LI Stevanus
28
Karbohidrat Karbohidrat tersebar luas pada tumbuhan dan hewan, mereka memiliki peran struktural dan metabolik penting. Di tanaman, glukosa disintesis dari karbon dioksida dan air oleh fotosintesis dan disimpan sebagai pati atau digunakan untuk mensintesis selulosa dari dinding sel tanaman. Hewan dapat mensintesis karbohidrat dari asam amino, tetapi kebanyakan berasal akhirnya dari tanaman. Glukosa adalah karbohidrat paling penting, karbohidrat yang paling diserap ke dalam aliran darah sebagai glukosa dibentuk oleh hidrolisis pati makanan dan disakarida, dan lainnya gula diubah menjadi glukosa di hati. Glukosa adalah bahan bakar metabolik utama mamalia (kecuali ruminansia) dan bahan bakar universal janin. Ini adalah prekursor untuk sintesis semua karbohidrat lain dalam tubuh, termasuk glikogen untuk disimpan, ribosa dan deoksiribosa asam nukleat, galaktosa dalam laktosa susu, dalam glikolipid, dan dalam kombinasi dengan protein dalam glikoprotein dan proteoglikan. Penyakit yang berhubungan dengan metabolism karbohidrat termasuk diabetes mellitus, galaktosemia, penyakit penyimpanan glikogen, dan laktosa intoleransi. Karbohidrat adalah aldehid atau keton derivate dari Alkohol polihidrat Karbohidrat diklasifikasikan sebagai berikut: 1. Monosakarida adalah mereka gula yang tidak dapat dihidrolisis menjadi karbohidrat sederhana. Mereka adalah aldehid atau keton dengan formula (CH2O)x, dimana x lebih dari
-
Upload
stevanus-handrawan -
Category
Documents
-
view
67 -
download
6
Transcript of LI Stevanus

Karbohidrat
Karbohidrat tersebar luas pada tumbuhan dan hewan, mereka memiliki peran
struktural dan metabolik penting. Di tanaman, glukosa disintesis dari karbon dioksida dan air
oleh fotosintesis dan disimpan sebagai pati atau digunakan untuk mensintesis selulosa dari
dinding sel tanaman. Hewan dapat mensintesis karbohidrat dari asam amino, tetapi
kebanyakan berasal akhirnya dari tanaman. Glukosa adalah karbohidrat paling penting,
karbohidrat yang paling diserap ke dalam aliran darah sebagai glukosa dibentuk oleh
hidrolisis pati makanan dan disakarida, dan lainnya gula diubah menjadi glukosa di hati.
Glukosa adalah bahan bakar metabolik utama mamalia (kecuali ruminansia) dan bahan bakar
universal janin. Ini adalah prekursor untuk sintesis semua karbohidrat lain dalam tubuh,
termasuk glikogen untuk disimpan, ribosa dan deoksiribosa asam nukleat, galaktosa dalam
laktosa susu, dalam glikolipid, dan dalam kombinasi dengan protein dalam glikoprotein dan
proteoglikan. Penyakit yang berhubungan dengan metabolism karbohidrat termasuk diabetes
mellitus, galaktosemia, penyakit penyimpanan glikogen, dan laktosa intoleransi.
Karbohidrat adalah aldehid atau keton derivate dari Alkohol polihidrat
Karbohidrat diklasifikasikan sebagai berikut:
1. Monosakarida adalah mereka gula yang tidak dapat dihidrolisis menjadi karbohidrat
sederhana. Mereka adalah aldehid atau keton dengan formula (CH2O)x, dimana x lebih dari
3. Mereka mungkin diklasifikasikan sebagai triosa, tetroses, pentosa, heksosa, atau heptoses,
tergantung pada jumlah karbon atom, dan sebagai aldoses atau ketoses, tergantung pada
apakah mereka memiliki aldehida atau kelompok keton. Selain aldehid dan keton, alkohol
polihidrat (gula alkohol atau poliol), dimana aldehid atau keton kelompok telah berkurang ke
grup alkohol, juga terjadi secara alami dalam makanan. Mereka disintesis oleh pengurangan
monosakarida untuk digunakan dalam pembuatan makanan untuk penurunan berat badan dan
untuk penderita diabetes. Mereka terserap sedikit, dan memiliki sekitar setengah hasil energy
gula.
2. Disakarida adalah dihidrolisis oleh enzyme pencernaan, dan menghasilkan monosakarida
yang diabsorbsi ke dalam tubuh. Contohnya :
Maltose = glucose + glucose
Lactose = glucose + galactose

