibisi sehingga
mencapai keadaan full imbibition sehingga
biji bermetabolisme dan tidak dapat
disimpan lebih lama. Diduga penyimpanan
biji V. leucoxylon dalam media moss
lembab dapat mempertahankan viabilitas
biji dengan memperpendek dormansi. Akan
tetapi dalam masa simpan biji cenderung
tidak dapat berkecambah lagi diduga biji
telah mengalami imbibisi sehingga biji
dalam keadaan full imbibition dan tidak
dapat disimpan lebih lama. Berkaitan
dengan hal tersebut diduga biji V.
leucoxylon memiliki sifat biji intermediate
dan sifat biji yang semipermeable terhadap
air sehingga biji yang dalam keadaan full
imbibition harus segera disemai dan tidak
dapat disimpan lagi. Biji kakao yang
memiliki sifat intermediate sangat sensitif
terhadap suhu rendah membuktikan adanya
penurunan tajam viabilitas biji. Hal itu
mengakibatkan laju penurunan viabilitas
benih berlangsung cenderung lebih cepat
pada masa lama penyimpanan.
SIMPULAN
Simpulan
1. Metode perkecambahan terbaik pada
perlakuan cahaya dalam rumah kaca
dengan naungan paranet 55% Daya
Kecambah Total (DKT) 40%, pada
perlakuan cahaya tanpa naungan
paranet Koefisien Kecepatan
Perkecambahan (KCP) 0,055%, pada
naungan paranet 65% Koefisien
Keserempakan Perkecambahan (KSP)
0,011%.
2. Perlakuan periode simpan dan
perlakuan simpan Daya Kecambah
Total (DKT) terbaik periode 2 MSP
dengan penyimpanan pada ruang
simpan suhu 18oC dengan media moss
sebesar 70%, Koefisien Kecepatan
Perkecambahan (KCP) terbaik periode
4 MSP dengan penyimpanan pada
ruang simpan suhu 18oC dengan media
moss sebesar 0,025%, Koefisien
Keserempakan Perkecambahan (KSP)
terbaik pada periode 2 MSP dengan
penyimpanan pada ruang simpan suhu
18oC dengan media moss sebesar
0,56%.
3. Karakter simpan biji V. leucoxylon
kemungkinan memiliki sifat biji
intermediate.
Saran
Untuk penelitian karakter simpan biji
V. leucoxylon perlakuan simpan dan
perlakuan cahaya sebaiknya lebih dari 6
minggu dan dilengkapi dengan metode
penyimpanan kriopreservasi. Perlakuan
pematahan dormansi (skarifikasi) biji
terhadap biji V. leucoxylon bisa dilakukan
sebelum penyemaian mengingat karakter
kulit biji V. leucoxylon yang keras.

DAFTAR PUSTAKA
Baskin C.C. & Baskin J.M. 2001. Seeds:
Ecology, Biogeography, and
Evolution of Dormancy and
Germination. Academic Press, San
Diego.
Bewley J.D. & Black M. 1994a. Physiology
and Biochemistry of Seeds in
Relation to Germination: Viability,
Dormancy, and Environmental
Control. Springer-Verlag, New
York.
Bewley J.D. & Black M. 1994b. Seeds:
Physiology of Development and
Germination. Plenum Press, New
York.
Borthwick H.A., Hendricks S.B., Toole
F.H. & Toole V.K. 1954. Action of
Light on Lettuce-seed Germination.
Botanical Gazette 115, 205-25.
Bullock J.M. 2000. Gaps and Seedling
Colonization. In: Seeds: The
Ecology Of Regeneration In Plant
Communities. Second edition (ed
M. Fenner) pp. 375-95. CABI
Publishing, Wallingford.
Carpita N.C., Naborsn M. W., Ross C. W.
& Petretic N. L. 1979. The Growth
Physics and Water Relations of
Red-light-induced germination in
Lettuce seeds: IV. Biochemical
changes in the embryonic axes of
Red- and Far-red-treated seeds.
Planta 144, 225-33.
Chaturvedi GSP, Ram C, Singh AK, Ram
P, Ingram KT, Singh BB, Singh RK,
Singh VK (1994). Carbohydrate
status of rainfed lowland ricein
relation to submergence, drought
and shade tolerance. Dalam:
Proceeding Physiology of Stress
Tolerance in Rice, Los Banos: IRRI
Philippines, pp 104-122
Chordia, M. A., R. Sangeetha Kumari and
R. Ramesh Kannan, 2010. In-Vitro
Regeneration of Plantlets from
Internodal Callus Culture of Vitex
leucoxylon L. – A Rare Medicinal
Plant. Journal of Medicinal Plants
Research Vol. 4(22), pp. 2399-
2403.
Daniel, T.W., J.A. Helms dan F.S Baker,
1997. Prinsip-prinsip Silvikultur.
Terjemahan Joko Marsono dan
Oemi Hani'in. Edisi Kedua. Gadjah
Mada University Press, Yogyakarta.
Dassanayake, M.D. 1983. A Revised
Handbook to the Flora of Ceylon.
New Delhi. The Smithsonian
Institution, and the National
Science Foundation, Washington,
D.C., by Amerind Publishing Co.
Pvt. Ltd.
Direktorat Perbenihan Tanaman Hutan.
2002. Petunjuk Teknis Pengujian
Mutu Fisik-Fisiologi Benih. Jakarta
: Departemen Kehutanan.
Gold, K dan F. Hay. 2014. Identifying
Desiccation Sensitive Seeds.
www.kew.org
Hasanah, M. 2002. Peran Mutu Fisiologik
Benih dan Pengembangan Industri
Benih Tanaman Industri. Jurnal
Litbang Pertanian, Volume 21, No.
3
Hasid, R. 2010. Keragaan dan Mutu
Fisiologi Biji Rekalsitran Kakao
(Theobroma cacao L). Pada
Berbagai Kondisi Penyimpanan.
Jurnal Hayati Vol.10
Hong, T.d., S. Linington and R.H. Ellis.
1998. Compendium of Information
on Seed Storage Behaviour, Vol. II.
Kew : Royal Botanic Gardens.

