laporan Praktikum Evaluasi Nilai Biologis Protein in vitro: Pengukuran Daya Cerna Protein
24
EVALUASI NILAI BIOLOGIS PROTEIN IN VITRO: PENGUKURAN DAYA CERNA PROTEIN Oleh : Golongan P2; Kelompok 1 Nurul Agustina Chandradewi F24090042 Mila Kharisma F24090043 Jian Septian F24090046 Ayu Cahyaning Wulan F24090130 Didiet Rayadi F24061503 Dosen Dr. Puspo Edi Giriwono, S.TP, M.Agr Asisten Praktikum Dede Saputra, S.Pi, M.Si Umi Kulsum, S.TP DEPARTEMEN ILMU DAN TEKNOLOGI PANGAN
-
Upload
jian-septian -
Category
Documents
-
view
1.024 -
download
41
Transcript of laporan Praktikum Evaluasi Nilai Biologis Protein in vitro: Pengukuran Daya Cerna Protein

EVALUASI NILAI BIOLOGIS PROTEIN IN VITRO:
PENGUKURAN DAYA CERNA PROTEIN
Oleh :
Golongan P2; Kelompok 1
Nurul Agustina Chandradewi F24090042
Mila Kharisma F24090043
Jian Septian F24090046
Ayu Cahyaning Wulan F24090130
Didiet Rayadi F24061503
Dosen
Dr. Puspo Edi Giriwono, S.TP, M.Agr
Asisten Praktikum
Dede Saputra, S.Pi, M.SiUmi Kulsum, S.TP
DEPARTEMEN ILMU DAN TEKNOLOGI PANGAN
FAKULTAS TEKNOLOGI PERTANIAN
INSTITUT PERTANIAN BOGOR
2012

1. PENDAHULUAN
1.1 Latar Belakang
Protein merupakan salah satu zat gizi yang dibutuhkan oleh tubuh
dengan porsi yang cukup besar disamping zat gizi lain, seperti karbohidrat.
Keberadaan protein terdapat di berbagai bahan pangan baik hewani maupun
nabati. Protein cukup banyak terkandung di bahan pangan hewani dengan
daya cerna yang lebih baik dibandingkan protein pada bahan pangan nabati.
Namun, terdapat juga bahan pangan nabati yang sarat akan kandungan
protein yang lengkap, seperti kedelai. Kedelai merupakan alternatif sumber
protein yang menjanjikan untuk menggantikan protein hewani. Protein
kedelai mengandung asam amino yang lengkap. Selain itu protein kedelai
mengandung asam amino yang relatif lebih tinggi daripada protein biji-bijian
lainnya, terutama asam amino lisin (FAO 1971).
Selain dikonsumsi secara langsung, terdapat juga berbagai jenis
produk olahan kedelai yang beredar di pasaran, seperti tempe, kecap, dan
minuman bubuk kedelai. Meskipun berbahan dasar sama, masing-masing
bentuk olahan kedelai tersebut memiliki nilai gizi yang berbeda. Secara
keseluruhan, tempe memiliki kadar dan daya cerna protein yang lebih tinggi
di antara produk-produk olahan kedelai lainnya (Sugiyono 2008).
Adanya perlakuan selama pengolahan menyebabkan peningkatan nilai gizi
protein dan ketersediaan zat-zat gizi yang terkandung di dalamnya (Palupi
2007). Hal tersebut disebabkan karena terlepasnya asam amino bebas,
sehingga lebih mudah dicerna oleh tubuh (Astawan 2008). Berdasarkan
pernyataan tersebut, dilakukan analisis lebih lanjut tehadap beberapa produk
olahan kedelai untuk mengetahui daya cerna protein kedelai secara in vitro,
serta membuktikan adanya peningkatan daya cerna terhadap protein kedelai
pada produk olahan kedelai tersebut.
2

1.2 TujuanMengukur daya cerna protein pada beberapa macam sampel produk
dari kedelai dengan menggunakan metode in-vitro dan mengetahui daya cerna
protein melalui analisa penurunan pH sampel setelah mengalami reaksi hidrolisis.
3

2. BAHAN DAN METODE
2.1 Bahan
Bahan-bahan yang digunakan dalam praktikum ini, yaitu kasein, tepung kedelai
mentah, tepung kedelai matang, tepung tempe mentah, tepung tempe matang,
akuades pH 8.0, NaOH 1N, campuran enzim (tripsin, kimotripsin, pankreatin), TCA
0.1M, Na2CO3 0.4M, dan pereaksi Folin.
2.2 Alat
Alat-alat yang digunakan dalam praktikum ini antara lain tabung reaksi bertutup,
gelas ukur, pH-meter, neraca analitik, tabung sentrifus plastik, vortex, pipet Mohr,
sentrifuge, waterbath dan spektrofotometer.
2.3 Prosedur Kerja
Daya cerna protein pada sampel dilakukan secara in vitro dengan
menggunakan campuran enzim (tripsin, kimotripsin, dan pankreatin) yang kemudian
akan dibandingkan dengan daya cerna kasein, sehingga diketahui daya cerna protein
relatif masing-masing sampel. Asam amino yang dihasilkan akibat reaksi enzimatis
kemudian direaksikan dengan pereaksi Folin, sehingga intensitas warna yang
dihasilkan diukur dengan menggunakan spektrofotometer pada panjang gelombang
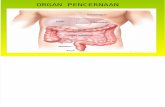
578 nm. Diagram alir prosedur analisis dapat dilihat pada gambar 1.