Sucrose = glucose + fructose
3.Oligosakarida adalah produk kondensasi tiga sampai sepuluh monosakarida. Kebanyakan
tidak dicerna oleh enzim manusia.
4. Polisakarida adalah produk kondensasi lebih dari sepuluh unit monosakarida, contoh
adalah pati dan dekstrin, yang mungkin polimer lurus atau bercabang. Polisakarida kadang-
kadang diklasifikasikan sebagai hexosans atau pentosan, tergantung pada identitas
monosakarida konstituen (heksosa dan pentosa, masing-masing). Selain pati dan dekstrin,
makanan mengandung berbagai macam lainnya polisakarida yang secara kolektif dikenal
sebagai polisakarida nonstarch, mereka tidak dapat dicerna oleh manusia enzim, dan
merupakan komponen utama dari serat makanan. Contohnya adalah selulosa dari dinding sel
tumbuhan (polimer glukosa) dan inulin, karbohidrat penyimpanan di beberapa tanaman
(polimer fruktosa).
Glycolisis
1. Glycolysis terdiri dari 5 reaksi yang menggunakan ATP dan 5 reaksi yang
memproduksi ATP
2. Pyruvate dehydrogenase mengkatalisis 3 reaksi dengan multienzyme kompleks yang
sama.
3. Glycolysis diregulasi dengan 3 point : hexokinase, phosphofructokinase (PFK), dan
pyruvate kinase.
4. Pyruvate dehydrogenase diatur dengan modifikasi kovalen dengan fosforilasi
5. Interfase glikolisis dengan metabolisme glikogen, jalur pentosa fosfat, pembentukan
gula amino, sintesis trigliserida (dengan cara gliserol 3-fosfat), produksi laktat (reaksi
buntu), dan transaminasi dengan alanin.
6. Interfase Pyruvate dehydrogenase dengan jalur lain seperti siklus asam sitrat atau
sintesis lemak melalui produk, asetil KoA.
7. Asidosis laktat disebabkan oleh overproduction dari pyruvate atau NADH.

Glycolysis and pyruvate oxidation: pathway reaction steps
1. Step 1
a. Fosforilasi dari glucose menjadi glucose 6-phophate, merupakan regulasi
pertama dalam glikolisis, bersifat irreversible dan memperangkap glukosa di
dalam sel Fosforilasi : Memperangkap glukosa di dalam sel
b. Menggunakan 1 ATP
c. 2 enzym yang mengkatalisis langkah ini adalah hexokinase dan glucokinase,
yang khusus berfungsi dalam keadaan yang berbeda.
d. Hexokinase, ada di semua jaringan, aktif saat konsentrasi glukosa rendah (low
Km) dan tidak dapat memfosforilasi dengan cepat glukosa yang berjumlah
besar (low Vmax).
Hexokinase: Fosforilasi glucose; low Km, low Vmax
Hexokinase: low Km memastikan penyerapan glukosa saat puasa
e. Glucokinase, ada di liver dan sel β prankeas, sangat aktif hanya saat kadar
glukosa yang tinggi (high Km) dan memfosforilasi dengan cepat glukosa yang
berjumlah besar (high Vmax).

Glucokinase: Fosforilasi glucose; high Km, high Vmax
2. Step 2
a. Reaksi reversibel yang melibatkan konversi glukosa 6-fosfat menjadi fruktosa
6-fosfat dengan phosphoglucose isomerase
3. Step 3
a. Reaksi ireversibel melibatkan konversi fruktosa 6-fosfat menjadi fruktosa 1,6-
bifosfat oleh phoisphofruktokinase 1 (PFK-1), yang merupakan tingkat
pembatas enzim glikolisis
b. Menggunakan 1 ATP
4. Step 4
a. Konversi reversible dari fruktosa 1,6-biphophate menjadi dua 3-carbon
intermediates oleh aldolase A
b. Triose phosphate isomerase mengkonversi secara reversible glyceraldehyde 3-
phosphate menjadi dihydroxyacetone phosphate (DHAP).
c. DHAP dikonversi secara reversible menjadi glycerol 3-phosphate oleh
glycerol 3-phosphate dehydrogenase, dimana digunakan NADH sebagai
kofaktor
DHAP dikonversi menjadi glycerol 3-phosphate: triacylglycerol synthesis,
gluconeogenesis, ETC shuttle
(1) Glycerol 3-phosphate adalah substrat primer untuk synthesis dari
triacylglycerol di liver dalam keadaan makan
(2) Glycerol 3-phosphate digunakan untuk mengangkut NADH
menuju ke electron transport chain (ETC) di dalam membrane
mitochondria
(3) Dalam keadaan puasa, glycerol 3-phosphate dikonversi menjadi
DHAP, yang digunakan sebagai substrat untuk gluconeogenesis.
Glycerol 3-phosphate keadaan puasa: dikonversi menjadi DHAP dan
digunkana sebagai substrat untuk gluconeogenesis
5. Step 5

