
MAKALAH RE Dan Aparatus Golgi Fix
62
1 BAB I PENDAHULUAN A. LATAR BELAKANG Semua sel eukariotik mempunyai retikulum endoplama (RE). Secara khas merupakan lebih dari separuh dari rata- rata total membran dari sel hewan. RE diorganizir oleh netlike labirin dari tubula bercabang dan flattened sacs (kantong yg rata) yang diperluas melalui sitosol (gambar 12-34). Tubula dan sacs saling berhubungan, dan membran mereka berkelanjutan dengan membran luar inti. jadi, ER dan membran iti membentuk lembaran berkelanjutan me-lampirkan ruang internal single, yang disebut rongga ER atau ruang ER cisternal, yang sering menempati lebih dari 10% dari volume total sel. ER memiliki peran sentral baik dalam lipid dan biosintesis protein, dan juga melayani penyaluran Ca 2+ intraseluler yang digunakan dalam respons banyak sel sinyal. Membran RE adalah tempat produksi dari semua protein transmembran dan lipid untuk sebagian besar organel sel, termasuk ER sendiri, aparatus Golgi, lisosom, endosomes, vesikel sekretorik, dan membran plasma. Membran ER juga membuat sebagian besar lipid untuk mitokondria dan membran peroxisomal. Selain itu, hampir semua protein yang akan disekresikan ke sel-eksterior tambahan yang
-
Upload
muhammad-zulhariadi -
Category
Documents
-
view
798 -
download
12
Transcript of MAKALAH RE Dan Aparatus Golgi Fix

1
BAB I
PENDAHULUAN
A. LATAR BELAKANG
Semua sel eukariotik mempunyai retikulum endoplama (RE). Secara khas
merupakan lebih dari separuh dari rata-rata total membran dari sel hewan. RE
diorganizir oleh netlike labirin dari tubula bercabang dan flattened sacs (kantong yg
rata) yang diperluas melalui sitosol (gambar 12-34). Tubula dan sacs saling
berhubungan, dan membran mereka berkelanjutan dengan membran luar inti. jadi, ER
dan membran iti membentuk lembaran berkelanjutan me-lampirkan ruang internal
single, yang disebut rongga ER atau ruang ER cisternal, yang sering menempati lebih
dari 10% dari volume total sel.
ER memiliki peran sentral baik dalam lipid dan biosintesis protein, dan juga
melayani penyaluran Ca2+ intraseluler yang digunakan dalam respons banyak sel sinyal.
Membran RE adalah tempat produksi dari semua protein transmembran dan lipid untuk
sebagian besar organel sel, termasuk ER sendiri, aparatus Golgi, lisosom, endosomes,
vesikel sekretorik, dan membran plasma. Membran ER juga membuat sebagian besar
lipid untuk mitokondria dan membran peroxisomal. Selain itu, hampir semua protein
yang akan disekresikan ke sel-eksterior tambahan yang diperuntukkan untuk lumen ER,
aparatus Golgi, atau lisosom yang awalnya dikirim ke lumen ER.
Istilah diktiosom, badan Golgi, kompleks golgi maupun aparatus Golgi,
sebenarnya semua menunjukkan organel yang sama.Organel ini dikaitkan dengan
fungsi ekskresi sel. Tidak perlu menggunakan mikroskop elektron untuk
mengamatinya, karena strukturnya dapat dilihat dengan menggunakan mikroskop
cahaya biasa. Karena memiliki fungsi yang berhubungan dengan ekskresi sel, maka
salah satu organel yang mengandung banyak diktiosom adalah ginjal. Meskipun
demikian, organel ini mempunyai struktur yang sama di hampir semua sel eukariotik,
dengan jumlah antara 10 hingga 20 buah setiap selnya.

2
Untuk mengoptimalkan
fungsi sekresi sel, badan golgi
bekerja sama dengan retikulum
endoplasma. Retikulum endoplasma
membantu proses penampungan dan
penyaluran protein ke Golgi.Protein
tersebut selanjutnya direaksikan
dengan glioksilat sehingga
terbentuk glikoprotein. Hasil reaksi
ini dibawa ke luar sel.
Kemampuan membawa hasil ke
luar sel inilah yang membuat badan Golgi mempunyai nama lain organel sekretori.
B. TUJUAN
Tujuan dari penyusunan makalah ini adalah untuk menjelaskan:
1. Sejarah penemuan Retikulum Endoplasma dan Aparatus Golgi
2. Bagian-bagian, struktur dan fungsi RE dan Aparatus Golgi
3. Mekanisme kerja RE dan Aparatus Golgi di sel.
4. Beberapa penyakit akibat disfungsi RE dan Aparatus Golgi
5.
Gambar 12-33 Bagian-bagian dari anggota sel

3
BAB II
PEMBAHASAN
A. SEJARAH PENEMUAN RETIKULUM ENDOPLASMA DAN APARATUS GOLGI
1. Retikulum Endoplasma
Retikulum berasal dari bahasa latin yang berarti “jaringan”, sedangkan kata
endoplasmik berarti “di dalam sitoplasma. RE sendiri adalah salah satu organel yang
terdapat di dalam sel, baik itu sel tumbuhan maupun sel hewan. Menurut sejarah, RE
ditemukan pada tahun 1887. Saat itu seorang ahli sel yang bernama Garnier menyelidiki
terjadinya perbedaan warna sitoplasma sel antara sel kelenjar dengan sejumlah bagian
lain dalam sitoplasma. Bagian yang berbeda warna itu seringkali diamati dan didapat
gambaran berupa struktur yang mirip seperti guratan. Bentuk inilah yang membuat
Garnier menduga, kalau organel ini ada hubungannya dengan fungsi sekresi pada sel.
Karena itulah Garnier kemudian menyebut organel ini dengan Ergatoplasma.
Ergatoplasma di sel saraf disebut dengan Badan Nissl, yang selanjutnya juga disebut
dengan Retikulum Endoplasma.
Selanjutnya seorang ahli sel yang bernama Porter, pada tahun 1945 menemukan
jala-jala yang halus pada sitoplasma fibroblas ayam. Ketika dipotong melintang dan
diamati di bawah mirksokop, jala-jala ini tampak seperti saluran buntu, dengan bentuk
yang mirip gelembung memanjang. Penelitian ini diteruskan oleh Fry Wyssling dan
Muhlethaler di tahun 1965. Mereka mengamati di bawah mikroskop elektron dan
mengambil kesimpulan bahwa terusan-terusan tersebut saling berhubungan dan saling
berkaitan di seluruh sitoplasma. RE yang berukuran besar seperti badan Nissl dapat
diamati pada mikroskop cahaya biasa, namun RE dengan ukuran yang lebih kecil harus
diamati di bawah mikroskop elektron.
2. Aparatus Golgi
Sistem endomembran pertama kali ditemukan pada 1800-an ketika ilmuwan
Camillo Golgi melihat bahwa noda tertentu selektif hanya ditandai beberapa membran
selular internal. Golgi berpikir bahwa membran intraseluler saling berhubungan, namun
kemajuan dalam mikroskop dan studi biokimia dari berbagai membran pembungkus

4
organel menegaskan bahwa organel dalam sistem endomembran adalah kompartemen
terpisah dengan fungsi tertentu. Struktur ini melakukan pertukaran bahan membran
melalui tipe khusus transportasi. Para ilmuwan akhirnya menyimpulkan bahwa sistem
endomembran mencakup retikulum endoplasma (ER), Golgi aparatus, dan lisosom.
Vesikel juga memungkinkan pertukaran komponen membran dengan membran plasma
sel. Aparatus Golgi, atau Golgi kompleks, berfungsi sebagai sebuah pabrik di mana
protein yang diterima dari ER lebih lanjut diproses dan disortir untuk transportasi ke
tujuan akhirnya mereka: lisosom, membran plasma, atau sekresi. Selain itu, seperti
disebutkan sebelumnya, glikolipid dan sphingomyelin disintesis dalam Golgi. Dalam
sel tumbuhan, aparatus Golgi lanjut berfungsi sebagai tempat di mana polisakarida yang
kompleks dari dinding sel disintesis. Aparatus Golgi demikian terlibat dalam
pengolahan berbagai konstituen seluler yang melakukan perjalanan sepanjang jalur
sekresi.
B. BAGIAN-BAGIAN RETIKULUM ENDOPLASMA DAN APARATUS GOLGI
Sistem membran menyusun struktur RE secara keseluruhan. Di sekeliling RE
terdapat semacam sitoplasma yang disebut sitosol. Struktur Retikulum Endoplasma
sendiri tersusun atas kompartemen atau ruangan-ruangan kosong dengan lapisan
membran berukuran tebal 4 nanometer. Membran ini kemudian berkaitan dengan
membran nukleus atau inti sel.
Retikulum Endoplasma juga
mempunyai ribuan ribosom yang
merupakan tempat sintesis protein dalam
sel. Bagian RE yang mengandung
ribosom disebut Retikulum Endoplasma
Kasar atau Rough Endoplasmic
Reticulum. RE kasar berfungsi untuk
mengambil protein yang telah disintesis
oleh ribosom, dan kemudian membawa protein tersebut ke bagian-bagian sel lainnya.
Karena sebagian besar protein tidak diperlukan di dalam sel, namun lebih dibutuhkan
oleh organ lain. Dua contoh protein yang dihasilkan RE kasar adalah enzim dan
hormon.

5
Adapula bagian Retikulum Endoplasma yang tidak dikelilingi oleh ribosom.
Bagian ini disebut dengan Retikulum Endoplasma Halus atau Smooth Endoplasmic
Reticulum. Fungsi dari RE halus adalah membentuk lemak dan steroid. Sel-sel dengan
RE halus akan banyak dijumpai di organ seperti hati atau hepar.
RE memiliki struktur yang mirip kantung yang tersusun berlapis-lapis. Kantung-
kantung ini disebut dengan cisternae. Retikulum Endoplasma (RE) sebenarnya
merupakan labirin membran dalam sel. Begitu banyaknya membran yang ada di RE
sehingga jumlahnya lebih dari lima puluh persen total membran dalam sel-sel
eukariotik. Sejumlah ahli menyebut bahwa RE sebenarnya merupakan perluasan
membran sel yang kemudian membentuk labirin atau kantung yang saling berhubungan.
Kantung-kantung ini berfungsi membantu gerakan substansi sel dari satu bagian sel ke
bagian sel lain.
Dengan pengamatan di bawah mikroskop elektron, struktur Aparatus Golgi
lebih mirip kantung-kantung pipih yang tersusun bertumpuk dan dibatasi membran.
Masing-masing tumpukan biasanya tersusun atas 3 hingga 7 sakulus atau kantung. Tiap
tumpukan akan tampak cembung menghadap ke inti sel dan cekung menghadap bagian
luar sel.
Bagian dalam apparatus golgi tersusun atas sisterna dengan sejumlah lubang
atau disebut dengan fenestrasi. Untuk sakulus bagian atas mempunyai fenestra di bagian
tepi. Jika membran plasma umumnya terdiri dari dua lapis sel, apparatus golgi
mempunyai struktur membran trilaminar yang lebih tipis dari plasmalema. Di
permukaan apparatus golgi tampak sejumlah vesikel-vesikel kecil dengan ukuran
diameter 40 hingga 80 nm. Vesikel-vesikel itu berhubungan dengan sakulus, diduga
berfungsi sebagai media transportasi.
Bagian cis dari apparatus golgi menerima vesikel-vesikel yang berasal dari
Retikulum Endoplasma Kasar. Vesikel ini kemudian diserap ke ruangan-ruangan di
dalam Badan Golgi. Ruangan-ruangan tersebut akan bergerak dari sisi cis menuju ke
sisi trans. Setelah itu masing-masingruangan tersebut akan memecahkan diri dan
membentuk vesikel yang lain. Dengan proses ini vesikel siap untuk disalurkan ke
bagian-bagian sel yang lain atau dikeluarkan dari dalam sel.

