
Immunologi lanjutan
22
Imunologi Drs. Saiful Bahri, M.S., Apt.
-
Upload
putridewiindayani -
Category
Documents
-
view
239 -
download
1
description
Immunologi
Transcript of Immunologi lanjutan

Imunologi Drs. Saiful Bahri, M.S., Apt.

SISTEM KOMPLEMENSistem komplemen adalah suatu sistem yang terdiri
dari seperangkat kompleks protein yang satu dengan lainnya sangat berbeda. Pada kedaan normal komplemen beredar di sirkulasi. darah dalam keadaan tidak aktif, yang setiap saat dapat diaktifkan melalui dua jalur yang tidak tergantung satu dengan yang lain, disebut jalur klasik dan jalur alternatif. Aktivasi sistem komplemen menyebabkan interaksi berantai yang menghasilkan berbagai substansi biologik aktif yang diakhiri dengan lisisnya membran sel antigen. Aktivasi sistem komplemen tersebut selain bermanfaat bagi pertahanan tubuh, sebaliknya juga dapat membahayakan bahkan mengakibatkan kematian, hingga efeknya disebut seperti pisau bermata dua. Bila aktivasi komplemen akibat endapan kompleks antigen-antibodi pada jaringan berlangsung terus-menerus, akan terjadi kerusakan jaringan dan dapat menimbulkan penyakit.

KomplemenUnsur pokok sistem komplemen diwujudkan oleh sekumpulan komponen
protein yang terdapat di dalam serum. Protein-protein ini dapat dibagi menjadi protein fungsional yang menggambarkan elemen dari berbagai jalur, dan protein pengatur yang menunjukkan fungsi pengendalian.
Komplemen sebagian besar disintesis di dalam hepar oleh sel hepatosit, dan juga oleh sel fagosit mononuklear yang berada dalam sirkulasi darah. Komplemen C 1 juga dapat di sintesis oleh sel epitel lain diluar hepar. Komplemen yang dihasilkan oleh sel fagosit mononuklear terutama akan disintesis ditempat dan waktu terjadinya aktivasi.
Sebagian dari komponen protein komplemen diberi nama dengan huruf C: C1q, C1r, C1s, C2, C3, C4, C5, C6, C7, C8 dan C9 berurutan sesuai dengan urutan penemuan unit tersebut, bukan menurut cara kerjanya
Komponen C3 mempunyai fungsi sangat penting pada aktivasi komplemen, baik melalui jalur klasik maupun jalur alternatif. Konsentrasi C3 jauh lebih besar dibandingkan dengan fraksi lainnya, hal ini menempatkan C3 pada kedudukan yang penting dalam pengukuran kadar komplemen di dalam serum. Penurunan kadar C3 di dalam serum dapat dianggap menggambarkan keadaan konsentrasi komplemen yang menurun. Juga penurunan kadar C3 saja dapat dipakai sebagai gambaran adanya aktivasi pada sistem komplemen