a. Konversi reversible dari glyceraldehyde 3-phosphate menjadi 1,3-
bisphosphoglycerate (1,3-BPG) oleh glyceraldehyde 3-phosphate
dehydrogenase menggunakan NAD+ sebagai kofaktor.
b. NAD+ harus diisi ulang untuk melanjutkan glikolisis.
c. NADH diangkut menuju ETC oleh glycerol phosphate shuttle atau malate-
aspartate shuttle
d. Methemoglobin reductase system dan jalur untuk sintesis 2,3-
bisphosphoglycerate (2,3-BPG) adalah jalur tambahan yang berasal dari reaksi
ini
6. Step 6
a. Konversi reversible dari 1,3-BPG menjadi 3-phosphoglycerate oleh
phosphoglycerate kinase
b. Reaksi ini menghasilkan 2 ATP per molekul glukosa, dimana digunakan 1
ATP untuk step 1 dan 1 ATP untuk step 3.
7. Step 7
a. Konversi reversible dari 3-phosphoglycerate menjadi 2-phosphoglycerate oleh
phosphoglycerate mutase
8. Step 8
a. Konversi reversible dari 2-phosphoglycerate menjadi phosphoenolpyruvate
(PEP) oleh enolase
9. Step 9
a. Diregulasi, reaksi irreversible yang menyangkut konversi dari PEP menjadi
pyruvate oleh pyruvate kinase
b. Ada pendapatan bersih dari 2 ATP per molekul glukosa di reaksi ini
PEP konversi menjadi pyruvate: keuntungan bersih 2 ATP
10. Step 10
a. Konversi reversible dari pyruvate menjadi laktat oleh lactate dehydrogenase
menggunakan NADH sebagai cofactor
Pyruvate menjadi lactate: anaerobic glycolysis; alcoholics
b. Reaksi ini terjadi di glikolisis anaerobic berhubungan dengan shock dan
latihan extrim

c. This reaction also occurs in alcoholics because of an increase in NADH from
alcohol metabolism (see Chapter 9).
Lactic acidosis: Biasanya alkoholik
11. Pyruvate dehydrogenase, kompleks multienzyme yang besar terletak di matrix
mitochondria, berperan pada piruvat untuk memproduksi acetyl CoA dan 2 NADH.
a. 5 Koenzyme dibutuhkan untuk reaksi di atas.
(1) Koenzym yang berasal dari vitamins termasuk thiamine
pyrophosphate, FAD, NAD+, dan coenzyme A.
(2) Lipoic acid adalah coenzyme diproduksi dari octanoic acid.
b. α-Ketoglutarate dehydrogenase, yang melakukan reaksi analog untuk
membentuk succinyl CoA dalam siklus asam sitrat, juga kompleks
multienzyme dan membutuhkan koenzyme yang sama
12. Maksimal ATP hasil dari oksidasi glukosa adalah 36 hingga 38 ATP
Glikolisis anaerobik: 2 ATP per molekul glukosa
Glikolisis aerobik: 36 hingga 38 ATP per molekul glukosa
Perbandingan dari Hexokinase dan Glucokinase
ARAKTERISTIK Heksokinase Glukokinase
Lokasi jaringan Semua Hati dan pankreas sel β
K m Rendah (afinitas substrat tinggi) Tinggi (afinitas substrat rendah)
V max Rendah Tinggi
Dihambat oleh
glukosa 6-fosfat
Ya Tidak
Diinduksi oleh insulin Tidak Ya
Spesifisitas substrat Glukosa, fruktosa, galaktosa Hanya glukosa
Peran fisiologis Menyediakan sel dengan tingkat
basal glukosa 6-fosfat dibutuhkan
untuk produksi energi
Izin akumulasi glukosa
intraseluler untuk konversi
menjadi glikogen atau
trigliserida

