
DESKRIPSI MORFOLOGI, IDENTIFIKASI MOLEKULER DAN … · perkembangan tidak langsung, yaitu dari...
69
DESKRIPSI MORFOLOGI, IDENTIFIKASI MOLEKULER DAN POSISI FILOGENI BERUDU DI PULAU JAWA BERDASARKAN GEN 12S rRNA DAN 16S rRNA LUTHFIA NURAINI RAHMAN SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR BOGOR 2012
Transcript of DESKRIPSI MORFOLOGI, IDENTIFIKASI MOLEKULER DAN … · perkembangan tidak langsung, yaitu dari...

i
DESKRIPSI MORFOLOGI, IDENTIFIKASI MOLEKULER
DAN POSISI FILOGENI BERUDU DI PULAU JAWA
BERDASARKAN GEN 12S rRNA DAN 16S rRNA
LUTHFIA NURAINI RAHMAN
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2012

ii
PERNYATAAN MENGENAI TESIS DAN SUMBER
INFORMASI
Dengan ini saya menyatakan bahwa tesis DESKRIPSI MORFOLOGI,
IDENTIFIKASI MOLEKULER DAN FILOGENI BERUDU DI PULAU JAWA
BERDASARKAN GEN 12S rRNA DAN 16S rRNA adalah hasil karya saya
dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun
kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip
dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah
disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir
tesis ini.
Bogor, Juli 2012
Luthfia N. Rahman
NRP. G352100031

iii
ABSTRACT
Luthfia N. Rahman. Morphological Description, Molecular Identification and
Phylogenetic Relationship of Tadpoles in Java Island Based on 12S rRNA and
16S rRNA Genes. Under supervision of ACHAMAD FARAJALLAH and
MIRZA D. KUSRINI.
Within life cycle of frogs, larval phase is the most difficult phase to
identify, as well as an important phase in the development of Anuran. This study
aims to identify tadpoles using morphological and molecular characteristics and to
assess the phylogenetic of Anuran in Java Island based on tadpoles. Molecular
phylogenetic relationship of tadpoles was estimated using 1362 bp of sequences
from the 12S and 16S rRNA genes. Morphological characteristics showed that
tadpole samples (N=94) were differentiated to 17 species in 6 families. Based on
molecular characteristics there are high similiarty between O. hosii and L.
hasseltii, and between F. limnocharis and F. iskandari. Anuran as monophyletic
group was well-supported except for Rhacophorid based on 12S rRNA gene.
Based on 12S and 16S rRNA genes P. aspera was more closely related to of B.
japonicus, while F. limnocharis was more closely related to F. iskandari based on
12S, 16S and 12S-16S rRNA genes.
Keywords: Identification, tadpole, 12S and 16S rRNA, phylogenetic, Java Islands

iv
RINGKASAN
Luthfia N. Rahman. Deskripsi Morfologi, Identifikasi Molekuler dan Filogeni
Berudu di Pulau Jawa Berdasarkan Gen 12S rRNA dan 16S rRNA. Dibimbing
oleh: ACHAMAD FARAJALLAH, MIRZA D. KUSRINI
Identifikasi spesies pada sebagian besar berudu sulit dilakukan karena
karakter morfologi yang mudah dikenali sangat beragam. Selain itu, berudu dan
Anura dewasa yang ditemukan pada lokasi yang sama tidak mudah untuk saling
dihubungkan terutama pada spesies-spesies yang melakukan aktivitas breeding
dan non-breeding tidak di lokasi yang sama atau berdekatan. Identifikasi spesies
berudu paling sulit dilakukan pada berudu yang berada pada tahap 23 – 25 dalam
proses metamorfosisnya karena merupakan tahap awal diferensiasi oral disc.
Hasil identifikasi spesies berudu dapat menjadi data dasar dalam
manajemen habitat demi kelestarian jenis Anura. Oleh karena itu diperlukan
klustering data molekular yang menjadi alternatif pelengkap untuk memperkuat
identifikasi spesies. Penelitian ini bertujuan untuk mengidentifikasi spesies berudu
di Pulau Jawa berdasarkan karakter morfologi dan molekuler, dan mengetahui
posisi filogeni berudu berdasarkan gen 12S dan 16S rRNA.
Sampel berudu yang dianalisis berjumlah 94 individu. Identifikasi
morfologi dilakukan berdasarkan kunci identifikasi Iskandar (1998). Deskripsi
morfologi dan morfometrika berudu dilakukan berdasarkan Altig (2007), dan
tahap perkembangan berudu ditentukan berdasarkan Gosner (1960). Genom
diekstraksi dari jaringan menggunakan metode fenol-kloroform (Farajallah 2002).
Ruas gen target (12S dan 16S rRNA) diamplifikasi menggunakan pasangan
primer AF05 (5’-ACTGGGATTAGATACCCCACTAT) dan AF08 (5’-
ATGTTTTTGGTAAACAGGCG) dengan suhu annealing 550C. Jarak genetik
antar sampel dihitung berdasarkan jumlah perbedaan nukleotida dan model
subsitusi Kimura 2 Paramater (K2P) menggunakan program MEGA v.4.
Rekonstruksi filogeni dilakukan dengan metode Maximum Parsimony dengan
bootstrap 1000 kali berdasarkan gen 12S rRNA, gen 16S rRNA dan gabungan gen
12S dan 16S rRNA (disimbolkan dengan 12S-16S rRNA).
Identifikasi morfologi menunjukkan bahwa berudu terdiferensiasi dalam
17 spesies dari 6 famili. Berdasarkan nilai terendah dari jarak genetik, hasil
identifikasi morfologi yang kongruen dengan hasil identifikasi molekuler antara
lain P. leucomystax, P. aspera, L. microdiscus, dan R. margaritifer. Spesies yang
hasil identifikasi morfologi tidak kongruen dengan hasil identifikasi molekuler
adalah F. limnocharis, sedangkan spesies yang belum dapat dipastikan adalah O.
hosii.
Percabangan monofiletik pada Anura didukung dengan baik pada
penelitian ini kecuali kelompok Rhacophorid berdasarkan gen 12S rRNA. Spesies
P. aspera berkerabat lebih dekat dengan Bufo berdasarkan gen 12S rRNA dan gen
16S rRNA, sedangkan berdasarkan gen 12S-16S rRNA spesies ini berkerabat
lebih dekat dengan B. japonicus. Spesies F. limnocharis berkerabat lebih dekat
dengan F. iskandari berdasarkan gen 12S rRNA, 16S rRNA dan 12S-16S rRNA.
Kata kunci : Identifikasi, Berudu, 12S dan 16S rRNA, Filogeni, Pulau Jawa