4

5
1.5 g tepung atau kasein 30 ml akuades pH 8.0
Pengadukan dan penepatan pH 8.0 ± 0.1
Vortex
Pengambilan 10 ml dan pemasukkan ke dalam tabung reaksi bertutup (lakukan 2 set)
Set 1: Penambahan 1.0 ml campuran enzim
Set 2 (sebagai blanko): Penambahan 1.0 ml akuades
Inkubasi 37°C, 10 menit (tepat)
Pengambilan 2.0 ml (homogen)
Penambahan 4.0 ml TCA
Vortex dan sentrifus (3500 rpm, 10 menit)
Pengambilan 1.5 ml supernatan
Penambahan 5.0 ml Na2CO3 Penambahan 1.0 ml reagen Folin
Pendiaman larutan selama 20 menit pada suhu 37 °C
Pengukuran absorbansi pada 578 nm
Gambar 1. Diagram alir penentuan daya cerna protein in vitro
Ukur pH larutan sisa
3. DATA HASIL PERCOBAAN
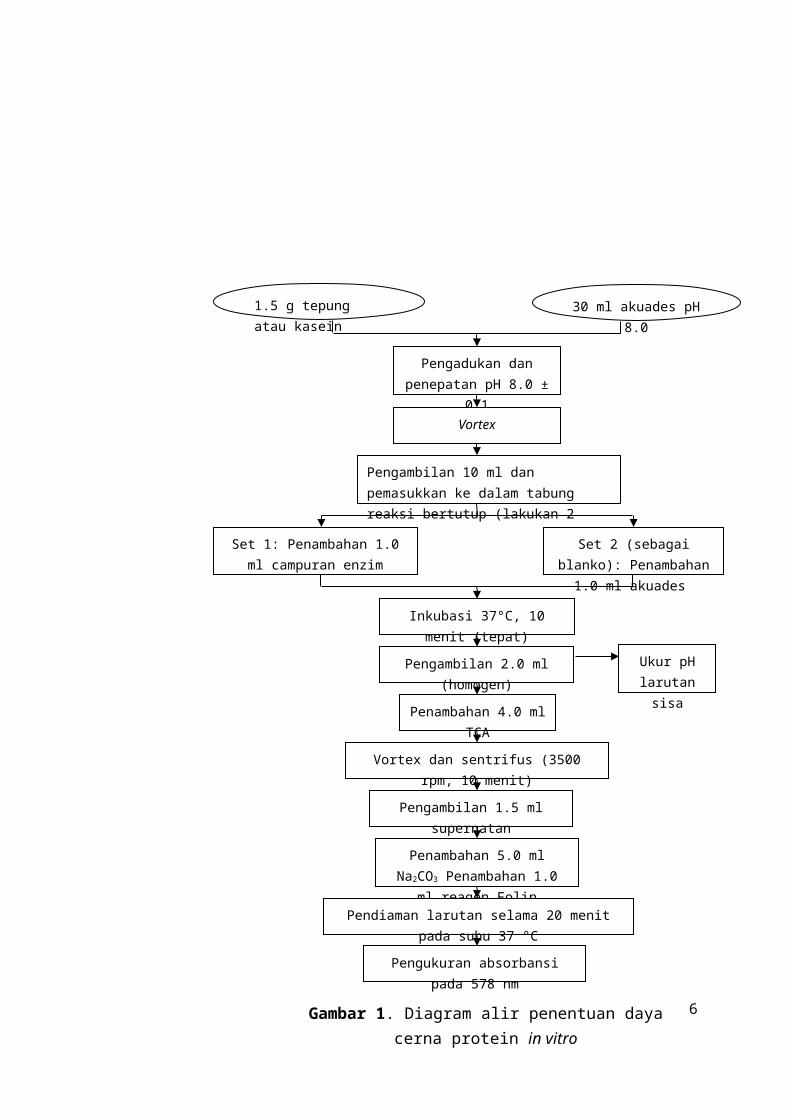
Tabel 1. Data hasil daya cerna protein in vitro
Sampel Absorbansi sampel
Absorbansi blanko
Daya cerna protein relatif (%)
Rata-rata daya cerna protein
relatif (%)
U1 U2 U1 U2 U1 U2Kasein control 1.450 1.467 0.059 0.050 100.00 100.00 100.00
Tepung kedelai mentah 0.940 1.022 0.831 0.722 7.84 21.17 14.50
Tepung kedelai matang 0.620 0.761 0.364 0.497 18.40 18.63 18.52
Tepung kedelai matang 0.682 0.786 0.368 0.395 22.57 27.59 25.08
Tepung tempe mentah 0.569 0.322 0.564 0.359 0.36 -2.61 -1.13
Tepung tempe matang 0.344 0.303 0.218 0.201 9.06 7.20 8.13
Keterangan :
*) : Nilai absorbansi dengan perlakuan pengenceran 2x
U1 : ulangan 1
U2 : ulangan 2
Contoh perhitungan :
Tepung kedelai mentah U1
x 100%
6
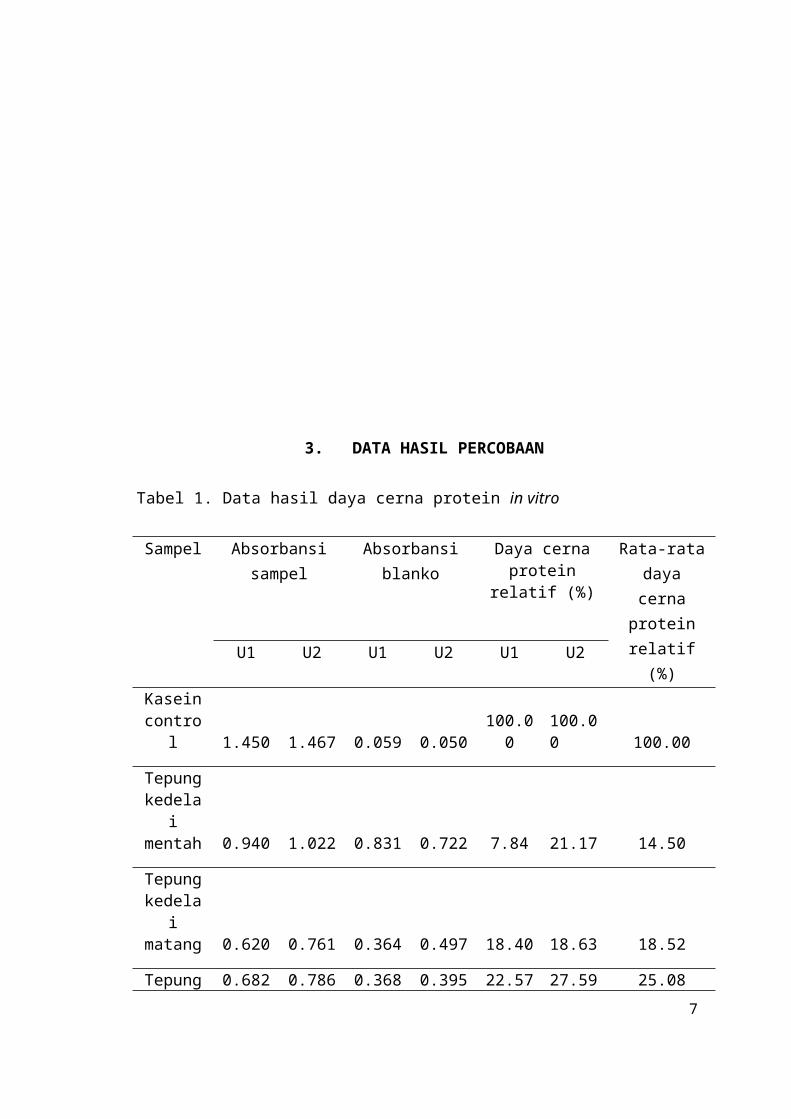
Daya cerna protein tepung kedelai mentah relatif adalah 14.50% terhadap standar (kasein).
Gambar 2. Diagram daya cerna protein pada beberapa sampel produk kedelai