AKTIVASI KOMPLEMENSistem komplemen dapat diaktifkan melalui dua jalur, yaitu jalur klasik
dan jalur alternatif. Aktivasi tersebut melalui suatu proses enzimatik yang terjadi secara berantai, berarti produk yang timbul pada satu reaksi akan merupakan enzim untuk reaksi berikutnya. Caranya ialah dengan dilepaskannya sebagian atau mengubah bangunan kompleks protein tersebut (pro enzim) yang tidak aktif menjadi bentuk aktif (enzim). Satu molekul enzim yang aktif mampu mengakibatkan banyak molekul komplemen berikutnya. Cara kerja semacam ini disebut the one hit theory.
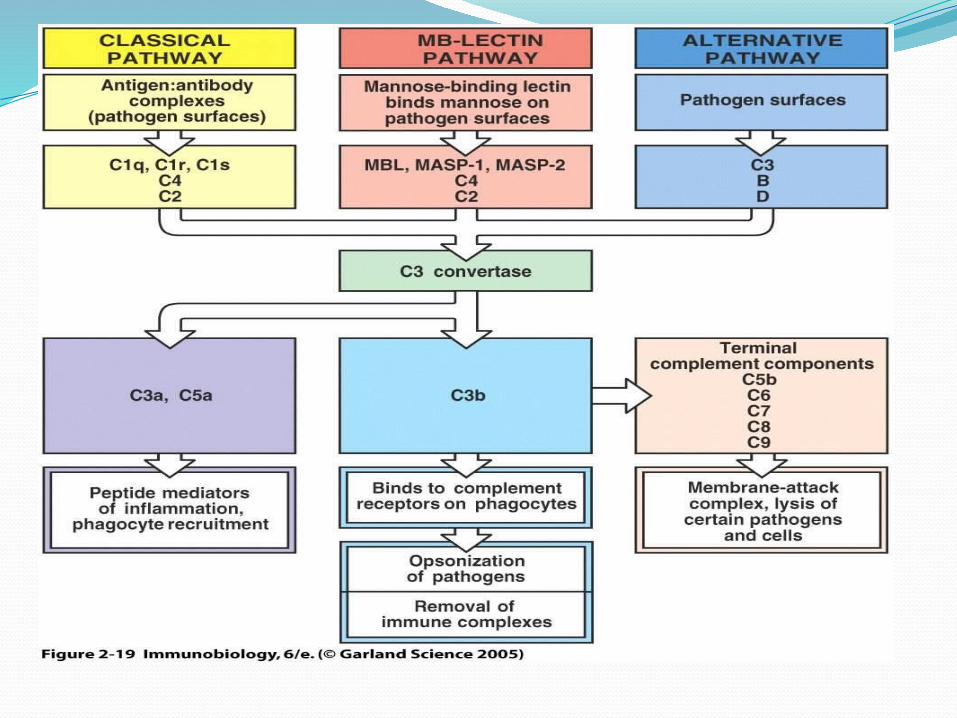
Secara garis besar aktivasi komplemen baik melalui jalur klasik maupun jalur alternatif terdiri atas tiga mekanisme, a) pengenalan dan pencetusan, b) penguatan (amplifikasi), dan c) pengakhiran kerja berantai dan terjadinya lisis serta penghancuran membran sel (mekanisme terakhir ini seringkali juga disebut kompleks serangan membran).
Aktivasi jalur klasik dicetuskan dengan berikatannya C1 dan kompleks antigen-antibodi, sedangkan aktivasi jalur alternatif dimulai dengan adanya ikatan antara C3b dengan berbagai zat aktivator seperti dinding sel bakteri. Kedua jalur bertemu dan memacu terbentuknya jalur serangan membran yang akan mengkibatkan lisisinya dinding sel antigen

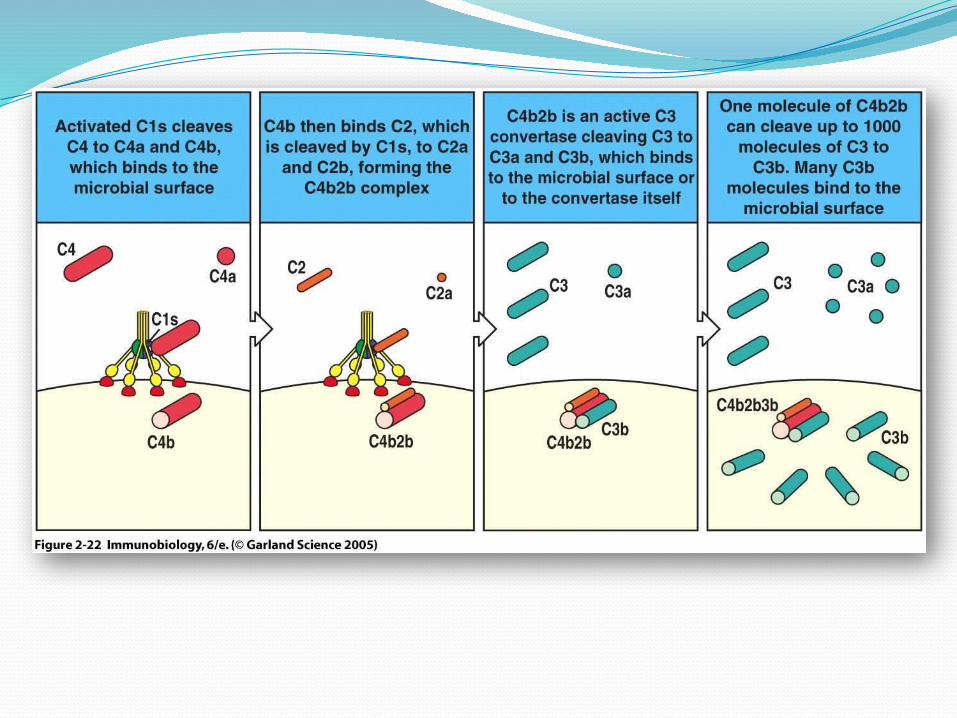
Aktivasi komplemen jalur klasikSeperti telah dibutkan diatas, aktivasi komplemen melalui jalur klasik
atau disebut pula jalur intrinsik, dibagi menjadi 3 tahap. 1. Regulasi jalur klasik Regulasi jalur klasik terutama terjadi
melalui 2 fase, yaitu melalui aktivitas C1 inhibitor dan penghambatan C3 konvertase.
2. Aktivitas C1 inhibitor. Aktivitas proteolitik C1 dihambat oleh
C1 inhibitor (C1 INH). Sebagian besar C1 dalam peredaran darah terikat pada C1 INH. Ikatan antara C1 dengan kompleks antigen-antibodi akan melepaskan C1 dari hambatan C1 INH.
3. Penghambatan C3 konvertase Pembentukan C3 konvertase
dihambat oleh beberapa regulator. C4 binding protein (C4bp) dan reseptor komplemen tipe 1 (CR1)
dapat berikatan dengan C4b sehingga mencegah terbentuknya C4b2b (C3 konvertase). Disamping itu kedua reseptor ini bersama dengan membrane cofaktor protein (MCP) juga dapat meningkatkan potensi faktor I dalam merusak C4b.
Decay accelerating faktor (DAF) dapat berikatan dengan C4b sehingga mencegah terbentulmya C4b2b.