v
@ Hak Cipta milik IPB, Tahun 2012
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan
atau menyebutkan sumber. Pengutipan hanya untuk kepentingan pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau
tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan
yang wajar IPB.
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis
dalam bentuk apapun tanpa izin IPB.

vi
DESKRIPSI MORFOLOGI, IDENTIFIKASI MOLEKULER
DAN FILOGENI BERUDU DI PULAU JAWA
BERDASARKAN GEN 12S rRNA DAN 16S rRNA
LUTHFIA NURAINI RAHMAN
Tesis
Sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada
Program Studi Biosains Hewan
SEKOLAH PASCASARJANA
INTITUT PERTANIAN BOGOR
BOGOR
2012

vii
Penguji Luar Komisi: Dr. Ir. Yeni Aryati Mulyani, M.Sc

viii
Judul Tesis : Deskripsi Morfologi, Identifikasi Molekuler dan Filogeni Berudu
di Pulau Jawa Berdasarkan Gen 12S rRNA dan 16S rRNA
Nama : Luthfia Nuraini Rahman
NIM : G352100031
Mayor : Biosains Hewan
Disetujui,
Komisi Pembimbing
Dr. Ir. Achamad Farajallah, M.Si
Ketua
Dr. Ir. Mirza Dikari Kusrini, M.Si
Anggota
Diketahui,
Ketua Program Studi Biosains Hewan
Dr. Bambang Suryobroto
Dekan Sekolah Pascasarjana IPB
Dr. Ir. Dahrul Syah, M.Sc.Agr
Tanggal ujian : 16 Juli 2012 Tanggal lulus:

ix
PRAKATA
Puji syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya
sehingga tesis ini dapat diselesaikan. Tesis ini disusun dalam rangka memenuhi
persyaratan memperoleh gelar Magister Sains pada Program Studi Biosains
Hewan (BSH) Sekolah Pascasarjana Institut Pertanian Bogor.
Pada kesempatan ini penulis mengucapkan terima kasih kepada:
1. Dr. Achamad Farajallah, M.Si dan Dr. Mirza D. Kusrini, M.Si selaku komisi
pembimbing dan Dr. Yeni A. Mulyani, M.Sc selaku penguji luar komisi yang
telah memberikan arahan dan masukan dalam penyusunan karya ini
2. Kelompok Studi Herpetologi (KSH) Fakultas Biologi Universitas Gadjah
Mada, Yogyakarta atas bantuan spesimen berudu yang diberikan
3. The Mohamed bin Zayed Conservation Fund yang telah memberikan dana
peneltian kepada penulis melalui dana penelitian Dr. Mirza D. Kusrini.
4. Annawaty dan Kamaliah yang telah membantu dalam proses pengumpulan
data di laboratorium
5. Rekan-rekan BSH 2010 yang telah memberikan dukungan dan motivasi
selama perkuliahan berlangsung
6. Kedua orang tua atas doa dan dukungannya yang tidak pernah berhenti kepada
penulis
Semoga karya ilmiah ini bermanfaat bagi perkembangan ilmu pengetahuan.
Bogor, Juli 2012
Luthfia N. Rahman

x
RIWAYAT HIDUP
Penulis dilahirkan di Yogyakarta pada tanggal 29 Juni 1987 dan merupakan
puteri kedua dari pasangan Asep Kusrahman, S.Pd, M.Pd.Si dan Mursilah, S.Pd,
M.Pd Penulis memperoleh gelar Sarjana Kehutanan (S.Hut) dari Departemen
Konservasi Sumberdaya Hutan dan Ekowisata, Fakultas Kehutanan IPB pada
tahun 2010, dan pada tahun yang sama penulis melanjutkan pendidikan Magister
Sains di Program Studi Biosains Hewan, Sekolah Pascasarjana IPB.
Penulis aktif di Kelompok Pemerhati Herpetofauna (KPH)-Phyton
HIMAKOVA IPB sejak tahun 2006 dan terdaftar sebagai anggota Perhimpunan
Herpetologi Indonesia (PHI) sejak tahun 2010.