Tabel 2. Data perubahan pH sampel sebelum dan sesudah perlakuan enzim
Sampel pH awalpH sampel setelah perlakuan enzim
Perubahan pH
Kasein (standar) 7.97 7.23 0.74
Tepung kedelai mentah 8.06 7.89 0.17
Tepung kedelai matang 1 8.02 7.09 0.93
Tepung kedelai matang 2 8.03 7.6 0.43
Tepung tempe mentah 8.02 7.6 0.42
Tepung tempe matang 8.02 7.37 0.65
7

Gambar 3. Diagram penurunan pH akibat aktivitas campuran enzim
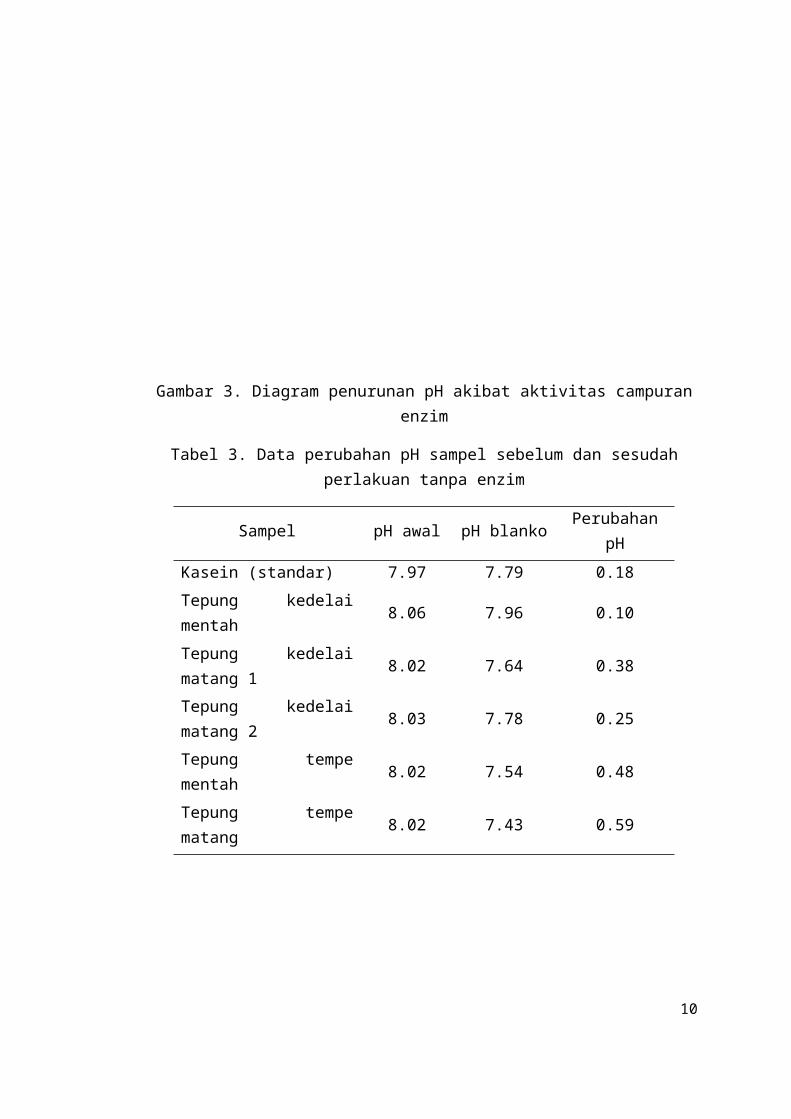
Tabel 3. Data perubahan pH sampel sebelum dan sesudah perlakuan tanpa enzim
Sampel pH awal pH blanko Perubahan pH
Kasein (standar) 7.97 7.79 0.18
Tepung kedelai mentah 8.06 7.96 0.10
Tepung kedelai matang 1 8.02 7.64 0.38
Tepung kedelai matang 2 8.03 7.78 0.25
Tepung tempe mentah 8.02 7.54 0.48
Tepung tempe matang 8.02 7.43 0.59
Gambar 4. Diagram penurunan pH sampel tanpa perlakuan enzim
8
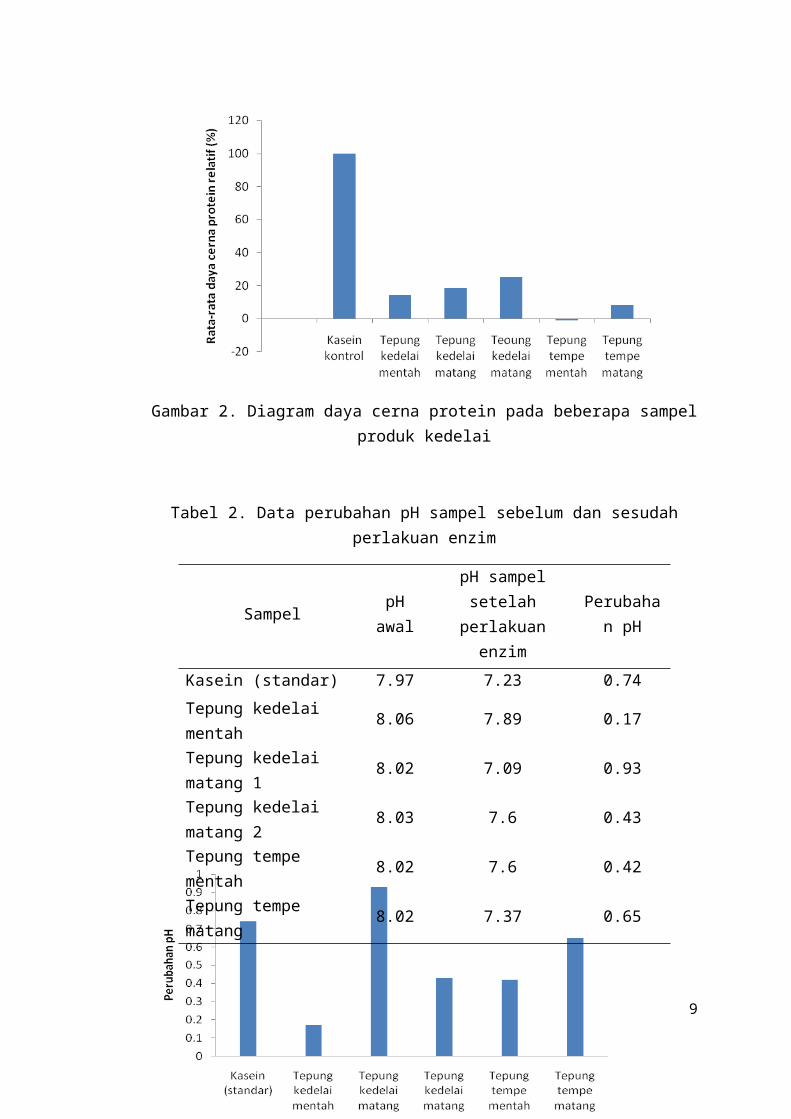
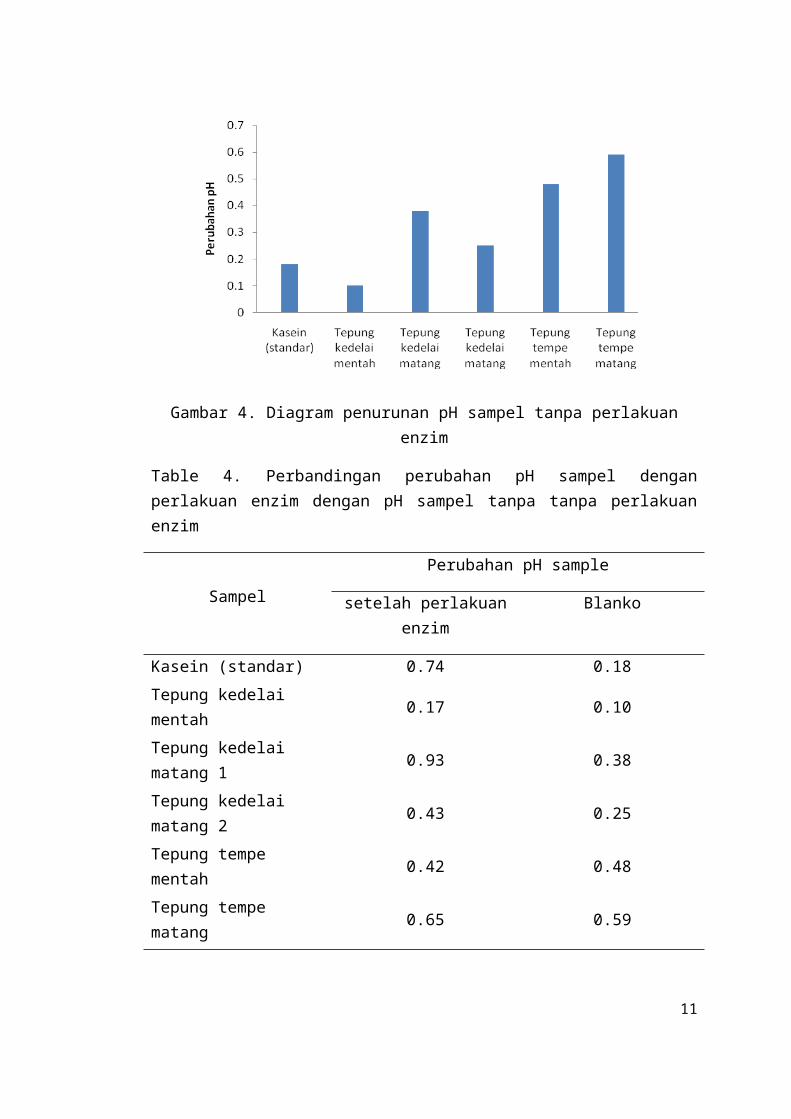
Table 4. Perbandingan perubahan pH sampel dengan perlakuan enzim dengan pH sampel tanpa tanpa perlakuan enzim
SampelPerubahan pH sample
setelah perlakuan enzim Blanko
Kasein (standar) 0.74 0.18
Tepung kedelai mentah 0.17 0.10
Tepung kedelai matang 1 0.93 0.38
Tepung kedelai matang 2 0.43 0.25
Tepung tempe mentah 0.42 0.48
Tepung tempe matang 0.65 0.59
Gambar 5. Diagram perbandingan perubahan pH sampel dengan perlakuan enzim dengan pH sampel tanpa perlakuan enzim
9
4. PEMBAHASAN
Penentuan daya cerna protein secara in vitro dapat dilakukan berdasakan
prinsip absorbansi dengan penggunaan indikator berupa pereaksi Folin yang