Aktivasi komplemen jalur alternatifAktivasi jalur alternatif atau disebut pula jalur properdin,
terjadi tanpa melalui tiga reaksi pertama yang terdapat pada jalur klasik (C1 ,C4 dan C2) dan juga tidak memerlukan antibodi IgG dan IgM.
Pada keadaan normal ikatan tioester pada C3 diaktifkan terus menerus dalam jumlah yang sedikit baik melalui reaksi dengan H2O2 ataupun dengan sisa enzim proteolitik yang terdapat sedikit di dalam plasma. Komplemen C3 dipecah menjadi frclgmen C3a dan C3b. Fragmen C3b bersama dengan ion Mg++ dan faktor B membentuk C3bB. Fragmen C3bB diaktifkan oleh faktor D menjadi C3bBb yang aktif (C3 konvertase) (Lihat Gambar 5-2). Pada keadaan normal reaksi ini berjalan terus dalam jumlah kecil sehingga tidak terjadi aktivasi komplemen selanjutnya. Lagi pula C3b dapat diinaktivasi oleh faktor H dan faktor I menjadi iC3b, dan selanjutnya dengan pengaruh tripsin zat yang sudah tidak aktif ini dapat dilarutkan dalam plasma.

Tetapi bila pada suatu saat ada bahan atau zat yang dapat mengikat dan melindurlgi C3b dan menstabilkan C3bBb sehingga jumlahnya menjadi banyak, maka C3b yang terbentuk dari pemecahan C3 menjadi banyak pula, dan terjadilah aktivasi komplemen selanjutnya. Bahan atau zat tersebut dapat berupa mikroorganisme, polisakarida (endotoksin, zimosan), dan bisa ular. Aktivasi komplemen melalui cara ini dinamakan aktivasi jalur alternatif. Antibodi yang tidak dapat mengaktivasi jalur klasik misalnya IgG4, IgA2 dan IgE juga dapat mengaktifkan komplemen melalui jalur alternatif.

Jalur alternatif mulai dapat diaktifkan bila molekul C3b menempel pada sel sasaran. Dengan menempelnya C3b pada permukaan sel sasaran tersebut, maka aktivasi jalur alternatif dimulai; enzim pada permukaan C3Bb akan lebih diaktifkan, untuk selanjutnya akan mengaktifkan C3 dalam jumlah yang besar dan akan menghasilkan C3a dan C3b dalam jumlah yang besar pula. Pada reaksi awal ini suatu protein lain, properdin dapat ikut beraksi menstabilkan C3Bb; oleh karena itu seringkali jalur ini juga disebut sebagai jalur properdin. Juga oleh proses aktivasi ini C3b akan terlindungi dari proses penghancuran oleh faktor H dan faktor I.
Tahap akhir jalur alternatif adalah aktivasi yang terjadi setelah lingkaran aktivasi C3. C3b yang dihasilkan dalam jumlah besar akan berikatan pada permukaan membran sel. Komplemen C5 akan berikatan dengan C3b yang berada pada permukaan membran sel dan selanjutnya oleh fragmen C3bBb yang aktif akan dipecah menjadi C5a dan C5b. Reaksi selanjutnya seperti yang terjadi pada jalur altematif (kompleks serangan membran).

FUNGSI BIOLOGIK PROTEIN KOMPLEMENFungsi sistem komplemen pada pertahanan tubuh
dapat dibagi dalam dua golongan besar, 1) lisis sel sasaran oleh kompleks serangan membran, dan 2) sifat biologik aktif fragmen yang terbentuk selama aktivasi.
1. Sitolisis Pada aktivasi sitolisis ini (kompleks
serangan membran) yang berfungsi adalah C5-C9. Mekanisme ini sangat penting bagi pertahanan tubuh melawan mikrooorganisme. Proses lisis ini dapat melalui jalur alternatif maupun jalur klasik.