xi
DAFTAR ISI
Halaman
DAFTAR ISI ......................................................................................................... xi
DAFTAR TABEL .............................................................................................. xiii
DAFTAR GAMBAR .......................................................................................... xiv
I. PENDAHULUAN .............................................................................................. 1
A. Latar Belakang ................................................................................................ 1
B. Tujuan ............................................................................................................. 2
II. TINJAUAN PUSTAKA ................................................................................... 3
A. Anura .............................................................................................................. 3
B. Morfologi Berudu Anura ................................................................................ 4
C. Tahap Perkembangan Berudu ......................................................................... 5
D. Karakter dalam Deskripsi Morfologi Berudu ................................................. 8
E. Kladistik ........................................................................................................ 11
F. Metode Analisis Filogeni .............................................................................. 12
G. Filogenetik Anura ......................................................................................... 13
III. METODE PENELITIAN ............................................................................ 15
A. Waktu dan Tempat Penelitian ...................................................................... 15
B. Sampel Berudu .............................................................................................. 15
C. Deskripsi Morfologi ...................................................................................... 16
D. Ekstraksi dan Isolasi DNA ........................................................................... 16
E. Amplifikasi (Perbanyakan) Ruas DNA......................................................... 17
F. Visualisasi Perbanyakan Ruas DNA ............................................................. 18
G. Perunutan (Sequencing) DNA Produk PCR ................................................. 18
H. Analisis Data ................................................................................................ 18
IV. HASIL ............................................................................................................ 20
A. Identifikasi Morfologi .................................................................................. 20
B. Amplifikasi dan Visualisasi DNA ................................................................ 22
C. Kongruensi Identifikasi Berdasarkan Morfologi dan Genetik ...................... 22
D. Rekonstruksi Filogeni ................................................................................... 24
V. PEMBAHASAN ............................................................................................. 28

xii
A. Kongruensi Identifikasi Morfologi dan Molekuler ....................................... 28
B. Filogenetik ..................................................................................................... 29
VI. KESIMPULAN DAN SARAN ..................................................................... 32
A. Kesimpulan ................................................................................................... 32
B. Saran .............................................................................................................. 32
DAFTAR PUSTAKA .......................................................................................... 33
LAMPIRAN ......................................................................................................... 38

xiii
DAFTAR TABEL
No. Halaman
1. Daftar nama spesies, asal lokasi sampel dan tahun koleksi sampel berudu ...... 15
2. Daftar spesies, wilayah sebaran dan nomor akses GenBank yang digunakan
dalam analisis keragaman nukleotida dan rekonstruksi filogeni ...................... 19
3. Hasil identifikasi morfologi sampel berudu ...................................................... 20
4. Jumlah perbedaan nukleotida dan jarak genetik berdasarkan gen 12S rRNA dan
16S rRNA antar sampel berudu ........................................................................ 23
5. Nilai CI (Consistency Index), RI (Retention index), RC (Rescaled consistency
index) ketiga pohon filogeni Anura .................................................................. 24

xiv
DAFTAR GAMBAR
No. Halaman
1. Morfologi berudu Anura secara umum ............................................................... 4
2. Tahapan perkembangan berudu ........................................................................... 7
3. Berudu tampak dorsal dan lateral ........................................................................ 8
4. Posisi mata berudu ............................................................................................... 9
5. Posisi vent tube pada berudu ............................................................................... 9
6. Contoh oral disc berudu dengan formula geligi I+1-1/III ................................. 10
7. Pohon filogeni secara umum ............................................................................. 12
8. Terminologi dan morfometrika berudu ............................................................. 16
9. Visualisasi amplikon pada PAGE 6% ............................................................... 22
10. Hasil rekonstruksi filogeni berdasarkan gen 12S dan 16S rRNA. .................. 26
11. Hasil rekonstruksi filogeni berdasarkan gabungan gen 12S dan 16S rRNA. .. 27

1
I. PENDAHULUAN
A. Latar Belakang
Anura merupakan salah satu ordo Amfibi yang dicirikan dengan tubuh
tampak seperti berjongkok dengan empat kaki untuk melompat, leher tidak jelas,
tidak berekor dan memiliki permukaan kulit bervariasi dari halus sampai kasar
dengan benjolan-benjolan. Seperti amfibi lainnya, Anura juga mengalami tahapan
perkembangan tidak langsung, yaitu dari embrio menjadi larva kemudian
bermetamorfosis hingga menjadi katak muda (Verma & Pande 2002). Identifikasi
spesies pada sebagian besar larva Anura (yang selanjutnya disebut berudu) sulit
dilakukan. Kesulitan identifikasi berudu menurut Iskandar (1998) berkenaan
dengan karakter morfologi yang mudah dikenali, seperti ukuran tubuh, pola warna
dan formula geligi, sangat beragam. Beberapa berudu diketahui merespon
keberadaan predator dengan mengembangkan sirip ekor menjadi lebih lebar,
terkadang otot ekor menjadi lebih panjang dan badan menjadi lebih kecil. Berudu
dengan keberadaan predator di habitatnya juga memiliki sirip ekor yang berwarna
lebih transparan walaupun respon ini tidak ditemukan pada kebanyakan berudu
(Van Buskirk dan McCollum 1999). Berudu dan Anura dewasa yang ditemukan
pada lokasi yang sama tidak mudah untuk dihubungkan terutama pada spesies-
spesies yang melakukan aktivitas breeding dan non-breeding tidak di lokasi yang
sama atau berdekatan (Starrett 1960).
Selain pada berudu, kesulitan identifikasi juga terjadi pada beberapa
spesies Anura dewasa. Identifikasi spesies Anura dewasa berdasarkan karakter
morfologi tidak berhasil dilakukan pada Fejervarya limnocharis (Dubois 1984,
1987; Inger and Voris (2001). Fejervarya limnocharis merupakan spesies dengan
penyebaran yang sangat luas. Morfologi antar populasi spesies tersebut sulit
dibedakan karena variasi morfologinya kurang dikenali dan tidak adanya diagnosa
spesifik antara spesies pada populasi lokal dan spesies pada populasi lainnya
sehingga seluruh spesies ini dinamai dengan nama yang sama, yaitu F.
limnocharis.
Perkembangan berudu dalam proses metamorfosisnya terjadi dalam 2 fase
(Gosner 1960). Fase pertama disebut fase embrio (tahap 1-25), dimulai dengan