memberikan warna pada asam amino dan peptida hasil hidrolisis oleh enzim.
Perbedaan intensitas warna larutan dapat menunjukkan perbedaan kandungan asam
amino dan peptida (protein) pada sampel. Dalam prinsip ini absorbansi berbanding
lurus dengan jumlah asam amino dan peptida dalam larutan. Nilai absorbansi sampel
yang telah dikurangi dengan absorbansi blanko kemudian dibandingkan dengan nilai
absorbansi kasein sebagai kontrol untuk menentukan daya cerna protein tersebut
secara relatif terhadap kasein.
Ada lima sampel yang diuji daya cerna proteinnya pada praktikum ini, antara
lain yaitu sampel tepung kedelai mentah, tepung kedelai matang 1 dan 2, tepung
tempe mentah, dan tepung tempe matang. Kelima sampel tersebut dibandingkan
dengan protein standar, yaitu kasein. Penggunaan kasein sebagai standar dilakukan
dengan alasan kualitas protein kasein yang baik serta merupakan satu protein tunggal
yang lebih mudah dicerna dibandingkan sampel sehingga akan menunjukkan daya
cerna yang baik pula dibandingkan sampel. Kasein didefinisikan sebagai beberapa
kelompok phosphoprotein yang digumpalkan dari susu skim pada pH sekitar 4.6
sampai dengan 4.7 (Damodaran 1996). Kasein komersial umumnya dihasilkan dari
susu skim yang mengalami pengendapan kasein dengan penambahan asam atau
rennet. Komposisi kasein komersial terdiri dari protein 88.5%, lemak 0.2%, air 7%,
dan mempunyai kadar abu 3.8% (Webb et al. 1981).
Kelima sampel yang diuji memiliki pH sekitar 5-6.5, sehingga perlu
ditambahkan NaOH agar pH tepat menjadi 8 dimana pH 8 merupakan standar awal
untuk mengetahui penurunan pH yang terjadi. Pengkondisian pH larutan menjadi 8