Karakter Unik
1. Perbandingan glikolisis aerobic dan anaerobic
a. NADH yang dihasilkan dalam jalur glikolisis harus dioksidasi untuk
meregenerasi NAD + untuk melanjutkan glikolisis
b. Dalam glikolisis aerobik, elektron dari NADH ditransfer ke ETC oleh glycerol
phosphate shuttle atau malate-aspartate shuttle, dan + NAD dikembalikan ke
sitosol
(1) Ada keuntungan bersih 2 ATP dan 2 NADH per molekul glukosa
dalam glikolisis aerobic
(2) 2 NADH menghasilkan total 4 ATP (glycerol phosphate shuttle)
atau 6 ATP (malate-aspartate shuttle) per molekul glukosa
Aerobic glycolysis: keuntungan bersih 2 ATP dan 2 NADH molekul
Anaerobic glycolysis: Keuntungan bersih 2 ATP molecules;
NAD+ diisi ulang
2. Dalam glikolisis anaerobic, reduksi dari pyruvate menjadi lactate oleh lactate
dehydrogenase meregenerasi NAD+
a. Keuntungan bersih 2 ATP dan tidak ada NADH per molekul glukosa dalam
glikolisis anaerobic.
b. Laktat dikonversi kembali menjadi piruvat dalam hati atau diekskresi melalui
urine.
c. RBCs matang sedikit mitochondria dan bergantung pada metabolism
anaerobic untuk sintesis ATP
Clinical Relevance
1. Asidosis Laktat (meningkatnya laktat dalam darah)
a. Dalam glikolisis aerobik, elektron dari NADH ditransfer ke ETC oleh glycerol
phosphate shuttle atau malate-aspartate shuttle, dan + NAD dikembalikan ke
sitosol
Pyruvate dehydrogenase deficiency: lactic acidosis, penurunan acetyl CoA,
kematian dini.

b. Dalam glikolisis anaerobic, reduksi piruvat menjadi laktat oleh lactate
dehydrogenase meregenerasi NAD+ (see Fig. 6-1).
(1) Hypoxia menyebabkan intramitochondrial NADH membalikan
malate shuttle , mengakibatkan meningkatnya cytosolic NADH
(2) Kondisi lain seperti defisiensi pyruvate dehydrogenase
(meningkatkan piruvat) dan metabolism etanol (meningkatkan NADH)
mengakibatkan laktat berlebih.
c. Laktat di konversi balik menjadi piruvat dalam hati atau diekskresi oleh urine.
(1) RBCs matang sedikit mitochondria dan bergantung pada
metabolism anaerobic untuk sintesis ATP
2. Defek Glycolytic dan pyruvate dehydrogenase enzyme (contohnya, defisiensi
pyruvate kinase, defisiensi pyruvate dehydrogenase)
DEKARBOKSILASI OKSIDATIF / REAKSI TRANSISI
Adalah proses konversi Asam piruvat menjadi gugus karboksil dengan atom C &
bergabung dengan CoA membentuk Asetil Coenzim ( CoA ) yang terjadi di mitokondria.
CoA adalah koenzim A yang dibentuk dari vitamin B asam pentatonat.
Asetil CoA adalah ikatan yang terdiri atas 2 ikatan C( asetat ) yang terkait pada 1
molekul CoA
Hasil :
2 NADH = 6 ATP / molekul glukosa
SIKLUS KREB
Ditemukan oleh Sir Hans Kreb
Disebut juga :
- Siklus ASAM SITRAT karena setelah asetil KoA memasuki siklus, ikatan
pertama yang terbentuk adalah asam sitrat.
- Siklus TCA ( Tri Carboxylic Acid ) karena melibatkan asam sitrat dengan 3