2
pembelahan sel telur hingga embrio menjadi berudu yang hidup bebas. Fase kedua
disebut fase larva (tahap 25-46), dimulai dari berudu yang hidup bebas hingga
menjadi katak muda. Akhir fase 1 (tahap 23-25) merupakan tahap yang paling
sulit dalam mengidentifikasi spesies. Pada tahap ini, oral disc dan barisan geligi
berada pada tahap awal diferensiasi. Barisan geligi terus berkembang secara
bertahap dan biasanya terbentuk sempurna pada awal fase kedua (tahap 25-26),
namun proporsi relatif pada oral disc akan terus berubah secara alometrik selama
metamorfosis. Perubahan proporsi pada oral disc tersebut berhubungan dengan
ekplorasi potensi pakan yang merupakan adaptasi awal berudu dengan habitatnya.
Klustering data molekular merupakan salah satu alternatif pelengkap yang
dapat mempermudah dan memperkuat identifikasi spesies. Penelitian mengenai
identifikasi berudu menggunakan data molekuler dapat digunakan untuk
mengetahui keanekaragaman hayati yang tersembunyi (hidden biodiversity).
Identifikasi spesies berudu terutama dari Anura dewasa yang melakukan aktivitas
breeding dan non-breeding di lokasi yang berbeda dan berjauhan dapat menjadi
data dasar dalam manajemen habitat demi kelestarian jenis Anura.
B. Tujuan
Penelitian ini bertujuan untuk:
1. Mengidentifikasi berudu di Pulau Jawa berdasarkan karakter morfologi
dan molekuler
2. Mengetahui posisi filogeni berudu di Pulau Jawa berdasarkan gen 12S dan
16S rRNA

3
II. TINJAUAN PUSTAKA
A. Anura
Anura merupakan salah satu ordo Amfibi selain Caudata (salamander) dan
Gymnophiona (sesilia). Famili-famili dari Ordo Anura yang terdapat di Indonesia
adalah Bombinatoridae (Discoglossidae), Megophryidae (Pelobatidae),
Bufonidae, Lymnodynastidae, Myobatrachidae, Mycrohylidae, Pelodryadidae
(sering dianggap sebagai anak famili dari Hylidae), Ranidae, Rhacophoridae dan
Pipidae (Iskandar 1998).
Katak mudah dikenal dari bentuk tubuhnya yang tampak seperti
berjongkok dengan empat kaki untuk melompat (kaki depan memiliki empat jari
dan kaki belakang memiliki lima jari), leher tidak jelas dan tidak berekor.
Matanya berukuran besar dengan pupil mata horizontal atau vertikal, ada yang
berbentuk berlian atau segiempat yang khas untuk spesies-spesies tertentu. Kulit
tubuhnya bervariasi dari halus pada beberapa spesies katak, sampai kasar dan
tertutup oleh tonjolan-tonjolan pada spesies katak lainnya. Ukuran katak di
Indonesia bervariasi, dari yang terkecil hanya 10 mm dengan berat 1-2 gram,
sampai dengan spesies yang berukuran 280 mm dan berat lebih dari 1500 gram
(Iskandar 1998).
Beberapa spesies Anura memiliki berudu dengan karakter morfologi yang
berbeda. Spesies Kalophrynus dan Kaloula memiliki berudu yang tidak memiliki
oral disc karena tidak makan sama sekali. Semua energi yang diperlukan untuk
pertumbuhan dan metamorfosis diperoleh dari kuning telur. Berudu Amolops,
Meristogenys dan Huia disebut sebagai berudu Gastromyzophorus, yaitu berudu
yang hidup di air deras. Berudu ini memiliki “mangkuk” penyedot pada ventral
tubuhnya sebagai alat untuk bertahan dari arus yang deras (Inger 1996).
Semua spesies katak mempunyai berudu yang hidup bebas sehingga dapat
diidentifikasi melalui berudu, kecuali spesies Oreophryne dan Philautus.
Keduanya mempunyai telur berukuran besar di mana seluruh fase metamorfosis
berlangsung di dalam telur, sehingga pada akhir metamorfosis telur akan
menetaskan katak muda (Inger 1996).