bertujuan untuk mendapatkan aktivitas enzim tripsin dan kimotripsin yang
maksimum, karena pH tersebut merupakan pH optimum untuk aktivitas enzim tripsin
dan kimotripsin atau dengan kata lain hal ini dilakukan untuk mengkondisikan
seperti dalam usus manusia. Saat praktikum dilakukan penambahan NaOH karena
dengan penambahan akuades pH 8, larutan sampel masih berada di bawah pH 8.
Suspensi sampel kemudian diberi larutan enzim dan sebagian dibuat sebagai blanko
dengan mengganti larutan enzim dengan akuades.
10
Pembuatan blanko untuk masing-masing sampel bertujuan untuk mengukur
asam amino awal (bukan hasil hidrolisis enzimatis) atau sebagai faktor koreksi
karena dikhawatirkan jika dalam sampel sudah terdapat asam amino bebas sebalum
diberi enzim. Setelah pengkondisian pH optimum enzim, kemudian dilakukan
inkubasi pada suhu 37 oC selama 10 menit. Tujuan dari inkubasi ini adalah untuk
mengkondisikan suhu sampel, dimana suhu ini merupakan suhu yang optimal untuk
aktivitas enzim. Enzim (protease) yang digunakan dalam praktikum ini adalah
tripsin, kimotripsin, dan pankreatin. Setelah sampel dihidrolisis oleh enzim selama
inkubasi 10 menit, sampel diberi TCA (Tri Chloro Acetic acid) untuk mengendapkan
sisa protein dan disentrifuse pada 3500 rpm selama 10 menit sehingga didapatkan
endapan protein dan supernatan. Perlakuan sentrifusa bertujuan untuk mengendapkan
sisa substrat yang bereaksi dengan TCA sehingga supernatant yang didapatkan terdiri
dari asam amino dan peptide. Supernatant diambil sebanyak 1.5 ml lalu ditambahkan
Na2CO3 dan reagen Folin-Ciocalteau.