gugus karboksil ( COOH ).
Terjadi di mitokondria
Hasil :
- 2 CO2
- 6 NADH = 18 ATP
- 2 FADH2 = 4 ATP
- 2 ATP = 2 ATP
Total = 24 ATP
SISTEM TRANSPOR ELEKTRON / FOSFORILASI OKSIDATIF
Adalah reaksi yang digunakan O2 untuk mengubah moleku-molekul NADH & FADH2
menjadi NAD & FAD, air & ATP yang terjadi pada membran mitokondria.
Reaksi ini membutuhkan unsur Fe & Cu sebagai katalisator.Elektron berenergi tinggi
dalam NADH dan FADH2 dilewatkan setahap demi setahap ke tingkat energi yang rendah
dan akhirnya diterima oksigen (O2) .
Di sistem transpor elektron, oksidasi :
- 1 NADH = 3 ATP
- ! FADH2 = 2 ATP
Gluconeogenesis
Overview
1. 4 enzim kunci dalam jalur gluconeogenesis memotong jalur irreversible glikolisis
a. Pyruvate carboxylase, phosphoenolpyruvate (PEP) carboxykinase, fructose
1,6-bisphosphatase, glucose 6-phosphatase.
2. Regulasi timbal-balik dengan rasio insulin - glucagon memastikan bahwa glikolisis
dan glukoneogenesis tidak terjadi bersamaan
3. Gluconeogenesis merupakan jalur yang membutuhkan energi.
a. Energi disuplai dari by oksidasi asam lemak
4. Gluconeogenesis jalur yang membutuhkan kerangka karbon

a. Kerangka karbon disuplai oleh asam amino dari otot rangka dan laktat dari
otot dan RBCs
5. Hanya hati yang dapat menggunakan gliserol bebas dalam mobilisasi asam lemak
untuk glukoneogenesis.
6. Kekurangan enzim gluconeogenic dalam berpuasa mengakibatkan hypoglycemia,
seperti kekurangan glukosa 6-phosphatase pada von Gierke’s disease
Jalur Gluconeogenesis
1. Enzym yang dibutuhkan :
a. Pyruvate carboxylase
b. Phosphoenolpyruvate (PEP) carboxykinase
c. Fructose 1,6-bisphosphatase (rate-limiting reaction)
d. Glucose 6-phosphatase
2. Konversi piruvat menjadi PEP, dimana memotong reaksi irreversible piruvat kinase,
terjadi dalam beberapa langkah :
a. Pyruvate carboxylase, biotin yang mengandung enzyme mitochondria,
mengkonversi piruvat menjadi (OAA) dalam sebuah reaksi irreversible yang
mengkonsumsi ATP
Pyruvate kinase bypass: pyruvate carboxylase + PEP carboxykinase
(1) Biotin deficiency membentuk piruvat, dimana dikonversi menjadi
asam laktat dan membawa ke asidosis laktat
b. OAA direduksi menjadi malate (misalnya, malate shuttle), dimana ditransport
menuju cytosol dan kemudian di-reoksidasi menjadi OAA
OAA tidak dapat keluar dari mitochondria dan harus dikonversi menjadi
malate
c. PEP carboxykinase decarboxylates OAA untuk memproduksi PEP dalam
reaksi reversible yang mengkonsumsi GTP
3. Konversi PEP menjadi fruktosa 1,6-bifosfat terjadi denganConversion of PEP to

fructose 1,6-bisphosphate occurs by pembalikan sederhana enam reaksi dalam jalur
glikolisis
4. Fructose 1,6-bisphosphatase (rate-limiting enzyme), mem-defosforilasi fructose 1,6-
bisphosphate untuk memproduksi fructose 6-phosphate (step 3), memotong reaksi
irreversible PFK-1
5. Pembalikan sederhana dari reaksi fosfoglucose isomerasi mengkonversi fruktosa 6-
fosfat menjadi glukosa 6-fosfat
6. Glucose 6-phosphatase mem-defosforilasi glucose 6-phosphate untuk memproduksi
glucose (step 4), memotong reaksi irreversible hexokinase atau glucokinase
Rasio insulin dan glucagon meregulasi piruvat kinase ( Konversi PEP menjadi piruvat) dan
fructose 2,6-bisphosphate, dimana mengaktivkan PFK-1.