2. Sifat biologik aktif Opsonisasi dan peningkatan fungsi fagositosisFagositosis yang diperkuat oleh proses opsonisasi
C3b dan iC3b mungkin merupakan mekanisme pertahanan utama terhadap infeksi bakteri dan jamur secara sistemik Fagositosis ini juga lebih meningkat bilamana bakteri disamping berikatan dengan komplemen juga berikatan dengan antibodi IgG atau IgM. Melekatnya antibodi dan fragmen komplemen pada reseptor spesifik yang terdapat pada sel fagosit tidak hanya menyebabkan opsonisasi, tetapi juga memacu untuk terjadinya fagositosis.

Anafilaksis dan kemotaksisC3a, C4a dan C5a disebut anafilatoksin oleh
karena dapat memacu sel mast dan sel basofil untuk melepaskan mediator kimia yang dapat meningkatkan permeabilitas dan kontraksi otot polos vaskular. Reseptor C3a dan C4a terdapat pada permukaan sel mast, sel basofil, otot polos dan limfosit. Reseptor C5a terdapat pada permukaan sel mast, basofil, netrofil, monosit, makrofag, dan sel endotelium.
Melekatnya anafilatoksin pada reseptor yang terdapat pada otot polos menyebabkan kontraksi otot polos tersebut. Untuk mekanisme ini C5a adalah yang paling poten dan C4a adalah yang paling lemah.

C5a juga mempunyai sifat yang tidak dimiliki oleh C3a dan C4a; oleh karena C5a juga mempunyai reseptor yang spesifik pada permukaan sel-sel fagosit maka C5a dapat menarik sel-sel fagosit tersebut bergerak ke tempat mikroorganisme, benda asing atau jaringan yang rusak; proses ini disebut kemotaksis. Juga setelah melekat C5a dapat merangsang metabolisme oksidatif dari sel fagosit tersebut sehingga dapat meningkatkan daya untuk memusnahkan mikroorganisme atau benda asing tersebut.
Proses peradanganKombinasi dari semua fungsi yang tersebut diatas
mengakibatkan terkumpulnya sel-sel dan serum protein yang diperlukan untuk terjadinya proses dalam rangka memusnahkan mikroorganisme atau benda asing tersebut; proses ini disebut peradangan.

Pelarutan dan eliminasi kompleks imunKompleks imun dalam jumlah kecil selalu terbentuk dalam
sirkulasi, dan dapat meningkat secara dramatis bilamana terdapat peningkatan antigen. Kompleks imun ini bilamana berlebihan dapat membahayakan oleh karena dapat mengendap pada dinding pembuluh darah, mengaktivasi komplemen dan menimbulkan kerusakan jaringan. Pembentukan kompleks imun bilamana berlebihan, tidak hanya membutuhkan Fab dari imunoglobulin tetapi juga interaksi dengan Fc. Oleh karena itu pengikatan komplemen pada Fc immunoglobulin suatu kompleks imun dapat membuat ikatan antigen-antibodi yang sudah terbentuk menjadi lemah.
Untuk menetralkan terbentuknya kompleks imun yang berlebihan ini, sistem komplemen dapat meningkatkan fungsi fagosit. Fungsi ini terutama oleh reseptor yang terdapat pada permukaan eritrosit. Kompleks imun yang beredar mengaktifkan komplemen dan mengaktifkan fragmen C3b yang menempel pada antigen. Kompleks tersebut akan berikatan dengan reseptor pada permukaan eritrosit. Pada waktu sirkulasi eritrosit melewati hati dan limpa, maka sel fagosit dalam limpa dan hati (sel Kupffer) dapat membersihkan kompleks imun yang terdapat pada permukaan sel eritrosit tersebut.

REGULASIAktivasi komplemen dikontrol melalui tiga
mekanisme utama, yaitu 1) komponen komplemen yang sudah diaktifkan biasanya ada dalam bentuk yang tidak stabil sehingga bila tidak berikatan dengan komplemen berikutnya akan rusak, 2) adanya beberapa inhibitor yang spesifik misalnya C1 esterase inhibitor, faktor I dan faktor H, 3) pada permukaan membran sel terdapat protein yang dapat merusak fragmen komplemen yang melekat.