Reagen Folin-Ciocalteau merupakan campuran asam fosfomolibdat dan asam
fosfotungstat. Reagen direduksi oleh asam amino tirosin dan triptofan (Winarno
1997). Asam amino sistin, sistein, dan histidin juga bisa mereduksi reagen, namun
tidak sekuat tirosin dan triptofan. Reaksi oksidasi-reduksi tersebut diikuti dengan
terbentuknya kompleks warna biru (kromatogen) dengan absorbansi maksimum pada
panjang gelombang 745-750 nm (Nollet 1996). Penambahan garam basa Na2CO3
bertujuan memberikan suasana basa karena pembentukan warna biru dari reagen
Folin-Ciocalteau sangat bergantung pada pH. pH yang paling sesuai adalah 10-10.5,
namun reagen Folin-Ciocalteau tidak stabil pada pH basa sehingga ketepatan waktu
dalam setiap tahap sangat diperlukan (Nollet 1996). Setelah penambahan Na2CO3
dan reagen Folin-Ciocalteau, larutan didiamkan selama 20 menit pada suhu 37 oC
agar reaksi dapat berjalan dengan sempurna. Reaksi yang tidak berjalan sempurna
dapat menyebabkan kesalahan negatif karena ion Cu2+ tidak berikatan secara
maksimum dengan gugus amina. Setelah didiamkan, absorbansi larutan sampel
diukur pada panjang gelombang 578 nm. Semakin tinggi daya cerna protein, semakin
tinggi asam amino yang terbentuk sehingga intensitas warna biru semakin tinggi dan
nilai absorbansi juga semakin tinggi. Nilai absorbansi kemudian digunakan dalam
menghitung daya cerna relatif dari sampel yang diuji.
11
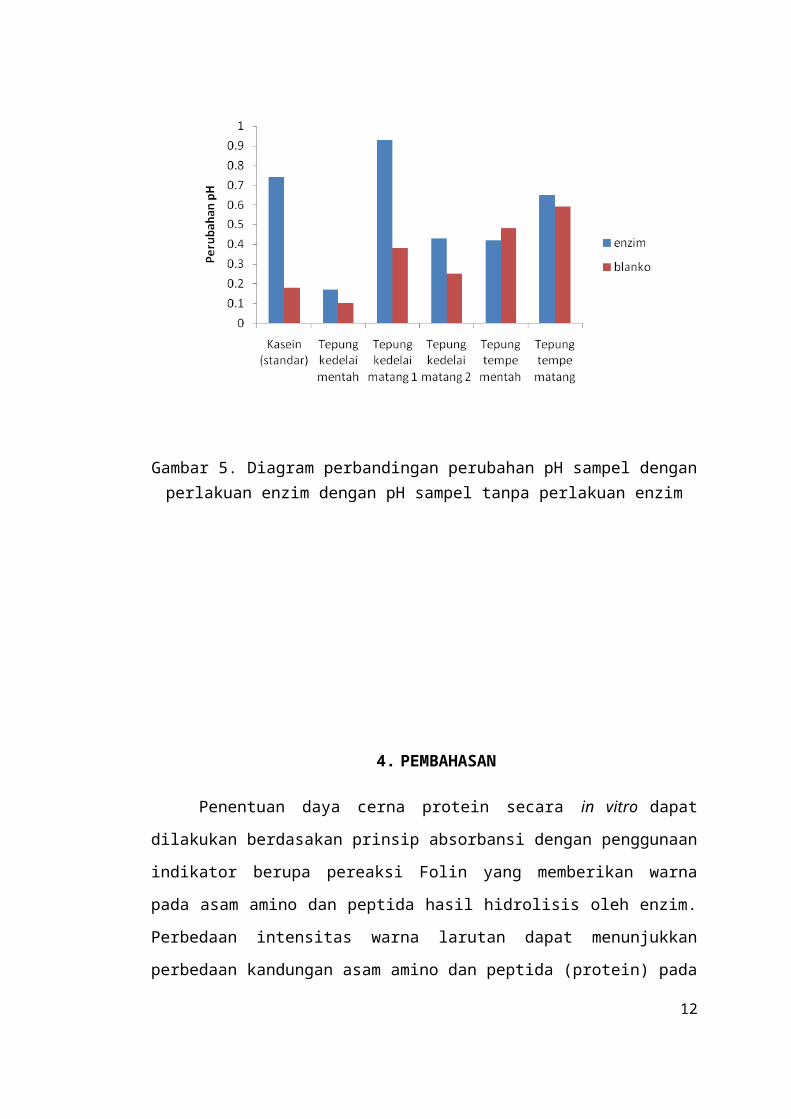
Berdasarkan hasil percobaan, setelah diinkubasi, pH kasein tanpa perlakuan
penambahan enzim mengalami perubahan sebesar 0,18, sedangkan perubahan pH
kasein dengan perlakuan penambahan enzim adalah sebesar 0,74. Hal ini
menunjukkan bahwa penurunan pH cenderung menjadi lebih cepat saat diberikan
tambahan enzim. Hasil analisis perubahan pH pada kelima sampel menunjukkan
bahwa pH tepung kedelai mentah tanpa perlakuan penambahan enzim mengalami
perubahan sebesar 0,10, sedangkan perubahan pH tepung kedelai mentah dengan
perlakuan penambahan enzim adalah sebesar 0,17. Tepung kedelai matang 1 tanpa
perlakuan penambahan enzim mengalami perubahan pH sebesar 0,38, sedangkan
perubahan pH tepung kedelai matang 1 dengan perlakuan penambahan enzim adalah
sebesar 0,93. Untuk tepung kedelai matang 2 tanpa perlakuan penambahan enzim
mengalami perubahan pH sebesar 0,25, sedangkan perubahan pH tepung kedelai
matang 2 dengan perlakuan penambahan enzim adalah sebesar 0,43.