4
B. Morfologi Berudu Anura
Berudu Anura memiliki tubuh berukuran pendek yaitu kira-kira 25-35%
dari panjang total. Bentuk ekor memipih secara lateral dan terdiri dari sumbu
utama caudal muscular dengan sirip ventral dan dorsal. Sirip ventral memanjang
dari posterior hingga ujung ekor, sedangkan sirip dorsal memanjang dari tengah
atau ujung badan hingga ujung ekor (Gambar 1).
Gambar 1. Morfologi berudu Anura secara umum (Gosner & Rosman 1960)
Tubuh berudu dicirikan dengan bentuk sedikit menggembung, tanpa
kelopak mata, nares lebar dan mulut pada ujung tubuh. Posisi mata, nares dan
mulut bervariasi antar spesies (Duellman & Trueb 1994). Rongga mulut berudu
terdiri dari geligi atas, geligi bawah, papila yang berada pada pinggir mulut dan
keratinous maxilla (Cogger & Zweifel 2003). Berudu memiliki kulit yang tipis,
berlapis-lapis, epidermis tanpa keratin, dan pada area tubuh yang berbeda terdapat
korium yang tebal dengan jaringan ikat (Hofrichter 1999).
Berudu memiliki insang luar pada awal perkembangannya. Insang luar
tersebut dilapisi oleh lipatan operkular yang tumbuh pada posterior dan terbuka ke
sebelah luar melalui spirakel. Letak dan jumlah spirakel bervariasi antar spesies
(Duellman & Trueb 1994).
Orton (1953) dalam Duellman & Trueb (1994) menyatakan bahwa
terdapat empat tipe utama berudu yaitu Tipe I – IV yang tergantung pada struktur
operkular dan bukaannya dari tubuh dan bentuk mulut:
a. Berudu Tipe I; terdiri atas famili Pipidae dan Rhinophrynidae. Berudu tipe ini
memiliki sepasang spirakel; rongga mulut tanpa keratin dan barisan geligi;
mulut memiliki sungut sensorik yang sederhana.

5
b. Berudu Tipe II; yaitu dari famili Microhylidae. Berudu tipe ini memiliki
spirakel tunggal pada posterior median; rongga mulut tanpa keratin dan
barisan; mulut tanpa sungut sensorik
c. Berudu Tipe III; terdiri atas famili Ascaphidae dan Discoglossidae. Berudu
tipe ini memiliki spirakel tunggal pada midventral tubuh; rongga mulut
dengan keratin dan barisan geligi; mulut tanpa sungut sensorik
d. Berudu Tipe IV; merupakan tipe berudu Anura pada umumnya, selain berudu
Tipe I – III. Berudu tipe ini memiliki spirakel sinistral (tunggal pada lateral
kiri tubuh); rongga mulut dengan keratin dan barisan geligi; mulut tanpa
sungut sensorik
C. Tahap Perkembangan Berudu
Gosner (1960) mengelompokkan dan mendeskripsikan perkembangan
berudu dalam 2 fase, yaitu fase embrio dan fase larva (Gambar 2). Setiap
perkembangan didasarkan pada perubahan morfologi dan fungsi tubuh berudu.
Masing-masing fase terdiri dari beberapa tahap, sebagai berikut:
1. Fase Embrio (Tahap 1-25)
Fase ini terdiri dari 25 tahap embrio atau tahap pre-feeding sebelum
berudu keluar dari clutch telur. Tahap awal perkembangan berudu yaitu fertilisasi
(tahap 1), pembelahan zigot (cleavage) menjadi dua, empat, delapan, hingga zigot
menjadi banyak sel (tahap 2-9). Tahap selanjutnya secara berurutan adalah
gastrulasi (tahap 10-12), pembentukan dorsal (tahap 13), pembentukan sistem
saraf (tahap 14-16), pembentukan ujung ekor (tahap 17), pembentukan otot dan
insang (tahap 18), pembentukan jantung (tahap 19). Pada tahap 20 mulai terjadi
sirkulasi pada insang. Perkembangan dilanjutkan dengan terbentuknya kornea
transparan dan mulut terbuka (tahap 21). Pada tahap 22 sirip ekor menjadi
transparan dan mulai terjadi sirkulasi pada sirip. Pada tahap 23 insang luar mulai
tertutup oleh lipatan operkular, bibir dan geligi mulai terdiferensiasi. Pada tahap
24 operkular menutup insang pada sisi kanan tubuh berudu. Pada tahap 25
operkular menutup insang pada sisi kiri tubuh, terbentuk spirakel dan berudu
menetas (keluar dari clutch telur).