a. Insulin tinggi dan glucagon rendah (fase makan)
(1) Meningkatkan aktivitas piruvat kinase dan meningkatkan
levelfructose 2,6-bisphosphate
(2) Hasil: meningkatkan glikolisis (biasanya di hati) dan mengurangi
gluconeogenesis
b. Insulin rendah dan glucagon tinggi (fase puasa)
(1) Menurunkan aktivitas pyruvate kinase dan menurunkan level
fructose 2,6-bisphosphate levels
(2) Hasil: mengurangi glycolysis (biasanya di hati) dan meningkatkan
gluconeogenesis (mengatur gula darah)
Perbandingan Glikolisis dan Gluconeogenesis
Glycolysis Gluconeogenesis
Hexokinase (or glucokinase) Glucose 6-phosphatase
Glucose → glucose 6-phosphate Glucose 6-phosphate → glucose (
(-) Glucose 6-phosphate (hexokinase)
Phosphofructokinase 1 Fructose 1,6-bisphosphatase
Fructose 6-phosphate → fructose 1,6-
bisphosphate (step 3)
Fructose 1,6-bisphosphate → fructose
6-phosphate (step 3)
(+) AMP, fructose 2,6-bisphosphate (-) AMP, fructose 2,6-bisphosphate
(-) ATP, citrate (+) Citrate
Rate-limiting reaction Rate-limiting reaction
Pyruvate kinase Pyruvate carboxylase (in mitochondria)
PEP → pyruvate Pyruvate → oxaloacetate (OAA)
(+) Acetyl CoA
Requires biotin; OAA is converted to
malate in the mitochondria, and malate
is converted back to OAA in the
cytosol
PEP carboxykinase (in cytosol)
OAA PEP (step 2)
Only reversible reaction in

gluconeogenesis
Karakter Unik
1. Secara total 6 ATP dikonsumsi dalam glukoneogenesis
2. Hati tempat terpenting untuk gluconeogenesis, ginjal, epitel intestinum tenue kurang
berperan (terutama dalam keadaan lapar)
a. Dalam lapar yang sangat lama, ginjal memegang peran penting
gluconeogenesis .
3. Gluconeogenesis penting untuk menjaga kadar gula darah dalam keadaan puasa untuk
keperluan energy otak, sel darah merah, olahraga
4. Gluconeogenesis terjadi di dalam bagian mitochondria (reaksi pyruvate carboxylase)
dan sebagian di sitosol

Clinical relevance
1. Kekurangan enzyme gluconeogenesis mengakibatkan hypoglycemia
2. Contoh: Von Gierke's disease, a glycogen storage disease, Tidak adanya glucose 6-
phosphatase mengakibatkan kurangnya sintesis glukosa
Lipid
Lipid adalah kelompok heterogen senyawa, termasuk lemak, minyak, steroid, lilin,
dan senyawa terkait, yang berhubungan lainnya fisik mereka dibandingkan dengan sifat
kimianya. Mereka memiliki milik umum menjadi (1) relatif tidak larut dalam air dan (2) larut
dalam pelarut nonpolar seperti eter dan kloroform. Mereka adalah konstituen makanan
penting bukan hanya karena nilai energi tinggi, tetapi juga karena larut dalam lemak vitamin
dan asam lemak esensial yang terkandung dalam lemak makanan alami. Lemak disimpan
dalam jaringan adiposa, di mana juga berfungsi sebagai termal insula-tor dalam jaringan
subkutan dan di sekitar organ-organ tertentu. Lipid nonpolar bertindak sebagai isolator listrik,
sehingga cepat rambat gelombang depolarisasi sepanjang saraf myelinated. Kombinasi lipid
dan protein (lipoprotein) berfungsi sebagai sarana transportasi lipid dalam darah.
Pengetahuan biokimia lipid diperlukan dalam memahami banyak daerah penting biomedis,
misalnya, obesitas, diabetes mellitus, aterosklerosis, dan peran berbagai asam lemak tak
jenuh ganda di bidang nutrisi dan kesehatan.
Lipid diklasifikasikan sebagai sederhana dan kompleks
1. Lipid sederhana: Ester dari asam lemak dengan berbagai alkohol.
a. Lemak: Ester dari asam lemak dengan gliserol. Minyak adalah lemak dalam
keadaan cair.
b. Lilin: Ester dari asam lemak dengan molekul alkohol monohidrat yang memiliki
berat molekul lebih tinggi.
2. Lipid kompleks: Ester dari asam lemak yang mengandung gugus selain alkohol dan asam
lemak.