6
C. STRUKTUR RETIKULUM ENDOPLAMA DAN APARATUS GOLGI
1. Retikulum Endoplasma
RE mempunyai struktur mirip kumpulan kantung dengan membran yang
memanjang seperti pipa, atau gelembung yang meluas dalam sitoplasma khususnya sel
eukariot. RE terbagi menjadi dua jenis yaitu retikulum endoplasma kasar dan retikulum
endoplasma halus. Meski berbeda, namun kedua jenis retikulum endoplasma ini
menyusun sistem membran yang melingkupi hampir seluruh ruangan dalam sel. Bagian
dalam membran pada RE disebut dengan ruang sisterna (cysternal space).Sedangkan
bagian luar luar membran di sekitar RE disebut ruang sitosolik (cytololic space).
Perbedaan penampakan atau morfologi antara retikulum endoplasma kasar dan
retikulum endoplasma halus terletak pada ada dan tidaknya ribosom di sekitar membran
yang berhadapan dengan ruang sitosolik. Perbedaan lainnya adalah RE kasar
merupakan organel dengan membran yang terusun dari suatu kantong pipih yang
disebut dengan sisterna. Sedangkan RE halus merupakan organel dengan membran
berbentuk tubular.
Walau berbeda, RE kasar dan
halus akan menyatu di tempat tertentu,
karena keduanya bekerja sama dalam
melakukan berbagai aktivitas sel.
Area ER halus dimana
transportasi vesikel membawa protein
yang baru disintesis dan tunas lipid pergi
untuk transportasi ke aparatus Golgi
disebut ER transisi.
RE kasar memiliki struktur khas
yaitu setiap lembarannya tersusun atas 2 membran sel yang kemudian menjadi satu
pada bagian tepi sel masing-masing. Membran ini dibatasi oleh kantong yang berbentuk
sakulus. Bentuk dan letak sakulus bervariasi, sesuai dengan jenis, struktur dan fungsi
sel.Misalnya RE kasar yang yang berada di pankreas, sakulus menjadi tampak
sistematis, terarah, serta paralel antara satu kantung dengan kantung lainnya. Contoh
RE kasar yang lain tampak pada sel-sel glandula dari acini pankreas dan paratoide
Gambar 12.36C RE kasar dan halus tiga dimensi pada sel hati (Bruce, A. 2008)

7
terdapat pada maxilla. Hampir seluruh sakulus yang diamati di bawah mikroskop
menempati bagian basal sitoplasma. Semakin aktif sebuah sel, maka jumlah ribosom
dan sakulur akan kian banyak.
Gambar 12-36. (A) Sebuah mikrograf elektron dari ER kasar dalam sel eksokrin pankreas yang membuat dan mengeluarkan sejumlah besar enzim pencernaan setiap hari. Sitosol diisi dengan lembaran dikemas erat membran ER dipenuhi dengan ribosom. Di kiri atas adalah sebagian dari inti dan selubung inti, perhatikan bahwa membran inti luar, yang kontinu dengan ER, juga dipenuhi dengan ribosom. (B) Sebuah mikrograf elektron bagian tipis polyribosom melekat pada membran ER. Bidang bagian di beberapa tempat pemotongan melalui RE kira-kira sejajar dengan membran, memberikan gambaran dari pola rosettelike dari polyribosom.
RE halus terbetuk dari satu labirin dengan kanalikuli yang halus, saling
berhubungan, serta berinfiltrasi dalam semua sitoplasma. RE halus tidak memiliki
ribosom pada permukaan luar membrannya. Jalur yang dibuka dengan RE halus adalah
jalur nutrisi dan mineral yang berhubungan dengan mitokondria, tempat glikogen dan
juga peroksisom.Sedangkan RE kasar mempunyai sistem organisasi membran sakular
dan penuh dilingkupi dengan ribosom. RE halus merupakan jalinan tubuli-tubuli yang
saling berkaitan dan tanpa adanya ribosom.

8
Gambar 12-38 (A) Kemelimpahan RE halus di sel yang mensekresi hormon steroid -. Mikrograf elektron ini adalah pensekresian testosteron oleh sel Leydig di testis manusia. (B) Sebuah rekonstruksi tiga dimensi dari wilayah ER halus dan ER kasar dalam sel hati. ER kasar membentuk tumpukan berorientasi cisternae rata, masing-masing memiliki ruang lumenal lebar 20-30 nm. Membran ER halus ini terhubung ke cisternae dan membentuk jaringan tubulus halus 30-60 nm.
2. Aparatus Golgi
Karena strukturnya yang besar dan teratur, aparatus Golgi adalah salah satu
organel pertama yang dideskripsikan menggunakan mikroskop cahaya. Tersusun atas
kumpulan membran-kompartemen tertutup yang ditumpuk rata, disebut cisternae, yang
menyerupai tumpukan roti pita. Setiap tumpukan Golgi biasanya terdiri atas empat
sampai enam cistemae (Gambar 2.2A), meskipun beberapa flagellata uniseluler dapat
memiliki hingga 60 cisternae. Pada sel-sel hewan, antara cisternae terkait ditautkan
oleh koneksi tubular dari banyak tumpukan, sehingga membentuk sebuah kompleks
tunggal, yang biasanya terletak di dekat inti sel dan dekat dengan sentrosom. Lokalisasi
ini tergantung pada mikrotubulus. Pada percobaan jika mikrotubulus depolymerisasi,
aparatus Golgi mereorganisasi ke tumpukan cisternae yang ditemukan di seluruh
sitoplasma, berdekatan dengan situs keluar ER (Gambar 2.3).
Beberapa sel, termasuk sel-sel tumbuhan, memiliki ratusan tumpukan cisternae
Golgi yang tersebar di seluruh sitoplasma (Gambar 2.2B). Setiap tumpukan cisternae
Golgi memiliki dua permukaan yang berbeda: permukaan cis (atau permukaan masuk)
dan permukaan trans (atau permukaan keluar). Kedua permukaan cis dan trans yang
terkait erat dengan kompartemen khusus, masing-masing terdiri dari jaringan struktur
tubular dan cisternal: jaringan cis Golgi (CGN) dan jaringan trans Golgi (TGN). CGN
adalah col-pembacaan dari leburan cluster tubular vesikuler dari ER. Protein dan lipid
memasuki jaringan cis Golgi dan keluar melalui jaringan trans Golgi menuju
permukaan sel atau kompartemen lain. Kedua jaringan penting untuk menyortir protein:
protein memasuki CGN tersebut dapat bergerak maju dalam aparatus Golgi atau
dikembalikan ke ER. Demikian pula, protein keluar dari TGN seterusnya bergerak dan
diurutkan sesuai dengan tujuan mereka berikutnya: lisosom, vesikel sekretorik, atau
permukaan sel. Mereka juga dapat dikembalikan ke kompartemen sebelumnya.

9
Gambar 2.2 Aparatus Golgi. (A) rekonstruksi tiga dimensi dari mikrograf elektron aparatus Golgi dalam sel sekretorik hewan tumpukan dari permukaan cis Golgi adalah paling dekat ke ER. (B) Sebuah Irisan tipis dari mikrograf elektron menekankan zona transisi antara ER dan aparat GoIgi dalam sel hewan. (C) Sebuah mikrograf elektron dari aparat Golgi dalam sel tumbuhan (alga hijau Chlornydomonas) terlihat pada penampang. Dalam sel tumbuhan, aparatus Golgi umumnya lebih jelas dan lebih jelas terpisah dari membran intraseluler selain dalam sel hewan. (A, digambar ulang dari A. Rambourg dan Y. Clermont, Eur J. your Biol 51:189 200, 1990 Dengan izin dari Wissenschaftliche Verlagsgesellschaft, B. milik Brij J. Gupta, C. milik George Palade.)
Gambar 2.3 Lokalisasi aparatus Golgi pada hewan dan sel tumbuhan. (A) Aparatus Golgi dalam kultur fibroblast diwarnai dengan antibodi fluorescent yang mengenali protein Golgi (merah). Aparatus Golgi terpolarisasi, menghadap ke arah di mana sel berada sebelum fiksasi. (3) aparatus Golgi dalam sel tanaman yang mengekspresikan fusi protein yang terdiri dari enzim Golgi resident bergabung dengan protein fluorescent hijau. Titik oranye terang (warna palsu) adalah tumpukan Golgi. (A courtesy, John Henley dan Mark McNiven, B, milik Chris Hawes.)
Peristiwa pengolahan yang berbeda dan pemilahan muncul untuk mengambil
tempat dan memerintahkan sekuen di wilayah yang berbeda dari aparatus Golgi,
sehingga aparatus Golgi dianggap terdiri dari kompartemen diskrit ganda. Meskipun
jumlah kompartemen tersebut belum terbentuk, aparatus Golgi paling sering dipandang
terdiri dari empat daerah fungsional yang berbeda: 1) jaringan cisGolgi; 2) tumpukan
Golgi (yang dibagi lagi menjadi sub medial kompartemen dan trans kompartemen); 3)
jaringan transGolgi (Gambar 2.4 ). Protein dari ER diangkut ke kompartemen antara
ER-Golgi dan kemudian dimasukkan ke aparatus Golgi pada jaringan cis Golgi. Untuk
kemudian maju ke kompartemen medial dan trans dari tumpukan Golgi, di mana
sebagian besar kegiatan metabolisme aparatus Golgi berlangsung. Protein dimodifikasi,
lipid, dan polisakarida kemudian pindah ke jaringan trans Golgi, yang bertindak sebagai
penyortiran dan pusat distribusi, mengarahkan lalu lintas molekuler untuk lisosom,
membran plasma, atau eksterior sel.