6
2. Fase Larva (Tahap 25-46)
Fase ini terdiri dari 21 tahap larva atau tahap berudu hidup bebas setelah
keluar dari clutch telur. Fase larva dimulai pada tahap 25. Tahap selanjutnya
adalah proses pembentukan kaki belakang (hindlimb bud) (tahap 26-30). Tahap ini
kemudian diteruskan dengan proses pembentukan jari kaki (toe development)
(tahap 31-37). Setelah kelima jari kaki terpisah, tuberkula inter-metatarsal
terbentuk (tahap 38). Tuberkula sub-artikular berkembang pada tahap 39.
Tuberkula sub-artikular terbentuk sempurna pada tahap 40. Pada tahap 41 kulit
yang menutupi kaki depan katak menipis dan transparan; bagian dalam rongga
mulut berudu mulai rusak. Pada tahap 42 kaki depan katak keluar; sudut mulut
sejajar nostril; barisan geligi hilang; bentuk mulut berudu menghilang. Tahap
selanjutnya adalah perkembangan mulut katak (mouth development) dan proses
menghilangnya ekor (tail resorption) (tahap 43-45). Tahap 46 adalah tahap akhir
metamorfosis ditandai dengan ekor berudu telah hilang sama sekali.
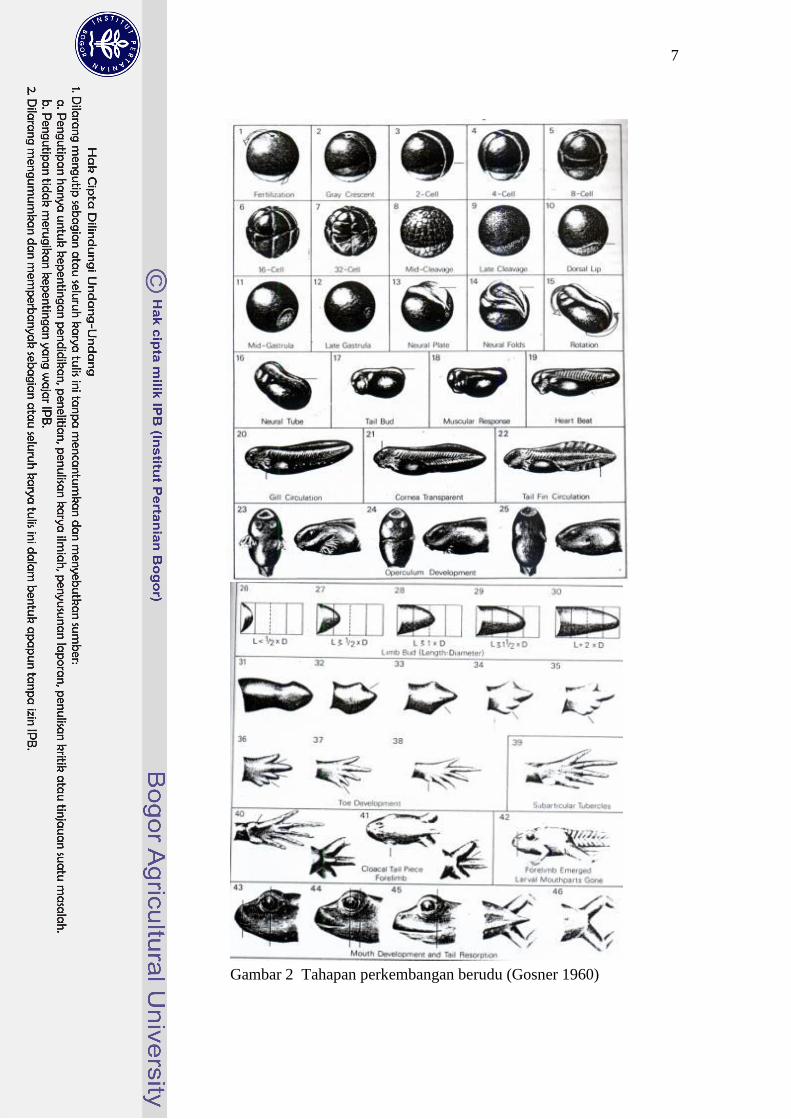
7
Gambar 2 Tahapan perkembangan berudu (Gosner 1960)

8
D. Karakter dalam Deskripsi Morfologi Berudu
Karakter yang digunakan dalam proses deskripsi morfologi berudu adalah
ciri-ciri pada tubuh dan ekor dan bagian mulut berudu. Altig (2007) menjelaskan
terminologi morfologi yang digunakan dalam proses deskripsi, yaitu:
1. Tubuh
a. Tubuh
Berudu terdiri dari tubuh dan ekor, di mana bentuk tubuh berkorelasi
dengan habitat (Gambar 3). Tubuh berudu pada umumnya berbentuk
compressed (lebih tinggi daripada lebar), dan dapat berbentuk depressed
(lebih lebar daripada tinggi) yang biasanya dimiliki oleh jenis-jenis
pemakan lumut dan hidup di aliran deras.
b. Spirakel
Spirakel merupakan lubang yang berfungsi mengeluarkan air dari ruang
buchoparyngeal setelah dipompa masuk ke mulut melalui insang (Gambar
3). Secara umum terdapat 4 konfigurasi spirakel pada berudu, yaitu di
bagian tengah pada dorsal, pada bagian tengah tubuh dekat vent tube,
sinistral (satu pada bagian kiri tubuh, sangat umum), dan dua pada sisi
kanan-kiri tubuh (lateral).
c. Lubang Narial
Lubang narial dimiliki oleh berudu secara umum kecuali pada jenis-jenis
Microhylidae (Gambar 3). Posisi lubang narial dapat lebih dekat dengan
mata daripada mulut, lebih dekat dengan mulut daripada mata, atau berada
di tengah-tengah antara mata dan mulut.
Gambar 3 Berudu tampak dorsal (A) dan lateral (B)
9
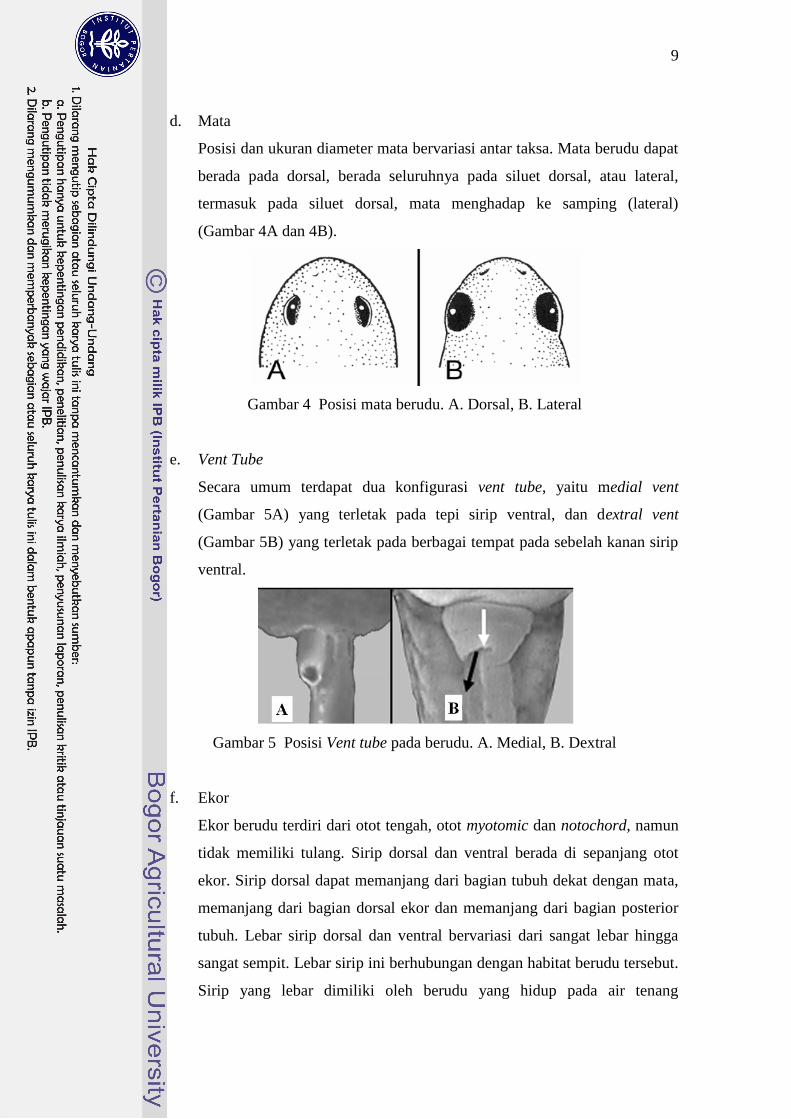
d. Mata
Posisi dan ukuran diameter mata bervariasi antar taksa. Mata berudu dapat
berada pada dorsal, berada seluruhnya pada siluet dorsal, atau lateral,
termasuk pada siluet dorsal, mata menghadap ke samping (lateral)
(Gambar 4A dan 4B).
Gambar 4 Posisi mata berudu. A. Dorsal, B. Lateral
e. Vent Tube
Secara umum terdapat dua konfigurasi vent tube, yaitu medial vent
(Gambar 5A) yang terletak pada tepi sirip ventral, dan dextral vent
(Gambar 5B) yang terletak pada berbagai tempat pada sebelah kanan sirip
ventral.
Gambar 5 Posisi Vent tube pada berudu. A. Medial, B. Dextral
f. Ekor
Ekor berudu terdiri dari otot tengah, otot myotomic dan notochord, namun
tidak memiliki tulang. Sirip dorsal dan ventral berada di sepanjang otot
ekor. Sirip dorsal dapat memanjang dari bagian tubuh dekat dengan mata,
memanjang dari bagian dorsal ekor dan memanjang dari bagian posterior
tubuh. Lebar sirip dorsal dan ventral bervariasi dari sangat lebar hingga
sangat sempit. Lebar sirip ini berhubungan dengan habitat berudu tersebut.
Sirip yang lebar dimiliki oleh berudu yang hidup pada air tenang