Regulasi jalur alternatifJalur altematif juga di regulasi pada berbagai fase
oleh beberapa protein dalam sirkulasi maupun yang terdapat pada permukaan membran.
Faktor H berkompetisi dengan faktor B dan Bb untuk berikatan dengan C3b. Juga CR1 dan DAF dapat berikatan dengan C3b sehingga berkompetisi dengan faktor B. Dengan adanya hambatan ini maka pembentukan C3 konvertase juga dapat dihambat. Faktor I, menghambat pembentukan C3bBb; dalam fungsinya ini faktor I dibantu oleh kofaktor H, CR1 dan MCP. Faktor I memecah C3b dan yang tertinggal melekat pada permukaan sel adalah inaktif C3b (iC3b), yang tidak dapat membentuk C3 konvertase, selanjutnya iC3b dipecah menjadi C3dg dan terakhir menjadi C3d.

Penyakit Dalam Sistem KomplemenPenyakit pada manusia yang berkaitan dengan
sistem komplemen dapat terjadi oleh karena dua keadaan. Pertama adalah adanya defisiensi dari salah satu protein komplemen atau protein regulator. Kedua, suatu sistem komplemen yang normal diaktifkan oleh stimulus yang tidak normal seperti mikroorganisme yang persisten atau suatu reaksi autoimun.
Keadaan itu antara lain :- Defisiensi protein regulator- Defisiensi genetik- Defisiensi komplemen- Defisiensi fragmen kompleks serangan
membran

Defisiensi protein regulator Pada beberapa keadaan dapat terjadi defisiensi protein regulator, baik yang larut maupun yang berikatan pada membran sel. Edema angioneurotik herediter (HANE) adalah suatu keadaan yang disebabkan oleh defisiensi C l INH. Manifestasi klinis kelainan ini adalah edema pada muka, ekstremitas, mukosa laring, dan saluran cerna yang akan menghilang setelah 24 sampai 72 jam. Pada serangan berat disamping gangguan saluran cerna juga dapat terjadi obstruksi saluran nafas. Mediator yang berperan dalam kelainan ini adalah C3a, C4a, dan C5a yang bersifat sebagai anafiltoksin. Di samping itu oleh karena fungsi C l INH juga merupakan regulator kalikrein dan faktor XII, maka kemungkinan aktivasi faktor ini juga memegang peran. Defisiensi regulator jalur alternatif yang larut (faktor H dan I) sangat jarang terjadi. Akibat defisiensi ini C3 akan diaktifkan terus menerus. Pasien dengan antibodi ini sering menderita glomerulonefritis yang mungkin disebabkan oleh kurang adekwatnya pembersihan kompleks imun dari sirkulasi dan mengendap pada membran glomerulus ginjal.

Defisiensi genetik Defisiensi genetik fragmen jalur klasik dan alternatif meliputi C1q, C1r, C1s, C4, C2, C3, properdin, dan faktor D. Defisiensi fragmen awal dari jalur klasik biasanya berhubungan dengan penyakit autoimun seperti glomerulonefritis dan lupus eritematosus sistemik (LES). Yang terbanyak dijumpai pada manusia adalah defisiensi C2. Lebih dari seperdua dari pasien dengan defisiensi C2 dan C4 menderita LES. Pasien dengan defisiensi C2 dan C4 tidak menunjukkan kenaikan frekuensi terkena infeksi. Defisiensi C3 biasanya berhubungan dengan sering terjadinya infeksi bakteri piogen yang fatal. Hal ini mungkin menunjukkan pentingnya peran C3 pada opsonisasi, peningkatan fagositosis, dan penghancuran mikroorganisme. Kenyataan ini menunjukkan bahwa kemungkinan fungsi utama dari jalur klasik adalah untuk eliminasi kompleks imun dan jalur altematif untuk eliminasi bakteri.

Defisiensi komplemen Defisiensi dalam sistem komplemen dapat terjadi pada jalur klasik, altematif, kompleks serangan membran, atau pada protein regulator. Defisiensi ini dapat terjadi sejak lahir, atau didapat setelah lahir oleh karena terdapatnya mutasi gen.
Defisiensi fragmen kompleks serangan membranDefisiensi fragmen kompleks serangan membran yang mencakup C5, C6, C7, C8 dan C9 menyebabkan tidak terdapatnya kemampuan untuk melisis organisme asing. Tetapi kenyataan yang menarik pada pasien dengan defisiensi kompleks serangan membran, hanya mendapat infeksi sistemik yang berat dengan bakteri neiseria intraselular termasuk N. meningitidis dali N. gonorrhoeae. Tetapi oleh karena jumlah sampel pasiennya hanya sedikit, belum dapat disimpulkan bahwa kompleks serangan membran terutarna penting untuk pertahanan terhadap organisme tersebut.

TERIMA KASIH




