10
Gambar 2.4. Aparatus Golgi paling sering dipandang terdiri dari empat daerah fungsional yang berbeda: 1) jaringan cisGolgi; 2) tumpukan Golgi (yang dibagi lagi menjadi sub medial kompartemen dan trans kompartemen); 3) jaringan trans Golgi.
D. FUNGSI RETIKULUM ENDOPLASMA DAN APARATUS GOLGI
Fungsi RE
RE memiliki peran sentral dalam lipid dan biosintesis protein. Membrannya
adalah tempat produksi semua protein transmembran dan lipid untuk sebagian besar
organel sel, termasuk RE sendiri, aparatus Golgi, lisosom, endosomes, vesikel
sekretorik, dan membran plasma. Membran RE membuat kontribusi besar untuk
membran mitokondria dan peroxisomal dengan memproduksi sebagian besar lipid
mereka. Selain itu, hampir semua protein yang akan disekresikan ke sel eksterior-plus
mereka yang ditakdirkan untuk lumen RE, aparatus Golgi, atau lisosom -pada awalnya
dikirim ke lumen ER.
Retikulum endoplasma kasar memiliki beberapa peran penting yaitu sebagai
berikut.
1. Glikosilasi protein
Fungsi utamanya adalah produksi dan pengolahan protein tertentu di lokasi
ribosom yang kemudian diekspor. Ribosom melakukan pekerjaan mereka dan
menghasilkan protein, yang kemudian dikirim ke retikulum endoplasma kasar, untuk

11
diproses lanjut. Penambahan kovalen gula ke protein merupakan salah satu fungsi
biosintesis utama dari RE. Protein di glikosilasi pada residu asparagin spesifik
(glikosilasi N-linked) dalam RE sementara translasi mereka masih dalam proses. Fungsi
retikulum melibatkan pembentukan dua jenis protein. Salah satunya adalah jenis yang
membentengi dan akan tertanam ke dalam membran retikulum. Jenis lain yang larut air,
yang, setelah penciptaan pada situs ribosom, melewati membran dan ke dalam lumen.
2. Folding Protein
Protein yang masuk diproses lebih lanjut dalam. Sama seperti kotak kardus dua
dimensi dilipat untuk membuat kotak, protein dilipat ke dalam bentuk tiga dimensi yang
tepat dan karbohidrat dapat ditambahkan. Banyak senyawa yang terhubung ke rantai
protein dirakit di lumen, sesuai dengan kebutuhan. Setelah lipat selesai, mereka siap
untuk pengiriman. Melipat dimungkinkan oleh kehadiran protein chaperon
(pendamping) dalam lumen. Molekul seperti hemoglobin diproduksi dalam retikulum
endoplasma kasar.
3. Transportasi protein
Fungsi lain RER adalah untuk mengangkut protein siap ke situs di mana mereka
diperlukan. Mereka juga dapat dikirimkan ke badan Golgi untuk diproses lebih lanjut,
melalui vesikel.
4. Pemeriksaan Kualitas Protein
Setelah perakitan, setiap protein dibuat dalam lumen RE dikenakan penilaian
kualitas menyeluruh. Protein diperiksa untuk penataan dan struktur yang benar dan jika
tidak sesuai dengan persyaratan yang tepat, maka ditolak, disimpan dalam lumen dan
dikirim kembali untuk didaur ulang. Banyak kondisi medis seperti Emfisema dan jenis
Cystic Fibrosis disebabkan karena penolakan terhadap protein krusial dengan sistem
memeriksa kualitas RE. Sehingga retikulum endoplasma kasar adalah bagian yang
sangat penting dari metabolisme sel, memainkan peran utama dalam fungsi setiap
jaringan sel yang berbeda.
Retikulum endoplasma halus akan banyak ditemui dalam sel berbagai organ
seperti sel otot pada rangka, tubulus pada ginjal dan juga kelenjar steroid. Protein
retikulum endoplasma halus mempunyai bermacam fungsi, seperti:
1. Proses sintesis hormon steroid, khususnya di kelenjar gonad maupun korteks ginjal
2. Proses pemurnian kembali atau detoksifikasi pada hati yang banyak mengandung
komponen organik seperti barbiturat serta etanol.

12
3. Proses pelepasan glukosa dari glukosa 6 fosfat pada hepar, maupun pelepasan gugus
gula yang lain.
4. Tempat penyimpanan glikogen dalam bentuk granula di membran luar RE halus,
yang berkaitan fungsi hepar.
Fungsi lain yang krusial dari RE pada banyak sel eukariotik adalah
mengasingkan Ca2+ dari sitosol. Perakitan Ca2+ pada sitosol dari ER dan selanjutnya
reuptake terjadi banyak rensopon cepat pada sinyal ekstraseluler. pompa transpor Ca2+
dari sitosol ke dalam lumen RE. konsentrasi tinggi pada ikatan Ca2+ protein pada ER
menfasilitasi penyimpanan Ca2+. beberapa jenis sel , dan mungkin banyak pada
lainnya, daerah spesifik dari ER dikhususkan pada penyimpanan Ca2+. di sel otot
jumlahnya melimpah, memodifikasi RE halus disebut sarkoplasmik retikulum.
pembuatan dan reuptake (penyerapan kembali) dari Ca2+ melalui sarkoplasmik
retikulum menggerakkan kontraksi miofibril dan relaksasi, berturut-turut selama
rentetan dari kontraksi otot .
Fungsi Aparatus golgi
Aparatus golgi merupakan organel
utama untuk sintesis karbohidrat, serta sortasi
(pemilihan), dan stasiun pengiriman produk
dari ER. Sel membuat banyak polisakarida di
dalam aparatus golgi, termasuk pektin dan
hemiselulosa dinding sel tumbuhan dan
sebagian besar glikosaminoglikan matriks
ekstraseluler pada hewan. Aparatus Golgi
juga ditemukan pada jalur keluar ER,
sebagian besar karbohidrat membuatnya
melekat sebagai rantai samping oligosakarida
dengan banyak protein dan lipid pada
ER. Potongan kecil (subset) dari
kelompok-kelompok oligosakarida
berfungsi sebagai tag/pengenal untuk mengarahkan proteins spesifik ke vesikel yang
kemudian mengangkut mereka ke lisosom. Setelah protein dan lipid memperoleh
oligosakarida yang tepat dalam aparatus Golgi, maka akan dikirimkan melalui
Gambar 2.1. Panah Biru menunjukkan jalur balik dari beberapa komponen sel dari aparatus golgi kembali ke ER. Panah merah menunjukkan jalur sekretori biosintetis dari ER ke aparatus golgi

13
transportasi vesikel ke bagian yang membutuhkan. Skema hubungan antara aparatus
golgi dan ER ditunjukkan Gambar 2.1.
E. MEKANISME KERJA RETIKULUM ENDOPLASMA DALAM SEL
1. Fungsi Dalam Proses Translasi dan Pascatranslasi Protein
Sel mamalia memulai mengimpor sebagian besar protein ke RE sebelum sintesis
lengkap dari rantai polipeptida yaitu, impor adalah proses kerja/co-translasi (Gambar
12-35A). Sebaliknya, impor protein ke mitokondria, kloroplas, inti, dan peroksisom
adalah proses pasca-translasi (Gambar 12-35B). Dalam co-translasi transportasi,
ribosom yang mensintesis protein adalah terpasang langsung ke membran ER,
memungkinkan salah satu ujung protein yang akan translokasi/berpindah tempat ke RE
sedangkan sisanya dari rantai polipeptida sedang dirakit.
2. Sequences Sinyal adalah Pertama Yang Ditemukan Di Protein Yang Diimpor Ke RE Kasar.
RE menangkap protein diseleksi dari sitosol ketika mereka sedang disintesis.
Protein ini terdiri dari dua jenis: protein transmembran, yang hanya sebagian
translokasi/pindah melintasi membran RE dan menjadi tertanam di dalamnya, dan
protein larut dalam air, yang sepenuhnya ditranslokasikan melintasi membran RE dan
dilepaskan ke dalam lumen RE. Beberapa protein transmembran berfungsi di RE, tetapi
banyak yang diperuntukkan untuk berada di membran plasma atau membran organel
lain. Protein -larut air diperuntukkan baik untuk sekresi atau untuk tinggal di lumen
organel. Semua protein ini, tanpa memperhatikan nasib mereka selanjutnya, diarahkan
ke membran RE oleh sekuen/ urutan sinyal RE, yang memulai translokasi mereka
dengan mekanisme umum. Sekuen sinyal (sekuen sinyal dan strategi pemilahan
Ga Fig.12-35. Co-translasi dan pascatranslasi protein translokasi (A) Ribosom berikatan dengan mem-bran RE selama Co-translasi translokasi (B) Perbedaannya, ribosom menyele-saikan sintesis protein dan melepaskan-nya pada pascatranslasi translokasi.

14
protein) pertama kali ditemukan pada awal tahun 1970 dalam sekresi protein yang
ditranslokasikan melintasi membran RE sebagai langkah pertama menuju pelepsan
akhir mereka dari sel. Dalam percobaan kunci, penyandian mRNA sekresi protein ini
diterjemahkan oleh ribosom secara in vitro. Ketika mikrosom dihilangkan dari sistem
sel-bebas, protein yang disintesis adalah sedikit lebih besar dari protein disekresikan
normal, panjang ekstra menjadi peptida yang memimpin N-terminal. Dengan
keberadaan mikrosom berasal dari RE kasar, bagaimanapun, protein dari ukuran yang
benar dihasikan. Menurut hipotesis sinyal, pemimpin adalah sekuen sinyal yang
mengarahkan protein disekresikan ke membran RE dan kemudian memutus melalui
sinyal peptidase di membran RE sebelum rantai polipeptida telah selesai (Gambar 12-
38).
3. Sinyal Pengenalan Partikel (SRP) Mengarahkan Sequences Signal RE Ke Reseptor Khusus Dalam Membran Re Kasar
Sekuen sinyal RE diarahkan ke membran RE oleh sekurang-kurangnya dua
komponen: partikel sinyal pengenalan (SRP), yang berputar/bersiklus antara membran
RE dan sitosol dan mengikat sekuen sinyal, dan sebuah reseptor SRP dalam membran
RE. SRP adalah partikel yang kompleks, yang terdiri dari enam rantai polipeptida yang
berbeda terikat dengan molekul RNA tunggal kecil (Gambar 12-39). SRP dan reseptor
Gambar 12.38 hipotesis sinyal

15
yang ditemukan di semua sel, menunjukkan bahwa mekanisme protein-target muncul di
awal evolusi dan telah dilestarikan.
Gambar 12-39 Sinyal pengenalan partikel.: (A). Mamalia SRP, (B). Ikatan SRP yang mengikat Ribosom
SRP adalah struktur rodlike yang membungkus di sekitar subunit ribosom besar, dengan
salah satu ujung mengikat urutan sinyal ER yang muncul sebagai bagian dari rantai
polipeptida yang baru dibuat dari ribosom, pada blok ujung lainnya faktor elongasi
mengikat situs yang menghubungkan antara subunit ribosom besar dan kecil (Gambar
12-39). Blok Ini menghentikan sintesis protein segera setelah sinyal peptida muncul
dari ribosom. Saat jeda sementara mungkin memberikan cukup waktu ribosom untuk
berikatan ke membran ER sebelum menyelesaikan rantai polipeptida, sehingga
memastikan bahwa protein tersebut tidak dilepaskan ke sitosol. Perangkat keamanan Ini
mungkin sangat penting untuk disekresikan dan hidrolisis lisosomal yang bisa
mendatangkan kerusakan di sitosol. Jeda tersebut juga memastikan bahwa sebagian
besar dari protein yang bisa melipat ke dalam struktur kompak tidak dibuat sebelum
mencapai Translocator dalam membran ER. Dengan demikian, berbeda dengan
pemasukan pasca-translasi pada protein ke mitokondria dan kloroplas, protein
pendamping tidak diperlukan untuk menjaga protein yangg dibentangkan.
Setelah terbentuk, kompleks SRP-ribosom mengikat reseptor SRP, yang
merupakan protein kompleks membran integral tertanam dalam membran ER kasar.
Interaksi ini membawa kompleks SRP-ribosom ke protein Translocator. SRP dan
reseptor SRP kemudian dilepaskan, dan Translocator mentransfer pertumbuhan rantai
polipeptida melintasi membran (Gambar 12-40).