10
(genangan, kolam), sedangkan berudu yang hidup pada aliran sedang-deras
memiliki sirip yang sempit.
2. Bagian Oral Disc
Pada sebagian besar taksa, oral disc berudu terdiri dari labial atas
(anterior) dan bawah (posterior) yang masing-masing memiliki tonjolan gigi
melintang dan papilla marjinal dan papilla sub-marjinal pada permukaannya
(Gambar 6). Papila marjinal pada tepi oral disc memiliki tiga tipe, yaitu lengkap
di seluruh tepi oral disc, dengan gap yang lebar pada dorsal (tipe paling umum),
dan dengan gap yang lebar pada tepi dorsal dan ventral. Papilla bervariasi antar
taksa dalam panjang, jumlah dan jumlah baris papilla marjinal. Tepi oral disc
dapat berlekuk (emarginate) (Gambar 6) atau tidak berlekuk (not emarginate).
Gambar 6 Contoh oral disc berudu dengan formula geligi I+1-1/III. Keterangan:
A-1 dan A-2 – baris geligi pertama dan kedua pada bibir atas
(anterior), E – lekuk lateral pada oral disc, G – gap dorsal pada papilla
marjinal, LJS - Lower Jaw Sheaths; bilah rahang bawah, MP – papilla
merjinal, P-1-3 – baris geligi pertama sampai ketiga pada bibir bawah
(posterior), SM – papilla submarjinal, TR – Tonjolan gigi pada baris
geligi P2, dan UJS - Upper Jaw Sheath; bilah rahang atas.