Sampel tepung tempe mentah tanpa perlakuan penambahan enzim mengalami
perubahan pH sebesar 0,48, sedangkan perubahan pH tepung tempe mentah dengan
perlakuan penambahan enzim adalah sebesar 0,42. Sampel tepung tempe matang
tanpa perlakuan penambahan enzim mengalami perubahan pH sebesar 0,59,
sedangkan perubahan pH tepung tempe mentah dengan perlakuan penambahan
enzim adalah sebesar 0,65. Dengan demikian, data besarnya penurunan pH kasein
dan kelima sampel tepung setelah dan sebelum perlakuan enzim dari yang terbesar
hingga yang terkecil secara berturut-turut, yaitu kasein, tepung kedelai matang1,
tepung kedelai matang 2, tepung kedelai mentah, tepung tempe matang, dan tepung
tempe mentah. Secara kualitatif, suatu protein yang lebih cepat melepaskan ion-ion
hidrogen yang ditunjukkan dengan penurunan pH yang lebih cepat dalam kurun
waktu tertentu berarti memiliki daya cerna yang lebih baik. Hal ini menunjukkan
daya cerna protein secara berurutan dari yang paling tinggi hingga yang paling
rendah adalah kasein, tepung kedelai matang 1, tepung kedelai matang 2, tepung
kedelai mentah, tepung tempe matang, dan tepung tempe mentah.
Daya cerna protein dipengaruhi oleh dua faktor, yaitu faktor eksogenus dan
endogenus (Guo et al. 2007). Faktor eksogenus misalnya interaksi protein dengan
polifenol, fitat, karbohidrat, lemak, dan protease inhibitor (Duodu et al. 2003; Ikeda
et al. 1986). Sedangkan faktor endogenus terkait dengan karakterisasi struktur
12

protein seperti struktur tersier, kuartener, serta struktur yang dapat rusak oleh panas
dan perlakuan reduksi (Deshpande dan Damodaran 1989; Ikeda et al. 1991;
Vaintraub et al. 1979). Fennema (1996) mengungkapkan bahwa daya cerna protein
dipengaruhi oleh konformasi protein, ikatan antar protein dengan metal, lipid, asam
nukleat, selulosa atau polisakarida lainnya, faktor anti nutrisi, ukuran dan luas
permukaan partikel protein dan pengaruh proses panas atau perlakuan dengan alkali.
Konformasi protein dapat berhubungan dengan proses pengolahan produk.
Pemanasan merupakan suatu proses termal yang dapat mengubah konformasi protein
(Fennema 1996). Proses pemanasan, seperti perebusan kedelai atau pengeringan
dapat meningkatkan daya cerna protein karena dapat mendenaturasi protein senyawa
anti-nutrisi (anti-protease) (Muchtadi 1993).
Hasil percobaan menunjukkan daya cerna relatif dari tepung kedelai mentah,
tepung kedelai matang 1, tepung kedelai matang 2, tepung tempe mentah, dan tepung
tempe matang terhadap kasein masing-masing adalah 14,50% , 18,52%, 25,08%,
-1,13%, dan 8,13%. Hal ini menunjukkan bahwa sampel yang memiliki daya cerna
protein relatif yang paling tinggi adalah tepung kedelai matang 2, sementara sampel
tepung tempe mentah memiliki daya cerna protein yang paling rendah. Pada
dasarnya, sampel dalam bentuk tepung-tepungan seperti yang diuji pada praktikum
ini akan memiliki daya cerna protein relatif yang tinggi karena perlakuan
pengeringan pada sampel dapat memperluas luas permukaan protein. Hal ini terjadi
karena proses pengeringan akan mengeluarkan air dari protein serta membuat protein
memiliki luas permukaan yang lebih luas dari sebelumnya dikarenakan partikel
protein yang menjadi lebih kecil ketika dikenakan proses pengeringan. Akibatnya,
enzim protease akan lebih mudah untuk menghidrolisis protein (Fennema 1996).
Sampel tepung kedelai matang memiliki daya cerna yang paling tinggi dikarenakan
adanya proses pemanasan dapat meningkatkan ketersediaan zat gizi protein yang
terkandung di dalamnya. Kebanyakan protein pangan terdenaturasi jika dipanaskan
pada suhu yang moderat (60-90 °C) selama satu jam atau kurang.
Rendahnya daya cerna protein relatif pada sampel tepung tempe mentah
maupun tepung tempe matang tidak sesuai dengan hasil penelitian Guo et al. (2007)
yang menyatakan bahwa proses fermentasi menyebabkan tempe memiliki beberapa
keunggulan dibandingkan kacang kedelai. Pada tempe, terdapat enzim-enzim
13

pencernaan yang dihasilkan oleh kapang tempe, sehingga protein, lemak dan
karbohidrat menjadi lebih mudah dicerna. Kapang yang tumbuh pada tempe mampu
menghasilkan enzim protease untuk menguraikan protein menjadi peptida dan asam
amino bebas (Astawan 2008). Ketidaksesuaian ini kemungkinan dipengaruhi oleh
sifat protein sebagai senyawa yang reaktif, dimana sisi aktif beberapa asam amino
dalam protein dapat bereaksi dengan komponen lain misalnya gula pereduksi,
polifenol, lemak dan produk oksidasinya, serta bahan kimia aditif seperti alkali,
belerang dioksida atau hidrogen peroksida (Muchtadi 1993). Selain itu, adanya
kesalahan prosedur praktikum, yaitu waktu inkubasi kedua yang lebih dari 20 menit
serta ketidakstabilan pereaksi Folin juga dapat mempengaruhi hasil uji.