16
Gambar 12-40 sekuen sinyal RE dan SRP memerintahkan ribosom ke membran RE.
Proses transfer co-translasi Ini menghasilkan dua spasial pemisah populasi
ribosom di sitosol. Membran mengikat ribosom, melekat pada sisi sitosol dari membran
ER, yang terlibat dalam sintesis protein yang sedang secara bersamaan ditranslokasi ke
RE. Ribosom bebas, tidak terikat pada membran apapun, mensintesis semua protein lain
yang disandikan oleh genom nuklir. membran terikat dan Ribosom bebas secara
struktural dan fungsional identik. Mereka hanya berbeda dalam protein yang mereka
buat pada waktu tertentu.

17
Gambar 12-41 Free dan membran-terikat ribosom. (A) Sebuah kolam umum ribosom mensintesis protein yang tinggal di sitosol dan mereka yang diangkut ke RE.
4. Translokasi Melewati Membran ER Tidak Selalu Memerlukan Pemanjangan Rantai Polipeptida Yang Berkelanjutan
Beberapa protein disintesis sepenuhnya, bagaimanapun, dimasukkan ke RE,
menunjukkan translokasi yang tidak selalu memerlukan penerjemahan yang
berkelanjutan. Untuk berfungsi pada pasca-translasi translokasi, Translocator
membutuhkan protein aksesori yang memberi makan rantai polipeptida ke dalam pori-
pori dan perjalanan translokasi (Gambar 12-44). Pada bakteri, suatu mesin penggerak
(motor) translokasi protein, SecA ATPase, menempel ke sisi sitosol dari Translocator, di
mana ia mengalami perubahan konformasi siklik didorong oleh hidrolisis ATP. Setiap
kali ATP dihidrolisis, sebagian dari sisipan protein SecA masuk ke pori Translocator,
mendorong segmen pendek dari protein Passenger dengannya. Sebagai hasil dari
mekanisme ratchet (roda bergerigi searah), protein SecA secara progresif mendorong
rantai polipeptida dari protein diangkut melintasi membran.
Gambar 12-44 Tiga cara di mana translokasi protein dapat didorong melalui struktural translocators yang mirip.
Sel eukariotik menggunakan perangkat yang berbeda dari protein aksesori yang
mengasosiasikan dengan kompleks Sec61. Protein ini menjangkau membran ER dan
menggunakan domain kecil di sisi lumenal dari membran ER untuk menyetorkan
protein pendamping seperti Hsp70 (disebut BIP, untuk mengikat protein) ke rantai
polipeptida yang muncul dari pori-pori dalam lumen ER. Siklus BIP mengikat dan

18
melepaskan translokasi gerakan searah, seperti yang dijelaskan sebelumnya untuk
mitokondria protein Hsp70 yang menarik protein melintasi membran mitokondria.
Protein yang diangkut ke RE melalui mekanisme pasca-translasi adalah pertama
kali dirilis ke sitosol, di mana mereka mengikat protein pendamping untuk mencegah
unfolding, seperti yang dibahas sebelumnya untuk protein diperuntukkan untuk
mitokondria dan kloroplas.
5. Dalam Single-Pass Protein Transmembran, Sebuah Sekuen Sinyal Intern ER Tunggal Tetap Di Dalam Lipid Bilayer Sebagai Membran Cakupan α Helix
Rangkaian sinyal dalam rantai polipeptida yang sedang tumbuh diperkirakan
memicu pembukaan pori-pori di translocator protein: setelah urutan sinyal dilepaskan
dari SRP dan rantai pertumbuhan telah mencapai panjang yang cukup, rangkaian sinyal
mengikat ke situs tertentu di dalam pori itu sendiri, sehingga membuka pori-pori.
Sebuah urutan sinyal ER kemudian dikenal dua kali: pertama oleh SRP dalam sitosol
dan kemudian oleh pengikatan daerah pada pori dari translator protein, di mana ia
berfungsi sebagai sinyal start-transfer (atau start-transfer peptida) yang membuka
pori-pori (diilustrasikan pada sebuah protein terlarut ER pada Gambar 12-45).
Pengenalan ganda mungkin membantu untuk memastikan bahwa hanya protein yang
tepat memasuki lumen RE.
Gambar 12-45 Sebuah model yang menjelaskan bagaimana protein larut ditranslokasikan melewati
membran RE.
Ketika rantai polipeptida yang baru lahir tumbuh cukup panjang, sinyal ER
peptidase memotong urutan sinyal dan melepaskan mereka dari pori-pori ke dalam
membran, di mana mereka dengan cepat terdegradasi menjadi asam amino oleh

19
protease lainnya dalam membran ER. Untuk melepaskan urutan sinyal ke membran,
Translocator telah membuka secara lateral (kesamping). Translocator kemudian
berpagar (gated) dalam dua arah: dapat membuka untuk membentuk pori-pori melewati
membran untuk melepaskan bagian hidrofilik dari protein melintasi lapisan ganda lipid,
dan dapat membuka lateral dalam membran untuk membiarkan bagian hidrofobik dari
partisi protein ke dalam lipid bilayer .
Dalam kasus yang paling sederhana, urutan sinyal N-terminal memulai
translokasi, hanya sebagai protein terlarut, tapi segmen hidrofobik tambahan dalam
rantai polipeptida menghentikan proses transfer sebelum rantai polipeptida seluruh
ditranslokasikan. Sinyal stop transfer ini menyatukan protein dalam membran setelah
sekuens sinyal ER (sinyal start-transfer) telah dibebaskan dari Translocator dan telah
dipotong (Gambar 12-46). Mekanisme gating lateralis mentransfer urutan stop-transfer
ke bilayer, dan tetap ada sebagai segmen α heliks tunggal rentangan membran , dengan
ujung N-protein pada sisi lumenal membran dan C-terminal pada sisi sitosol.
Gambar 12-46 Bagaimana single-pass protein transmembran dengan urutan sinyal ER dibelah diintegrasikan ke dalam membran ER.
Dalam dua kasus lain, rangkaian sinyal adalah internal, bukan di ujung N-
terminal dari protein. Seperti N-terminal rangkaian sinyal ER, SRP juga mengikat ke
urutan sinyal internal. SRP membawa ribosom membuat protein ke membran ER dan
berfungsi sebagai sinyal start-transfer yang memulai translokasi protein. Setelah
dibebaskan dari Translocator tersebut, urutan start- transfer internal tetap dalam lapisan
lipid ganda sebagai alfa heliks bentangan membran tunggal.

Gambar 12-51. Glikosilasi protein dalam RE kasar. setelah ada rantai polipeptida yang memasuki lumen RE, terjadi glikosilasi dengan target asam amino asparagin
20
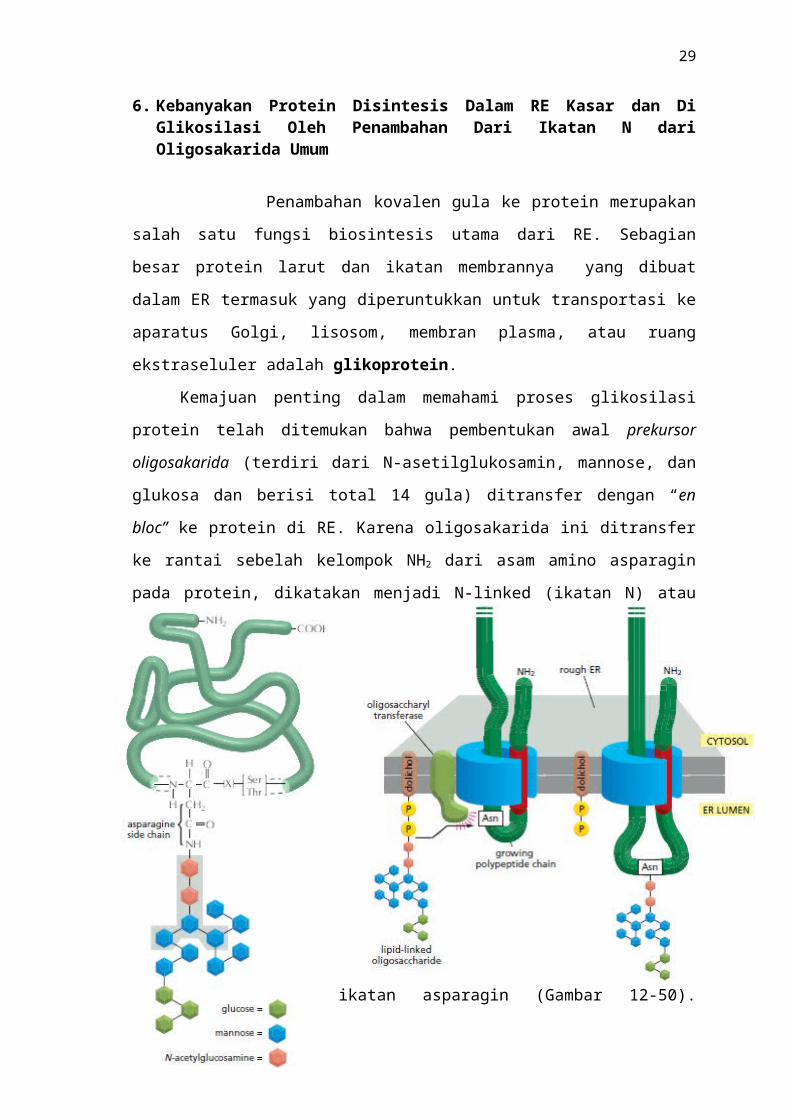
6. Kebanyakan Protein Disintesis Dalam RE Kasar dan Di Glikosilasi Oleh Penambahan Dari Ikatan N dari Oligosakarida Umum
Penambahan kovalen gula ke protein merupakan salah satu fungsi biosintesis
utama dari RE. Sebagian besar protein larut dan ikatan membrannya yang dibuat dalam
ER termasuk yang diperuntukkan untuk transportasi ke aparatus Golgi, lisosom,
membran plasma, atau ruang ekstraseluler adalah glikoprotein.
Kemajuan penting dalam memahami proses glikosilasi protein telah ditemukan
bahwa pembentukan awal prekursor oligosakarida (terdiri dari N-asetilglukosamin,
mannose, dan glukosa dan berisi total 14 gula) ditransfer dengan “en bloc” ke protein di
RE. Karena oligosakarida ini ditransfer ke rantai sebelah kelompok NH2 dari asam
amino asparagin pada protein, dikatakan menjadi N-linked (ikatan N) atau ikatan
asparagin (Gambar 12-50). Transfer dikatalisis oleh kompleks enzim ikatan membran-,
sebuah oligosaccharyl transferase, yang memiliki situs aktif terpapar di sisi lumenal di
membran RE, ini
menjelaskan mengapa protein sitosol tidak
glikosilasi dengan cara ini. Sebuah molekul
Gambar 12–50 Asparagine-linked (Ikatan N) pendahulu oligosaccharide yang ditambahkan pada kebanyakan pro- tein di membran RE kasar