a. Fosfolipid: Lipid yang mengandung, selain asam lemak dan alkohol, residu asam
fosfat. Mereka sering telah mengandung nitrogen dasar dan substituen lainnya,
misalnya, dalam gliserofosfolipid alcohol gliserol dan sphingophospholipids alkohol
adalah sphingosine.
b. Glikolipid (glycosphingolipids): Lipid yang mengandung asam lemak, sphingosine,
dan karbohidrat.
c. Lipid kompleks lainnya: Lipid seperti sulfolipids dan aminolipids. Lipoprotein juga
dapat ditempatkan dalam kategori.
3. Prekursor dan turunan lipid: Ini termasuk asam lemak, gliserol, steroid, alkohol lainnya,
aldehida lemak, badan keton , hidrokarbon, vitamin larut lemak, dan hormon.
Metabolisme Lipid
Triasilgliserol atau trigliserida adalah senyawa lipid utama yang terkandung dalam
bahan makanan dan sebagai sumber energi yang penting, khususnya bagi hewan. Sebagian
besar triasilgliserol disimpan dalam sel-sel jaringan adiposa, adipocytes. Triasilgliserol secara
konstan didegradasi dan diresintesis.
Pemrosesan dan distribusi lipid dijelaskan dalam 8 tahap yaitu:
1. Triasilgliserol yang berasal dari diet makanan tidak larut dalam air. Untuk mengangkutnya
menuju usus halus dan agar dapat diakses oleh enzim yang dapat larut di air seperti lipase,
triasilgliserol tersebut disolvasi oleh garam empedu seperti kolat dan glikolat membentuk
misel.
2. Di usus halus enzim pankreas lipase mendegradasi triasilgliserol menjadi asam lemak dan
gliserol. Asam lemak dan gliserol diabsorbsi ke dalam mukosa usus.
3. Di dalam mukosa usus asam lemak dan gliserol disintesis kembali menjadi triasilgliserol
4. Triasilgliserol tersebut kemudian digabungkan dengan kolesterol dari diet makanan dan
protein khusus membentuk agregat yang disebut kilomikron.
5. Kilomikron bergerak melalui sistem limfa dan aliran darah ke jaringan-jaringan.
6. Triasilgliserol diputus pada dinding pembuluh darah oleh lipoprotein lipase menjadi asam
lemak dan gliserol.

7. Komponen ini kemudian diangkut menuju sel-sel target.
8. Di dalam sel otot (myocyte) asam lemak dioksidasi untuk energi dan di dalam sel adipose
(adipocyte) asam lemak diesterifikasi untuk disimpan sebagai triasilgliserol. Selama olah
raga, otot membutuhkan dengan cepat sejumlah energi simpanan. Asam lemak yang disimpan
dalam adipocyte dapat dilepaskan dan ditransport ke myocyte oleh serum albumin untuk
didegradasi menghasilkan energi.
Ada 3 sumber asam lemak untuk metabolisme energi pada hewan, yaitu:
- suplai triasilgliserol dari makanan
- sintesis triasilgliserol dalam hati jika sumber energi internal melimpah
- simpanan triasilgliserol dalam adipocytes.2

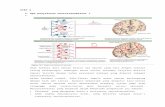
Gambar 3.1 Pemrosesan dan distribusi lipid pada vertebrata. Pencernaan dan absorpsi lipid
dari diet terjadi di usus halus. Asam-asam lemak hasil pengurain trigliserida di pak dan
ditransport ke otot dan jaringan adiposa.
Metabolisme lipid pada jaringan adiposa:
Untuk proses lipogenesis (sintesis lipid) pada jaringan adiposa, triasilgliserol disuplai
dari hati dan usus dalam bentuk lipoprotein, VLDL dan kilomikron. Asam lemak dari
lipoprotein dilepaskan oleh lipoprotein lipase yang berlokasi pada permukaan sel-sel
endotelial pembuluh kapiler darah. Asam lemak kemudian diubah mejadi triasilgliserol.

Proses lipolisis (degradasi lipid) pada jaringan adiposa dikatalisis oleh
Hormonesensitive lipase, yang dikontrol oleh hormon, dengan mobilisasi sebagai berikut
(gambar 3.2):
Gambar 3.2 Mobilisasi triasilgliserol yang disimpan dalam jaringan adiposa.
1. Jika glukosa dalam darah rendah, akan memicu pelepasan epinefrin atau glukagon. Kedua
hormon meninggalkan aliran darah dan mengikat molekul reseptor yang ditemui di dalam
membran adipocyte atau sel lemak.
2. Hal ini menyebabkan adenilat siklase melalui protein G mengubah ATP menjadi cAMP.
3. cAMP kemudian mengaktifkan protein kinase. Protein kinase aktif mengaktifkan
triasilgliserol lipase (Hormone-sensitive lipase) melalui forforilasi.