ISTA, 2007. International Rules for Seed
Testing. Edition 2007. Zurich,
Zwitzerland.
ISTA, 2010. International Rules for Seed
Testing. Edition 2010. Zurich,
Zwitzerland.
IPB. 2010. Tinjauan Pustaka Fisiologi
Benih Padi Dan Viabilitas Benih.
Hak Cipta Milik Institut Pertanian
Bogor : Bogor.
Irawanto R 2009. Peningkatan Mutu
Koleksi Biji Melalui
Manajemen Pengelolaan dan
Penelitian Biji. Prosiding
Seminar Stemcell dan Hasil-Hasil
Penelitian Ilmu Hayati.
Universitas Brawijaya. Malang.
Justice OL, LN Bass. 2002. Prinsip &
Praktek Penyimpanan Benih,
Diterjemahkan oleh Rennie Roesli.
Jakarta: PT. Raja Grafindo Persada.
Kamil, J. 1982. Teknologi Benih. Bandung:
Angkasa.
Marjenah. 2001. Pengaruh Perbedaan
Naungan di Persemaian terhadap
Pertumbuhan dan Respon
Morfologi Dua Jenis Semai
Meranti. Jurnal Ilmiah Kehutanan
”Rimba Kalimantan” Vol. 6. No. 2.
Samarinda. Kalimantan Timur. Meena A. K., Niranjan U. S., Rao M. M.,
Padhi M. M., Babu Ramesh. 2011.
A review of the important chemical
constituents and medicinal uses of
Vitex genus. Asian Journal of
Traditional Medicines Vol. 6,
Second edition.
Nabors M. W. & Lang A. 1971. The
Growth physics and water relations
of Red-light induced germination in
Lettuce seeds: I. Embryos
germinating in osmoticum. Planta
101, 1-25.
Nurwadani, P. 2008. Teknik Pembibitan
Dan Produksi Benih. Jakarta.
Pinheiro, F. & Borghetti, F. 2003. Light and
Temperature Requirements for
Germination of Seeds of Aechmea
nudicaulis (L.) Grieseba and
Streptocalyx floribundus (Martius
ex Schultes F.) Mez
(Bromeliaceae). Acta Bot. Bras. 17,
27-35.
Pons, T.L. 2000. Seed Responses to Light.
In: Seeds: The Ecology of
Regeneration in Plants
Communities (ed M. Fenner). CABI
Publishing, London.
Richards, D. & Beardsell, D. 1987. Seed
Dormancy. In: Germination of
Australian Nattive Plant Seed (ed P.
Langkamp) pp. 1-13. Inkata Press,
Melbourne.
Royal Horticultural Society (RHS). 2007.
The Royal Horticultural Society’s
Colour Chart. Fifth Edition. The
Royal Horticultural Society,
London.
Sadjad, S., Endang, M., dan Satriyas I.
1989. Parameter Pengujian Vigor
Benih. Grasindo : Jakarta.
Schmidt, L. 2000. Pedoman Penanganan
Benih Tanaman Hutan Tropis Dan
Subtropis. Departemen Kehutanan
Dan Ifsp, Jakarta.
Sopandie D, Chozin MA, Sastrosumarjo S,
Juhaeti T, Sahardi (2003) Toleransi
padi gogo terhadap naungan.
Jurnal Hayati Vol.10 Halaman : 71-
75

Sudomo, A. 2009. Pengaruh Naungan
Terhadap Pertumbuan Dan Mutu
Bibit Manglid (Manglieta glauca
BI). Jurnal Teknologi Hutan
Tanaman Vol. 2 No. 2. Halaman :
59-66.
Sutjipto Sutjipto dan Y. Widiyastuti. 2009.
Pengaruh Cara Pengeringan
terhadap Perubahan Fisikokimia
Daun Kumis Kucing (Orthosipon
stamineus Benth). Jurnal Tumbuhan
Obat Indonesia Vol. 2 (1).
Sutopo, L. 2002. Teknologi Benih. Jakarta:
PT. Raja Grafindo Persada.
Whitmore T,C, 1988. An Introduction to
Tropical Rain Forests. Oxford
University Press, Oxford.
www.indiabiodiversity.org Diakses tanggal
27-08-2016.
Yang Q. -H., Ye W. -H., Deng X., Cao H. –
L., Zhang Y. & Xu K.-Y. 2005.
Seed Germination Eco-Physiology
of Mikania micrantha H.B.K. Bot.
Bull. Acad. Sin. 46, 293-9.
Yuniarti, N. 2002. Metode Penyimpanan
Biji Merbau (Intsia bijuga O. ktze).
Jurnal Manajemen Hutan Tropika
Vol. VIII No. 2 Halaman : 89-95













![VIABILITAS BENIH KEDELAI (Glycine max [L.] Merril) V ...digilib.unila.ac.id/24628/3/SKRIPSI TANPA BAB PEMBAHASAN.pdf · abstrak viabilitas benih kedelai (glycine max [l.] merril)](https://static.fdokumen.com/doc/165x107/5d5505fa88c993ed7f8b728f/viabilitas-benih-kedelai-glycine-max-l-merril-v-tanpa-bab-pembahasanpdf.jpg)