Pereaksi Folin merupakan senyawa yang sangat reaktif dan mudah berikatan
dengan senyawa lain seperti amonium sulfat, sesium bikarbonat, glisin (lebih besar
dari 0,5%), sukrosa, glukosa, EDTA, NaCl, sorbitol, octyl glucoside, chaps, chapso,
lubrol, tris, Triton X-100, dan lain-lain. Amonium sulfat, lubrol, chaps, chapso, dan
sesium bikarbonat merupakan contoh senyawa pengganggu yang dapat
mengendapkan protein. Glisin dan EDTA adalah contoh senyawa pengganggu yang
menyebabkan tidak terbentuknya warna biru pada reaksi (Walker 2002). Selain itu,
merkaptan (2-mercaptoethanol) dan ditiotreitol (DTT) merupakan senyawa
pengganggu yang mereduksi protein untuk bereaksi dengan pewarna. Inkubasi kedua
(setelah penambahan pereaksi Folin) pada sampel tepung tempe mentah dengan
perlakuan penambahan enzim yang lebih dari 20 menit kemungkinan menyebabkan
asam amino glisin pada sampel bereaksi secara berlebihan dengan ion Cu2+ dalam
pereaksi Folin, akibatnya intensitas warna biru yang terbentuk pada sampel tepung
tempe mentah dengan perlakuan penambahan enzim lebih rendah daripada tepung
tempe mentah perlakuan tanpa penambahan enzim, sehingga daya cerna protein
relatif yang terukur bernilai negatif.
14

5. KESIMPULAN
Berdasarkan percobaan yang dilakukan terhadap beberapa sampel tepung
berbahan dasar kedelai dapat diketahui bahwa sampel yang memiliki daya cerna
protein tertinggi berturut-turut adalah kasein, tepung kedelai matang, tepung kedelai
mentah, tepung tempe matang, dan tepung tempe matang. Semakin besar penurunan
pH sample setelah diinkubasi memperlihatkan bahwa sample tersebut memiliki daya
cerna protein yang tinggi, karena penurunan pH yang lebih cepat dalam kurun waktu
tertentu menunjukkan bahwa protein lebih cepat melepaskan ion-ion hidrogen
sehingga lebih banyak protein yang mudah dicerna oleh tubuh. Tepung kedelai
matang memiliki daya cerna protein yang lebih tinggi daripada tepung tempe yang
telah mengalami proses fermentasi. Hal ini dimungkinkan adanya pengaruh oleh sifat
protein sebagai senyawa yang reaktif, dimana sisi aktif beberapa asam amino dalam
protein dapat bereaksi dengan komponen lain misalnya gula pereduksi, polifenol,
lemak dan produk oksidasinya, serta bahan kimia aditif seperti alkali, belerang
dioksida atau hidrogen peroksida.
15

6. DAFTAR PUSTAKA
Astawan M. 2008. Sehat dengan Tempe, Panduan Lengkap Menjaga Kesehatan
dengan Tempe. Jakarta : PT Dian Rakyat
Damodaran S. 1996. Amino Acids, Peptides, and Proteins. Di dalam: Fennema, OR.
(Ed.). Food Chemistry Third Edition. Marcel Dekker Inc, New York.
Deshpande SS, Damodaran S. 1989. Heat induced conformational changes in
phaseolin and its relation to proteolysis. Biochimica et Biophysica Acta (BBA)
– Protein Structure and Molecular Enzymology 998: 179–188.
Duodu KG, Taylor JRN, Belton PS, Hamaker BR. 2003. Factors affecting sorghum
protein digestibility. J of Cereal Sci 38: 117–131.
[FAO] Food and Agriculture Organization. 1971. Technology production from
soybean. Agriculture Service Bulletin, Roma.
Fennema ON. 1996. Food Chemistry Third Edition. Marcel Dekker Inc, New York.
Guo X, Huiyuan Y, Zhengxing C. 2007. Effect of heat, rutin and disulfide bond
reduction on in vitro pepsin digestibility of Chinese tartary buckwheat protein
fractions. J of Food Chem 102:118–122.
Ikeda K, Oku M, Kusano T, Yasumoto K. 1986. Inhibitory potency of plant
antinutrients towards the in vitro digestibility of buckwheat protein. J of Food
Sci 51: 1527–1530.
Muchtadi D. 1993. Teknik Evaluasi Nilai Gizi Protein. Institut Pertanian Bogor,
Bogor.
Owusu RK. 2002. Food Protein Analysis: Quantitative Effects on Processing. NewYork:
Marcel Dekker.
Palupi, Ns, FZ Zakaria, E Prangdimurti. 2007. Pengaruh Pengolahan terhadap Nilai
Gizi Pangan. Modul e-Learning ENBP, Bogor : Departemen Ilmu dan
Teknologi Pangan IPB .
Sugiyono. 2008. K a n d u n g a n G i z i K e d e l a i (terhubung berkala).
http://id.shvoong.com [22 September 2011]
16

Vaintraub IA, Seliger P, Shutov AD. 1979. Action of pepsin on the reserve proteins
of some leguminous seeds. Nahrung 23: 15–21.
Walker JM. 2002. The Protein Protocols Handbook. Totowa: Humana.
Winarno FG. 1997. Kimia Pangan dan Gizi. PT Gramedia Pustaka Utama, Jakarta.
17