21
khusus yang disebut lipid dolichol memegang prekursor oligosakarida dalam membran
RE. Dia mentransfer rantai oligosakarida kepada asparagin target dalam langkah
enzimatik tunggal segera setelah asam amino telah mencapai lumen RE selama
translokasi protein (Gambar 12-51). Satu salinan dari oligosaccharyl transferase
dihubungkan dengan tiap Translocator protein, yang mengijinkannya untuk memindai
dan glycosylate rantai polipeptida yang masuk secara efisien.
Gambar 12-52. Sintesis dari ikatan lipid prekursor dari oligosakaridae pada membran RE kasar
Ikatan N dalam oligosakarida yang jauh oligosakarida yang paling umum,
ditemukan pada 90% dari semua glikoprotein. jarang sekali, oligosakarida ini
dihubungkan dengan kelompok hidroksil pada rantai samping dari suatu serin, treonin,
atau asam amino hidroksilin. Ikatan O pada oligosakarida terbentuk dalam aparatus
Golgi.
7. Oligosakarida Digunakan Sebagai Kata Kunci Sebagai Penanda Protein Yang Melipat (Protein Folding)

22
Salah satu pengamatan yang sangat membingungkan adalah bahwa beberapa
protein membutuhkan ikatan N glikosilasi untuk melipat dengan tepat di ER, namun
lokasi yang tepat dari oligosakarida menempel permukaan protein tampaknya tidak
menjadi masalah. Sebuah petunjuk untuk peran glikosilasi dalam pelipatan protein
berasal dari studi dua pendamping protein ER, yang disebut calnexin dan calreticulin
karena mereka membutuhkan Ca2 + untuk kegiatan mereka. chaperone/pendamping Ini
adalah ikatan karbohidrat protein, atau lektin, yaitu yang mengikat oligosakarida dengan
lipatan protein tidak lengkap dan mempertahankan mereka di RE. Seperti chaperones
lain, mereka mencegah protein tidak sempurna melipat agar tidak menjadi ireversibel
agregat (gabungan yang tidak dapat dirubah). Baik calnexin dan calreticulin juga
mendorong hubungan antara protein tidak sempurna dilipat dengan chaperone RE yang
lain, yang mengikat sistein yang belum terbentuk ikatan disulfida.
Gambar 12-53. Peranan N-linked (ikatan N) glikolisasi dalam proses folding protein di RE.
Calnexin dan calreticulin mengenali N-linked oligosakarida yang mengandung
glukosa terminal tunggal, dan kemudian mengikat protein setelah dua dari tiga glucoses
pada prekursor oligosakarida yang telah dihapus oleh glukosidase RE. Ketika glukosa
ketiga telah dihapus, protein dipisahkan dari caperon dan dapat meninggalkan RE.
Enzim lain RE, sebuah transferase glucosyl yang terus menambahkan glukosa
kepada oligosakarida yang telah kehilangan glukosa terakhir mereka. Dengan demikian,
sebuah protein yang dibentangkan mengalami siklus terus menerus pada pemotongan

23
glukosa (oleh glukosidase) dan penambahan glukosa (oleh transferase glycosyl),
mempertahankan ketertarikan untuk calnexin dan calreticulin sampai telah mencapai
keadaan sepenuhnya melipat (Gambar 12-53).
8. Protein Yang Tidak Dilipat Akan Ekspor Dari RE Dan Terdegradasi Di
Sitosol
Meskipun semua bantuan dari chaperones, banyak molekul protein (lebih dari
80% untuk beberapa protein) translokasi ke RE gagal untuk melipat dengan tepat atau
tempat oligomer. Protein tersebut diekspor dari belakang RE ke sitosol, di mana mereka
terdegradasi. Misalnya, seperti translokasi dalam mitokondria atau kloroplas, protein
chaperone mungkin diperlukan untuk menjaga rantai polipeptida dalam keadaan tidak
dilipat sebelum dan selama transportasi. Demikian pula, sumber energi yang dibutuhkan
untuk memberikan pengarahan untuk transportasi dan untuk menarik protein ke dalam
sitosol. Akhirnya, sebuah Translocator, mungkin terdiri dari beberapa komponen yang
sama yang digunakan untuk meneruskan transportasi ke RE (seperti Sec61), diduga
diperlukan.
Gambar 12-54. Proses ekspor dan degradasi dari protein gagal melipat di RE.
Memilih protein dari ER untuk degradasi adalah proses yang unik. Protein yang
gagal melipat atau subunit protein dirakit harus terdegradasi. Membantu dalam
membuat perbedaan ini berasal dari ikatan N oligosakarida, yang berfungsi sebagai
timer yang mengukur berapa lama protein telah menghabiskan di RE. Pemotongan
Lambat dari mannose tertentu pada pohon inti-oligosakarida oleh suatu enzim

24
(mannosidase) di ER diperkirakan membuat struktur baru oligosakarida bahwa
peralatan retrotranslocation dikenal. Protein yang melipat dan keluar dari ER lebih
cepat daripada aksi dari mannosidase karena itu akan lolos degradasi.
Setelah protein gagal melipat akan diretrotranslokasikan ke sitosol, N-glycanase
menghilangkan rantai oligosakarida yang en bloc. Polipeptida deglycosylasi dengan
cepat diubiquitylasi oleh ikatan ER enzim konjugasi ubiquitin- dan kemudian
dimasukkan ke dalam proteasomes di mana dia didegradasi (Gambar 12-54).
9. Kegagalan Protein Melipat Di RE Mengaktifkan Suatu Respon
Akumulasi protein yang gagal melipat di sitosol, misalnya, memicu respon heat-
shock (syok panas) , yang merangsang transkripsi gen penyandi chaperone sitosolik
yang membantu untuk melipat kembali protein. Demikian pula, sebuah akumulasi
protein yang gagal melipat di ER memicu respon protein yang gagal melipat, yang
mencakup peningkatan transkripsi gen penyandi chaperone ER, protein yang terlibat
dalam retrotranslocation dan degradasi protein dalam sitosol, dan banyak protein lain
yang membantu untuk meningkatkan kapasitas protein folding dari ER.
Bagaimana sinyal protein yang gagal melipat dalam ER bisa ke sampai ke inti?
Ada tiga jalur paralel yang menjalankan respon protein yang gagal melipat (Gambar 12-
55A). Jalur pertama, yang awalnya ditemukan dalam sel ragi, sangat luar biasa. Protein
yang gagal melipat di ER mengaktifkan kinase transmembran protein di ER, sehingga
terjadi oligomerisasi dan memfosforilasi sendiri dengan katalis kinase. Oligomerisasi
dan autofosforilasi mengaktifkan sebuah domain endoribonuclease di bagian sitosol
dari molekul yang sama, yang memotong sebuah molekul RNA spesifik sitosol pada
dua posisi, memotong intron. Ekson kemudian bergabung oleh ligase RNA kemudian
menjadi mRNA, yang diterjemahkan untuk menghasilkan gen aktif yang meregulasi
protein. Protein ini mengaktifkan transkripsi gen pengkode protein memediasi respon
protein unfoding /tidak melipat(Gambar 12-55b).
Protein yang gagal melipat juga mengaktifkan kinase transmembran kedua di
ER, yang menghambat faktor inisiasi terjemahan oleh fosforilasi dan dengan demikian
mengurangi produksi protein baru di seluruh sel.

25
Gambar 12-55 Respon terhadap protein yang tidak melipat. (A) Melalui tiga jalur paralel sinyal intraseluler, akumulasi protein yang gagal melipat di lumen ER mengirim sinyal ke inti untuk mengaktifkan transkripsi gen yang menyandi protein yang membantu sel untuk mengatasi kelimpahan protein yang gagal melipat di RE. (B) Mengatur splicing/penyambungan mRNA adalah tombol kunci peraturan dalam Pathway 1 dari respon protein tidak dilipat.

26
Salah satu konsekuensi dari pengurangan dalam terjemahan protein adalah untuk
mengurangi fluks dari protein ke RE, sehingga membatasi beban protein yang perlu
dilipat di sana. Beberapa protein, bagaimanapun, adalah istimewa diterjemahkan ketika
faktor inisiasi translasi langka (lihat hal. 490), dan salah satunya adalah gen pengatur
protein yang membantu mengaktifkan transkripsi gen pengkode protein aktif dalam
respon protein yang tidak dilipat.
Akhirnya, pada tahap ketiga , gen pengatur protein ini awalnya disintesis
sebagai protein membran ER terpadu. Karena secara kovalen ditambatkan ke membran,
tidak dapat mengaktifkan transkripsi gen dalam inti. Ketika protein yang gagal melipat
menumpuk di ER, protein transmembran diangkut ke aparatus Golgi, di mana ia
bertemu protease yang memotong domain sitosol, yang sekarang dapat bermigrasi ke
nukleus dan membantu mengaktifkan transkripsi gen pengkode protein yang terlibat
dalam respon pada masalah protein yang tidak melipat.
10. RE Merakit Sebagian Besar Lipid Bilayers
Membran RE mensintesis hampir semua kelas utama lipid, termasuk fosfolipid
dan kolesterol, diperlukan untuk produksi membran sel baru. Fosfolipid utama dibuat
oleh fosfatidilkolin (juga disebut lesitin), yang dapat dibentuk dalam tiga langkah dari
kolin, dua asam lemak, dan gliserol fosfat (Gambar 12-57). Karena asam lemak yang
tidak larut dalam air, mereka digiring dari situs mereka untuk mensintesis ke RE
melalui ikatan asam lemak protein dalam sitosol. Setelah tiba di membran ER dan
aktivasi dengan CoA, transferases asil berurutan menambahkan dua asam lemak untuk
fosfat gliserol untuk menghasilkan asam phosphatidic. Asam phosphatidic cukup larut
air untuk tetap dalam lipid bilayer, dan tidak dapat diekstraksi dari bilayer oleh ikatan
protein asam lemak. Oleh karena itu ini adalah langkah pertama yang memperbesar
lapisan ganda lipid RE.
Karena sintesis fosfolipid berlangsung pada paruh sitosol dari lapisan ganda
lipid ER, perlu ada mekanisme yang mentransfer beberapa molekul fosfolipid yang baru
dibentuk untuk selebaran lumenal dari bilayer. Dalam lipid bilayers sintetik, lipid tidak
"flip-flop" dengan cara ini. Di RE, bagaimanapun, fosfolipid menyeimbangkan
persimpangan membran. Gerakan antar-bilayer cepat dimediasi oleh Translocator
fosfolipid buruk disebut scramblase, yang menyeimbangkan fosfolipid antara dua
selebaran dari lapisan ganda lipid "flip flop-." (Gambar 12-58) . Dengan demikian,
berbagai jenis fosfolipid yang dianggap merata antara dua selebaran dari membran RE.