4. Protein kinase aktif juga mengkatalisis fosforilasi molekul perilipin pada permukaan
butiran lemak (lipid droplet) sehingga triasilgliserol lipase dapat mengakses permukaan
butiran lemak.
5. Selanjutnya triasilgliserol diuraikan menjadi asam lemak bebas dan gliserol oleh
triasilgliserol lipase.
6. Molekul asam lemak yang dihasilkan dilepaskan dari adipocyte dan diikat oleh protein
serum albumin dalam darah untuk diangkut melalui pembuluh darah menuju myocyte (sel
otot) jika dibutuhkan. Jumlah asam lemak yang dilepaskan oleh jaringan adiposa ini
tergantung pada aktivitas triasilgliserol lipase. Hanya asam lemak rantai pendek yang dapat
larut dalam air, sedangkan asam lemak rantai panjang tidak. Oleh karena itu untuk
pengangkutannya asam lemak rantai panjang diikatkan pada serum albumin.
7. Asam lemak tersebut dilepaskan dari albumin dan masuk ke myocyte melalui transport
khusus.
8. Di myocyte asam lemak mengalami ß-oksidasi yang menghasilkan CO2 dan energi ATP.
Degradasi asam lemak di dalam hati
Jaringan menangkap asam lemak dari aliran darah untuk dibangun kembali menjadi
lipid atau untuk memperoleh energi dari oksidasinya. Metabolisme asam lemak intensif
khususnya di dalam sel hati (hepatocytes) Proses terpenting dari degradasi asam lemak
adalah ß-oksidasi yang terjadi di dalam mitokondria. Asam lemak dalam sitoplasma
diaktifkan dengan mengikatkannya pada coenzyme A, kemudian dengan sistem transport
karnitin masuk ke mitokondria untuk didegradasi menjadi acetyl-CoA melalui proses ß-
oksidasi. Residu acetyl hasil dapat dioksidasi lanjut menjadi CO2 melalui TCA dan rantai
respirasi dengan menghasilkan ATP. Jika produksi acetyl-CoA melebihi kebutuhan energi sel
hepatocyte akan diubah menjadi keton bodi untuk mensuplai energi pada jaringan lain. Hal
ini terjadi jika suplai asam lemak dalam plasma darah tinggi, misal dalam kondisi kelaparan
atau diabetes mellitus.
Biosintesis lipid dalam hati.
Biosintesis asam lemak terjadi di sitoplasma, khususnya di hati, jaringan adiposa,
ginjal, paru-paru, dan kelenjar mammae. Pensuplai karbon yang paling penting adalah
glukosa. Akan tetapi prekursor asetyl-CoA yang lain seperti asam amino ketogenik dapat

digunakan. Mula-mula acetyl-CoA dikarboksilasi menjadi malonil CoA, kemudian
dipolimerisasi menjadi asam lemak. Asam lemak selanjutnya diaktivasi dan disintesis
menjadi lipid (triasilgliserol) dengan gleserol 3-fosfat. Untuk mensuplai jaringan lain, lipid
tersebut dipak ke dalam kompleks lipoprotein (VLDL) oleh hepatocyte dan dilepaskan ke
dalam darah.
Rangkuman metabolism lipid
Triasilglisrol dari diet makanan diemulsikan dengan asam empedu di dalam usus
halus, dihidrolisis oleh lipase usus halus menjadi asam lemak dan gliserol, diabsorbsi oleh
sel-sel epitel usus halus, dan diubah kembali menjadi menjai triasilgliserol. Triasilgliserol
kemudian digabungkan dengan chilomicron dan apolipoprotein khusus. Chilomicron
mengangkut triasilgliserol ke jaringan-jaringan, dimana lipoprotein lipase melepaskan asam
lemak untuk masuk ke sel-sel. Triasilgleserol yang disimpan dalam jaringan adiposa
dimobilisasi oleh triasilgliserol lipase (Hormon-sensitive lipase). Asam lemak yang
dilepaskan diikat oleh serum albumin untuk diangkut melalui darah ke jaringanjaringan yang
membutuhkan. Di sel-sel hati asam lemak didegradasi atau disintesis menjadi lipid
Pelly, W John, Edward F. Goljan, Rapid Review Biochemistry, 3rd Ed.
Lehninger, A.L. (1982). Biochemistry, Worth Pub. Inc
Murray, R.K., Granner, D.K., Mayes, P.A., Rodwell, V.W. Lehninger, A.L. (2003). Harper’s
Illustrated Biochemistry, 26th Ed., Lange Medical Books/Mc.Graw-Hill.