27
Membran plasma dan membran aparatus Golgi, lisosom, dan endosomes semua
merupakan bagian dari sistem membran yang berkomunikasi dengan RE melalui
vesikel transportasi, yang mentransfer baik protein dan lipid. Mitokondria dan plastida,
tidak termasuk sistem ini, karena itu mereka membutuhkan mekanisme yang berbeda
untuk mengimpor protein dan lipid untuk pertumbuhan. Kita telah melihat bahwa
mereka mengimpor sebagian besar protein mereka dari sitosol.
Gambar 12–58 Peranan phospholipid translocators pada synthesis lipid bilayer. (A) karena new lipid molecules di masukkan hanya ke cytosolic setengah dari molecules bilayer and lipid berputar secara spontan dari sebuah monolayer ke lainnya, sebuah ikatan membran phospholipid translocator (disebut scramblase) dibutuhkan untuk transfer molecules lipid dari setengah cytosolic menuju separuh lumenal sehingga membrane tumbuh sebagai bilayer. Scramblase tidak specific sebagai keterangan ujung grup phospholipid and kemudian perbedaan berimbang antara phospholipids dengan dua monolayers. (B) Bahan energi melalui hydrolysis ATP, sebuah ujung specific grup flippase pada membrane plasma mengaktifkan perputaran phosphatidylserine and phosphatidyl-ethanolamine diarahkan dari extracellular ke selebaran cytosolic, membuat characteristically asymmetric lipid bilayer dari membrane plasma pada cells animal .
F. BERBAGAI GANGGUAN ATAU PENYAKIT AKIBAT DISFUNGSI RE
Protein yang tidak melipat (unfolding) dan kondisi lain seperti hipoksia dan
hipoglikemia, dan paparan racun mempengaruhi homeostasis endoplasma retikulum
(RE) menyebabkan RE stres. Sel bereaksi terhadap stres RE dengan aktivasi respon
protein yang tidak melipat (UPR), yang menginduksi perubahan besar dalam

28
metabolisme sel, termasuk pelemahan umum translasi, peningkatan regulasi transkripsi
dari gen molekul pendamping, dan aktivasi RE-terkait degradasi. Namun, hasil stress
berkepanjangan atau RE akut menghasilkan kematian sel. Kemajuan terbaru
menunjukkan bahwa stress ER dan UPR memainkan peran kunci dalam respon
kekebalan tubuh, diabetes, pertumbuhan tumor dalam kondisi hipoksia, dan dalam
beberapa penyakit neurodegenerative.
1. Respon stres RE dan protein unfolding terhadap respon imun
Selain memiliki peran dalam menghilangkan stres RE, UPR juga digunakan
untuk sintesis sejumlah besar protein dalam RE oleh beberapa sel sekretori. Sebagai
contoh, antibodi-mensekresi sel plasma memanfaatkan komponen UPR untuk
diferensiasi selular dan upregulation sintesis protein. Dalam experiments transplantasi
sel vivo menunjukkan bahwa ketika Ire1 sel-sel hematopoietik dapat tumbuh menjadi
sel normal, pro-B mereka gagal untuk berkembang lebih lanjut ke dalam sel pre- B,
yang menunjukkan bahwa Ire1 diperlukan untuk diferensiasi sel B progenitor tetapi
tidak penting untuk hematopoiesis awal. Selain itu, over ekspresi dari pnyambungan
membentuk Xbp1 (Xbp1s) tidak mengembalikan diferensiasi sel pro-B, memberi kesan
bahwa IRE1 lain diatur protein yang diperlukan untuk diferensiasi sel-sel.
Tahap akhir dari diferensiasi sel plasma juga membutuhkan jalur IRE1. Plasma
sel dengan mutasi dominant negative baik dalam kinase atau endoribonuclease tersebut
domain dari IRE1, daerah terbukti diperlukan untuk splicing XBP1, gagal untuk
mengeluarkan antibodi secara efisien. Meskipun ekspresi Xbp1s tidak cukup untuk
diferensiasi sel pro-B, ekspresi ektopik dari Xbp1s mengembalikan sepenuhnya fungsi
antibodi penghasil plasma sel ini. Selain itu, ekspresi dominan- negatif eIF2a tidak
mengganggu diferensiasi sel plasma atau produksi antibodi, menyarankan keterlibatan
selektif cabang IRE1 dari UPR dalam diferensiasi sel B. Bahkan, untuk menghambat
degradasi protein dan kematian sel sekaligus memaksimalkan translasi dan folding
protein, jalur PERK direpresi dalam plasma sel melalui p58IPK, regulator negatif dari
aktivitas kinase PERK yang tidak langsung diregulasi oleh XBP1
2. Respon Stress ER dan protein yang tidak melipat terhadap diabetes
Diabetes adalah penyakit metabolik umum disebabkan oleh gagal pankreas
dikaitkan dengan otoimun (tipe I) atau resistensi insulin pada jaringan perifer (tipe II).
DM tipe I bisa terjadi akibat insulin mutan gagal melipat yang menyebabkan stres RE
kronis pada sel b-pankreas, seperti pada tikus Akita. Deregulasi dari penekanan
29
translasi menyebabkan kelebihan protein juga menyebabkan mengetik diabetes I,
seperti yang ditunjukkan oleh tikus dan manusia membawa mutasi di cabang PERK dari
UPR.
Kelebihan aktivasi dari cabang PERK dari sinyal UPR juga dapat menyebabkan
stres RE dalam sel b-pankreas dan mengakibatkan diabetes. Derepression pelemahan
translasi diperlukan oleh pemulihan stres RE dan dimediasi oleh p58IPK, kinase PERK
inhibitor. Baru-baru ini, tikus KO p58IPK juga telah ditemukan menunjukkan gejala
diabetes tipe I (Tabel 2), termasuk kadar glukosa darah tinggi dan kadar insulin
menurun seiring dengan apoptosis pankreatik yang sel b islet.
3. RE stres dan respon protein unfolding pada kanker
Stres RE dan UPR telah diamati pada tumor, tapi bagaimana aktivasi UPR
kontribusi untuk kelangsungan hidup sel kanker tidak jelas. Baru-baru ini, telah
dilaporkan bahwa UPR diaktifkan oleh hipoksia, suatu kondisi sering ditemukan di inti
tumor padat. Secara In vitro, hipoksia telah terbukti menaikkan meregulasi splicing
Xbp1 dan transkripsi pada banyak yang terkait protein UPR, termasuk BIP dan GRP94
(keluarga protein ER HSP90). sel yang kekurangan XBP1 menunjukkan kepekaan
terhadap hipoksia yang berkorelasi terbalik dengan jumlah XBP1 ekspresi, dan
menghasilkan tumor yang lebih kecil ketika ditanamkan ke tikus SCID.
G. MEKANISME KERJA DARI APARATUS GOLGI PADA SEL
1. Proses pembentukan rantai oligosakarida
Satu spesies N-linked oligosakarida terpasang en bloc untuk banyak protein di
ER dan kemudian dipangkas ketika protein masih di ER. Oligosakarida Intermediet
yang dihasilkan oleh reaksi pemangkasan berfungsi untuk membantu protein melipat
dan untuk membantu transportasi protein yang gagal melipat ke sitosol untuk degradasi.
Dengan demikian, mereka memainkan peran penting dalam mengontrol kualitas protein
keluar dari ER. Setelah fungsi ER telah dipenuhi, sel bebas untuk mendesain ulang
oligosakarida untuk fungsi baru. Hal ini terjadi dalam aparatus Golgi, yang
menghasilkan struktur oligosakarida heterogen terlihat pada protein matang. Setibanya
di CGN, protein melewati cis Golgi jaringan, sebelum memasuki pertama dari
pemrosesan kompartemen Golgi (cis Golgi cisternae). Mereka kemudian pindah ke
kompartemen berikutnya (cisternae medial) dan akhirnya ke cisternae trans, di mana

30
glikosilasi selesai. Lumen cisternae trans dianggap berhubungan dengan TGN, tempat
di mana protein dipisahkan ke dalam paket transportasi yang berbeda dan dikirim ke
tujuan akhir mereka. Langkah-langkah pengolahan oligosakarida terjadi dalam urutan
terorganisir dalam tumpukan Golgi, dengan masing-masing cisterna mengandung
kelimpahan karakteristik enzim pengolahan. Protein yang dimodifikasi berturut-turut
dalam tahap ketika mereka bergerak dari cisterna ke cisterna di tumpukan Golgi,
sehingga tumpukan membentuk unit pengolahan banyak tahap/fase.
Kompartementalisasi ini mungkin tampak tidak perlu, karena masing-masing enzim
pengolahan oligosakarida dapat menerima glikoprotein sebagai substrat hanya setelah
diproses dengan benar oleh enzim sebelumnya. Meskipun demikian, jelas bahwa proses
terjadi dalam tata ruang serta urutan biokimia: enzim katalisator langkah-langkah
pengolahan awal terkonsentrasi pada cisternae di permukaan cis tumpukan Golgi,
sedangkan langkah-langkah pengolahan berikutnya enzim katalisator terkonsentrasi di
cisternae pada permukaan trans. Ketika lumen ER penuh protein resident lumenal
terlarut dan enzim, semua protein resident dalam aparatus Golgi terikat membran.
Reaksi enzimatik dalam aparatus Golgi tampaknya dilakukan sepenuhnya pada
permukaan membran. Semua Golgi glycosidases dan transferases glycosyl adalah
protein transmembran single-pass, yang banyak diatur dalam kompleks multi-enzim.
Dengan demikian, kedua organel dalam jalur biosintesis-sekresi, ER dan aparatus
Golgi, diatur dalam cara yang berbeda.
Gambar 2.5
Pengolahan oligosakarida dalam kompartemen Golgi. Lokalisasi setiap proses ditunjukkan ditentukan oleh kombinasi teknik, termasuk subtractionasi biokimia dari membran aparatus GoIgi dan mikroskop elektron setelah pewarnaan dengan antibodi spesifik untuk beberapa enzim pengolahan. Enzim Pengolahan tidak terbatas pada cisterna tertentu, melainkan, distribusi mereka dinilai melintasi tumpukan-seperti yang enzim awal yang berperan terdapat pada sebagian besar cisternae cis GoIgi dan enzim yang bertindak sesudahnya ditemukan sebagian besar di cisternae trans Golgi.

31
2. Proses Glikosilasi Protein berlangsung dalam aparatus Golgi
Gambar 2.6 Pengolahan oligosakarida terikat N dalam Golgi. Oligosakarida terikat N dari glikoprotein diangkut dari ER untuk lebih lanjut dimodifikasi oleh reaksi yang berurutan di aparatus Golgi
Pengolahan Protein dalam Golgi melibatkan modifikasi dan sintesis dari bagian
karbohidrat dari glikoprotein. Salah satu aspek utama dari proses ini adalah modifikasi
dari N-linked oligosakarida yang ditambahkan ke protein di ER. Protein yang diubah
dalam ER dengan penambahan oligosakarida yang terdiri dari 14 residu gula. Residu
Tiga glukosa dan satu mannose kemudian dihapus sementara polipeptida masih di ER.
Setelah transportasi ke aparatus Golgi, N-linked oligosakarida dari glikoprotein tunduk
pada modifikasi lebih lanjut yang luas.
N-linked oligosakarida diproses dalam aparatus Golgi dalam memerintahkan
urutan reaksi (Gambar 9.24). Modifikasi pertama protein adalah penghapusan tiga
residu mannose tambahan. Hal ini diikuti dengan penambahan berurutan dari N-
asetilglukosamin, penghapusan dua mannoses, dan penambahan fucose dan dua lagi N-
acetylglucosamines. Akhirnya, tiga galaktosa dan tiga residu asam sialat ditambahkan.
Glikoprotein yang berbeda dimodifikasi untuk luasan yang berbeda selama perjalanan
mereka melalui Golgi, tergantung pada kedua struktur protein dan jumlah enzim

32
pengolahan yang hadir dalam kompleks Golgi dari berbagai jenis sel. Akibatnya,
protein dapat muncul dari Golgi dengan berbagai N-linked oligosakarida berbeda.
Pengolahan oligosakarida terikat N pada protein lisosomal berbeda dengan
protein yang disekresikan membran plasma. Awal tidak melakukan penghapusan tiga
residu mannose, protein ditujukan untuk dimasukkan ke dalam lisosom yang
dimodifikasi oleh fosforilasi mannose. Pada langkah pertama dari reaksi ini, N-
asetilglukosamin fosfat yang ditambahkan ke residu mannose tertentu, mungkin saat
protein masih dalam jaringan cis Golgi (Gambar 2.7). Hal ini diikuti dengan
penghapusan kelompok N-asetilglukosamin, meninggalkan mannose-6-fosfat residu
pada N-linked oligosakarida. Karena modifikasi ini, residu ini tidak dihapus selama
proses lebih lanjut. Sebaliknya, residu ini mannose terfosforilasi secara khusus diakui
oleh reseptor mannose-6-fosfat di jaringan trans Golgi, yang mengarahkan
pengangkutan protein untuk lisosom.
Gambar 2.7 Penargetan protein lisosomal oleh fosforilasi residu mannose
Fosforilasi residu mannose merupakan langkah penting dalam memilah protein
lisosomal ke tujuan intraseluler mereka yang benar. Kekhasan dari proses ini berada
dalam enzim yang mengkatalisis langkah pertama dalam urutan reaksi-penambahan
selektif N-asetilglukosamin fosfat untuk protein lisosomal. Enzim ini mengenali
penentu struktural yang hadir pada protein lisosomal tetapi tidak pada protein ditujukan
untuk membran plasma atau sekresi. Penentu pengenalan ini bukanlah urutan sederhana
asam amino, melainkan terbentuk dalam protein terlipat oleh penjajaran urutan asam
amino dari berbagai daerah dari rantai polipeptida. Berbeda dengan urutan sinyal bahwa

33
protein translokasi langsung ke ER, penentu pengenalan yang mengarah ke fosforilasi
mannose, dan pada akhirnya menargetkan protein untuk lisosom, tergantung pada
konformasi tiga dimensi dari protein dilipat. Determinan tersebut patch called signal,
berbeda dengan sinyal menargetkan linear.
Protein juga dapat dimodifikasi dengan penambahan karbohidrat untuk rantai
samping serin akseptor dan residu treonin dalam urutan tertentu dari asam amino (O-
linked glikosilasi) (lihat Gambar 2.8). Modifikasi ini berlangsung di aparatus Golgi
dengan penambahan berurutan residu gula tunggal. Serin atau treonin biasanya
dihubungkan langsung ke N-asetilgalaktosamin, maka gula yang lain dapat
ditambahkan. Dalam beberapa kasus, gula ini lebih lanjut dimodifikasi dengan
penambahan kelompok sulfat.
Gambar 2.8. Modifikasi dengan penambahan karbohidrat
Glikosilasi terikat N berperan dalam mempromosikan protein terlipat dalam dua
cara. Pertama, berperan langsung dalam membuat lipatan intermediet lebih mudah larut,
sehingga mencegah agregasi protein. Kedua, modifikasi berurutan dari N-linked
oligosakarida membentuk "glyco-kode" yang menandai perkembangan protein folding
dan memediasi pengikatan protein untuk pendamping. Misalnya: Lektin, dalam
mengarahkan transportasi ER-ke-Golgi. Lektin juga berpartisipasi dalam memilah
protein dalam jaringan trans Golgi. Dengan cara ini, misalnya, kehadiran oligosakarida
cenderung membuat glikoprotein lebih tahan terhadap pencernaan oleh enzim
proteolitik. Ini mungkin bahwa oligosakarida pada sel-permukaan protein awalnya

34
disediakan sel leluhur dengan lapisan pelindung. Dibandingkan dengan dinding sel
bakteri kaku, mantel lendir memiliki keuntungan untuk meninggalkan sel dengan
kebebasan mengubah bentuk dan bergerak. Rantai gula sejak menjadi dimodifikasi
untuk melayani keperluan lain juga. Lapisan lendir dari paru-paru dan usus sel,
misalnya, melindungi terhadap banyak patogen. Pengakuan rantai gula oleh lektin
dalam ruang ekstraseluler adalah penting dalam banyak proses perkembangan dan
dalam sel-sel pegenalan: selectins, misalnya, lektin yang berfungsi dalam adhesi sel sel
selama migrasi limfosit. Kehadiran oligosakarida dapat mengubah sifat antigen protein,
membuat glikosilasi merupakan faktor penting dalam produksi protein untuk tujuan
farmasi.
Glikosilasi juga dapat memiliki peran regulasi yang penting. Signaling melalui
Notch reseptor permukaan sel-sinyal, misalnya, menentukan nasib sel dalam
pembangunan. Notch adalah protein transmembran yang 0-glikosilasi dengan
penambahan fucose tunggal untuk beberapa serines, threonines, dan hydroxylysines.
Beberapa jenis sel mengungkapkan transferase glycosyl tambahan yang menambahkan-
N-asetil glukosamin untuk masing-masing fucoses di aparatus GoIgi. Selain mengubah
spesifisitas reseptor Notch untuk sel-permukaan protein sinyal yang mengaktifkan
reseptor.
3. Metabolisme Lipid dan Polisakarida dalam aparatus Golgi
Selain kegiatan dalam pengolahan dan pemilahan glikoprotein, fungsi aparatus
Golgi khususnya dalam metabolisme lemak, dalam sintesis glikolipid dan
sphingomyelin. Seperti telah dibahas sebelumnya, fosfolipid gliserol, kolesterol, dan
ceramide disintesis di ER. Sphingomyelin dan glikolipid kemudian disintesis dari
ceramide dalam aparatus Golgi (Gambar 2.9). Sphingomyelin (satu fosfolipid
nonglycerol hanya dalam membran sel) disintesis oleh transfer gugus
phosphorylcholine dari fosfatidilkolin ke ceramide. Atau, penambahan karbohidrat
untuk ceramide dapat menghasilkan berbagai glikolipid yang berbeda.

35
Gambar 2.9 Synthesis dari sphingomyelin dan glikolipid. Ceramide, yang disintesis di UGD, diubah baik untuk sphingomyelin (fosfolipid) atau glikolipid dalam aparatus Golgi. Dalam reaksi pertama, kelompok phosphorylcholine ditransfer dari fosfatidilkolin ke ceramide. Atau, berbagai glikolipid yang berbeda dapat disintesis dengan penambahan satu atau lebih residu gula (misalnya, glukosa).
Sphingomyelin disintesis pada permukaan lumenal dari Golgi, tetapi glukosa
ditambahkan ke ceramide di sisi sitosol ternyata kemudian Glucosylceramide membalik
dan karbohidrat tambahan ditambahkan di sisi lumenal membran. Baik sphingomyelin
maupun glikolipid kemudian dapat mentranslokasi melintasi membran Golgi, sehingga
hanya ditemukan pada paruh lumenal dari bilayer Golgi. Setelah transportasi vesikuler,
diterjemahkan ke setengah luar membran plasma, dengan gugus kepala polar mereka
terkena pada permukaan sel. Oligosakarida dari glikolipid merupakan penanda
permukaan penting dalam sel-sel pengenal. Dalam sel tumbuhan, aparatus Golgi
memiliki tugas tambahan sebagai tempat sintesis polisakarida yang kompleks dari
dinding sel.
Dinding sel tanaman terdiri dari tiga jenis utama dari polisakarida. Konstituen
dominan Selulosa adalah polimer linear sederhana residu glukosa. Hal ini disintesis
pada permukaan sel oleh enzim dalam membran plasma. Polisakarida dinding sel lain
(hemiselulosa dan pektin) adalah kompleks molekul rantai bercabang yang disintesis
dalam aparatus Golgi dan kemudian diangkut dalam vesikel ke permukaan sel. Sintesis
dari polisakarida dinding sel adalah fungsi selular utama, dan sebanyak 80% dari
aktivitas metabolisme dari aparatus Golgi dalam sel tanaman dapat dikhususkan untuk
sintesis polisakarida.

36
4. Sortasi dan Ekspor Protein dari Aparatus Golgi
Protein, serta lipid dan polisakarida, yang diangkut dari aparatus Golgi ke tujuan
akhir mereka melalui jalur sekresi. Ini melibatkan penyortiran protein menjadi berbagai
jenis vesikel transportasi, tunas dari jaringan trans Golgi memberikan isinya ke lokasi
selular yang sesuai (Gambar 2.10). Beberapa protein yang dibawa dari Golgi ke
membran plasma oleh jalur sekresi konstitutif, yang menyumbang penggabungan
protein baru dan lipid ke dalam membran plasma, serta untuk sekresi terus menerus
protein dari sel. Protein lain diangkut ke permukaan sel dengan jalur yang berbeda dari
sekresi diatur atau secara khusus ditargetkan untuk tujuan intraseluler lainnya, seperti
lisosom dalam sel hewan atau vakuola dalam ragi.
Gambar2.10Transport dari aparatus Golgi. Protein yang diurutkan dalam jaringan trans Golgi diangkut dalam vesikel ke tujuan akhir mereka. Dengan tidak adanya sinyal menargetkan khusus, protein dibawa ke membran plasma oleh sekresi konstitutif. Atau, protein dapat dialihkan dari jalur sekresi konstitutif dan ditargetkan untuk tujuan lain, seperti lisosom atau sekresi diatur dari sel.
5. Transport Melalui Badan Golgi Dapat Terjadi oleh Transport vesikular atau Pematangan Cisternal
Bahan-bahan bergerak melalui berbagai kompartemen dari kompleks Golgi yang
telah lama dibentuk, namun, dua pandangan bertentangan tentang cara ini telah
mendominasi selama bertahun-tahun. Sampai pertengahan 1980-an, secara umum
diterima bahwa Golgi cisternae adalah struktur sementara. Hal ini menunjukkan bahwa
Golgi cisternae terbentuk di daerah cis dari tumpukan, oleh fusi dari pengangkut
membran membran dari ER dan ERGIC dan bahwa setiap cisterna secara fisik pindah
dari cis sampai trans yang merupakan akhir tumpukan, perubahan dalam komposisi

37
mengalami perkembangan. Hal ini dikenal sebagai model pematangan cisternal karena,
menurut model, masing-masing cisterna "matang" ke dalamcisterna selanjutnya di
sepanjang tumpukan.
Dari pertengahan 1980-an sampai pertengahan 1990-an, model pematangan dari
pergerakan Golgi sebagian besar ditinggalkan dan diganti dengan model lain, yaitu
yang berpendapat bahwa cisternae dari tumpukan Golgi tetap di tempat sebagai
kompartemen stabil. Dalam model yang terakhir ini, yang dikenal sebagai model
transportasi vesikuler, muatannya (seperti, sekretorik, lisosomal, dan protein membran)
bergerak melalui tumpukan Golgi, dari CGN ke TGN, dalam vesikel yang tunasnya dari
satu kompartemen membran dan bergabung dengan kompartemen tetangga sepanjang
tumpukan. Model transportasi vesikuler diilustrasikan dalam Gambar 2.11, dan
penerimaan ini sebagian besar didasarkan pada pengamatan berikut:
1. Setiap cisternae Golgi dari berbagai tumpukan memiliki jumlah enzim yang berbeda
(Gambar 2.11A). Bagaimana bisa berbagai cisternae memiliki sifat yang berbeda
jika setiap cisterna mengalami peningkatan pada jalan ke yang berikutnya, seperti
disarankan oleh model pematangan cisternal.
2. Sejumlah besar vesikel dapat dilihat pada mikrograf elektron dari awal sampai
pinggiran Golgi cisternae. Pada tahun 1983, James Rothman dan rekan-rekannya di
Stanford University menunjukkan, dengan menggunakan sel bebas dari membran
Golgi bahwa transportasi vesikel telah bisa melewati satu cisterna Golgi dan
bergabung dengan Golgi cisterna in vitro. Percobaan ini terbentuk dari hipotesis
dasar yang menunjukkan bahwa dalam sel, muatan-membentangkan ujung vesikula
dari cis cisternae dan menyatu dengan cisternae yang terletak di posisi yang lebih
trans dalam tumpukan.

38
Gambar 11. Model transport dalam aparatus golgi (A) Vesikular Transport (B)Cisternal Maturation Model
Meskipun kedua model fungsi Golgi terus memiliki pendukung mereka,
konsensus pendapat telah bergeser kembali ke model pematangan cisternal. Beberapa
alasan utama untuk pergeseran ini dapat dicatat:
1. Model pematangan cisternal memperlihatkan kedinamisan yang tinggi dari
kompleks Golgi di mana unsur utama dari organel ini, cisternae, terus-menerus
terbentuk di permukaan cis dan tersebar ke permukaan trans. Menurut pandangan
ini, keberadaan kompleks Golgi sendiri sangat tergantung pada terus-menerus
masuknya operator transportasi dari ER dan ERGIC. Seperti yang diperkirakan oleh
pematangan cisternal. Model, ketika pembentukan operator transportasi dari ER
dihambat oleh perlakuan sel yang dipengaruhi obat-obatan spesifik atau penggunaan
suhu-sensitif mutan, kompleks Golgi menghilang.ketika obat tersebut dihilangkan
atau sel mutan dikembalikan ke temperatur permisif, kompleks Golgi dengan cepat
berkumpul sebagai ER dan transportasi Golgi diperbarui.
2. Beberapa bahan yang diproduksi dalam reticulum endoplasma dan berjalan melalui
kompleks Golgi terbukti tetap dalam cisternae Golgi dan tidak pernah terlihat Golgi
terkait vesikel transportasi. Misalnya, studi tentang fibroblast menunjukkan bahwa
kompleks besar molekul prokolagen (prekursor dari ekstraseluler kolagen) pindah
dari cisternae cis ke trans cisternae tanpa pernah meninggalkan lumen cisternal.
3. Diasumsikan sampai pertengahan 1990-an bahwa vesikel transport selalu bergerak
ke arah "depan" (anterograde), yaitu, dari daerah asal cis ke daerah trans. Tapi
sebagian besar bukti telah menunjukkan bahwa vesikula dapat bergerak ke arah
"mundur" retrograde), yaitu dari trans donor membran ke membran akseptor cis.

39
4. Studi pertumbuhan sel ragi mengandung fluorescently berlabel Golgi protein telah
menunjukkan secara langsung bahwa komposisi dari cisterna Golgi individu dapat
berubah waktu dari satu yang berisi (cis) protein Golgi menjadi salah satu yang
berisi (trans) protein Golgi. Hasil dari penelitian ini ditampilkan pembukaan
mikrograf pada Bab 18 dan mereka tidak kompatibel dengan model vesikel
transport. Apakah iya atau tidak, hasil pada ragi ini dapat diekstrapolasikan ke
mamalia Golgi kompleks, yang lebih kompleks, struktur yang bertumpukan, masih
harus dijelaskan.
H. KERUSAKAN APARATUS GOLGI DAN AKIBATNYA
Apa yang terjadi ketika ada kerusakan fungsi aparatus Golgi? Cacat dalam
berbagai aspek fungsi Golgi dapat mengakibatkan gangguan glikosilasi kongenital,
beberapa bentuk distrofi otot, dan dapat menyebabkan diabetes, kanker, dan cystic
fibrosis (Ungar 2009).
1. Kanker
Salah satu fungsi badan golgi sebagai tempat sekresi asam amino untuk
membentuk hormon. Salah satu hormon yang berperan dalam perkembangan sel
kanker adalah hormon estrogen. Hormon estrogen ini berfungsi dalam merangsang
pertumbuhan sel tidak terkecuali juga untuk sel kanker. Sehingga dapat berpotensi
meningkatkan perkembangan sel kanker tersebut. Salah satu hormon yang dapat
menghambat perkembangan sel adalah hormon progestron yang dapat melindungi
perkembangan sel yang berlebihan. Hal ini akan menjadi salah satu masalah
apabila badan golgi mensekresi terlalu banyak hormon estrogen serta terlalu sedikit
mensekresikan hormon progestron. Sehingga terjadinya gangguan keseimbangan
pada hormone tersebut. Akibatnya hormon estrogen akan terus merangsang
perkembangan sel kanker tanpa dihalangi oleh hormon progestron.
2. Dwarfism
Hormon pertumbuhan barupa polipeptida dengan bm 22.000. Secara fisioligis,
sekresinya diatur oleh hipothalamus. Hipothalamus menghasilkan faktor
pengelepas hormon pertumbuhan (GHRF – growth hormone releasing factor) dan
juga menghasilkan somatostatin (GHIH – growth hormone inhibitory hormone)
yang menghambat sekresi hormon pertumbuhan. Defisiensi hormon pertumbuhan
dapat disebabkan oleh defek hipofisis (tidak adanya hormon pertumbuhan) atau

40
sekunder dari disfungsi hipotalamus (tidak adanya GHRH). Hiposekresi hormon
pertumbuhan pada anak-anak menimbulkan cebol (dwarfism). Hiposekresi hormon
pertumbuhan disebabkan oleh menurunnya sintesis protein oleh sel, sehingga
pertumbuhan tulang terhambat. Menurunnya sintesis protein oleh sel dapat
disebabkan oleh gangguan pada badan golgi dalam mengemas hasil produksi
ribosom.
3. Cystic fibrosis
Cystic fibrosis atau CF, adalah penyakit kelenjar sekretori warisan , termasuk
kelenjar yang memproduksi berlebihan lendir dan keringat. CF kebanyakan
mempengaruhi paru-paru, pankreas, hati, usus, sinus, dan organ-organ seks.
Lendir merupakan zat yang dibuat oleh lapisan dari beberapa jaringan tubuh.
Biasanya, lendir adalah zat, licin berair. Itu membuat lapisan-lapisan dari organ-
organ tertentu lembab dan mencegah mereka dari pengeringan atau mendapatkan
terinfeksi. Hal ini tentu saja akan mempengaruhi kerja badan golgi pada sel-sel
organ tersebut.
Misalnya pada organ paru-paru,dimana lendir menumpuk di paru-paru dan saluran
udara yang membawa udara masuk dan keluar dari paru-paru. Penumpukan lendir
memudahkan bakteri untuk tumbuh. Badan golgi sendiri akan terhalangi fungsinya
oleh lendir tersebut. Sehingga memungkinkan terjadinya infeksi paru-paru serius.
4. Tay-Sachs
Lisosom; organ pencernaan sel. Tersusun dari membran yang mengandung enzim-
enzim hidrolitik kuat. Enzim-enzim tersebut berasal dari kompleks golgi. Lisosom
baru terbentuk dari kumpulan khusus enzim hidrolitik yang baru disintesis dan
tertangkap di dalam vesikel berselubung yang kemudian melepaskan diri dari
kompleks golgi. Suatu bahaya inheren bahkan pada sel sehat dan utuh adalah
pecahnya membran lisosom tanpa sengaja dan menimbun di dalam lisosom.
Faktornya bisa disebabkan oleh enzim-enzim hidrolitik yang bekerja optimal pada
suasana asam, sehingga bagian dalam lisosom lebih asam dari bagian luarnya.
Salah satu penyakit ini adalah penyakit Tay-Sachs, ditandai oleh adanya
penimbunan abnormal senyawa golongan gangliosida, yaitu molekul kompleks
yang ditemukan di sel-sel saraf.

41
DAFTAR PUSTAKA
Alberts, B. et al. 2008. Molecular Biology Of The Cell Ed. 5th. USA: Garland Science.
Anonim. 2013. Rough Endoplasmic Reticulum Function. (Online), Buzzle, (http://www.buzzle.com/articles/rough-endoplasmic-reticulum-function.html), Diakses 11 mei 2013.
Anonim. 2013. Endoplasmic Reticulum, Golgi Apparatus, and Lysosomes. (Online) (http://www.nature.com), Diakses tanggal 9 Maret 2013.
Anonim. 2013. Kajian Sistem Endomembran, Retikulum Endoplasma, Vesikula, Badan Golgi Dan Lisosom. (Online), Makalah. 2012. (www.scribb.com). Diakses tanggal 9 Maret 2013.
Anonim. 2013. Struktur dan Fungsi Retikulum Endoplasma (RE). (Online), (http://www.bimbie.com/struktur-retikulum-endoplasma.htm), diakses 9 maret 2013.
Anonim. 2013. The Golgi Aparatus . (Online) (http://micro.magnet.fsu.edu). Diakses
tanggal 9 Maret 2013.
Alim, T. 2013. Retikulum Endoplasma (RE).(Online), ( www.biologi-sel.com/2013/.../ retikulum - endoplasma -re.htm ), diakses 9 Maret 2013.
Cooper, G.M. 2000. The Cell: A Molecular Approach, 2nd Ed. (Online), NCBI Bookshelf. diakses 11 Mei 2013.
Zhao, L. dan Ackerman, S. L. 2006. Endoplasmic Reticulum Stress In Health And Disease. (Online), Jurnal Current Opinion in Cell Biology 2006, 18:444–452, (http://www8.umoncton.ca/umcm-gauthier_didier/bc6423/4SRE/Zhao06%20-%20ER%20stress.pdf), diakses 9 Maret 2013.




