11
Formula geligi merupakan jumlah baris geligi yang berupa garis-garis
hitam pada bibir atas dan bawah berudu. Formula geligi dinyatakan dalam simbol
angka romawi, angka arab, tanda (+), tanda (-) dan tanda (/). Angka romawi
menunjukkan jumlah baris geligi yang utuh, sedangkan angka arab menunjukkan
jumlah baris geligi yang mempunyai celah di tengah. Tanda (+) memisahkan
jumlah baris geligi yang utuh dengan jumlah baris geligi yang memiliki celah di
tengah. Tanda (-) menujukkan bahwa jumlah baris memiliki celah tunggal. Tanda
(/) memisahkan jumlah baris geligi pada bibir atas dan bibir bawah.
E. Kladistik
Kladistik atau sistematika filogenetik adalah metode pengelompokan
organisme berdasarkan karakter turunan yang dimiliki bersama (share derived
characters). Taksa yang memiliki lebih banyak karakter turunan yang sama
berhubungan lebih dekat daripada taksa yang tidak banyak memiliki karakter
turunan. Hubungan antar taksa digambarkan secara hirarkial dalam percabangan
yang disebut Kladogram (Lipscomb 1998).
Analisis filogeni mentransformasi karakter yang homolog dari
sekelompok organisme ke dalam pohon filogeni. Karakter homolog adalah
karakter yang berasal dari karakter nenek moyang yang sama. Sebagai contoh,
sayap pada burung dan kelelawar merupakan karakter yang homolog dengan kaki
depan anjing dan kuda serta tangan pada manusia dan primata. Karakter homolog
tersebut dapat berupa shared primitive characters (Plesiomorf) dan shared
derived characters (Apomorf) (Quicke 1993).
Pohon filogeni terdiri dari akar yang merupakan titik awal atau dasar dari
pohon. Poin yang bercabang disebut nodus internal, sedangkan segmen antara
nodus disebut cabang internal (atau lebih jarang disebut internodus). Taksa yang
ditempatkan di ujung-ujung cabang disebut taksa terminal dan cabang yang
mengarah pada taksa tersebut disebut cabang terminal (Lipscomb 1998) (Gambar
7).

12
Gambar 7 Pohon filogeni secara umum (Lipscomb 1998)
F. Metode Analisis Filogeni
Metode untuk merekonstruksi pohon filogeni dibagi menjadi dua
kelompok besar, yaitu metode jarak (distance) dan metode kriteria (discrete)
(Page & Holmes 1998).
1. Metode Jarak (Distance)
Metode jarak menghitung similaritas jarak antara karakter-karakter yang
ada pada sepasang atau lebih taksa terminal (Quicke 1993). Metode jarak yang
paling banyak digunakan adalah Neighbour Joining (NJ) dan UPGMA
(Unweighted Pair Group Method with Arithmetic means) (Page & Holmes 1998).
Neighbour Joining merupakan metode yang menghitung jarak clustering karakter-
karakter pada dua atau lebih taksa terminal (Page & Holmes 1998). UPGMA
adalah metode berdasarkan rata-rata jarak cluster dari taksa terminal satu dengan
taksa terminal lainnya. Perhitungan jarak rata-rata antara dua cluster dilakukan
dengan menambah jarak secara bersamaan dari semua kemungkinan kombinasi
taksa terminal dan membagi dengan jumlah kombinasi tersebut (Quicke 1993).
2. Metode Kriteria (Discrete)
Metode kriteria yang paling sering digunakan adalah Maximum Parsimony
(MP) (Page & Holmes 1998). Maximum Parsimony menilai karakter yang
parsimoni untuk menentukan posisi taksa terminal dalam pohon dan mencari
semua kemungkinan topologi (bentuk) pohon untuk mendapatkan pohon yang
memiliki langkah paling pendek (Kitching et al. 1998).

13
G. Filogenetik Anura
Beberapa peneliti telah melakukan penelitian mengenai hubungan
filogenetik Anura baik pada spesies Indonesia maupun spesies dunia. Penelitian-
penelitian tersebut antara lain:
1. Emerson et al. (2000), meneliti hubungan filogeni Katak Bertaring
(Limnonectes) di Asia Tenggara berdasarkan gen 12S dan 16S rRNA. Hasil
penelitian ini menunjukkan bahwa Limnonectes merupakan kelompok
monofiletik yang terdiri dari 4 kelompok besar, yaitu kelompok L. kuhlii dan
kerabatnya, kelompok L. leporinus yang terdapat di Kalimantan, kelompok
yang tersebar di Filipina dan Sulawesi dan kelompok L. blythii dan
kerabatnya
2. Liu et al. (2000), meneliti hubungan filogenetik Bufonidae Asia Timur
berdasarkan DNA mitokondria. Hasilnya menunjukkan bahwa Bufonidae
Asia Timur mengelompok menjadi 2 klad utama, klad pertama terdiri dari
B. andrewsi, B. bankorensis, B. tibetanus, B. gargarizans, B.tuberculatus,
klad sisternya B. cryptotympanicus, dan 2 spesies Torrentophryne, dan klad
kedua terdiri dari B. galeatus, B. himalayanus.
3. Hillis dan Wilcox (2005), meneliti hubungan filogeni katak genus Rana di
Dunia Baru berdasarkan gen 12S dan 16S rRNA. Hasilnya menunjukkan
bahwa spesies-spesies Rana Amerika tidak monofiletik; spesies Amerana di
daerah sebelah barat Amerika Utara lebih dekat dengan kelompok R.
temporaria dari Eurasia. Spesies Amerika lainnya membentuk kelompok
Novirana.
4. Yu et al. (2009), memeriksa ulang hubungan filogenetik Rhacophoridae dari
penelitian Li et al. (2008) berdasarkan DNA mitokondria dan DNA inti. Hasil
yang diperoleh menunjukkan bahwa genus Feihyla dan Chiromantis
merupakan sister taksa dari klad yang dibentuk oleh kelompok Feihyla,
Polypedates, dan Rhacophorus. Spesies Aquixalus odontotarsus ditempatkan
ke dalam Kurixalus, spesies Aquixalus lainnya dan Philautus jinxiuensis
dimasukkan ke dalam genus Gracixalus.
5. Matsui et al. (2010) merekonstruksi hubungan filogenetik Leptobrachium
berdasarkan DNA mitokondria. Hasilnya menunjukkan bahwa

14
Leptobrachium merupakan kelompok monofiletik dengan dua klad utama.
Klad pertama terdiri dari 3 sub-klad yang beranggotakan spesies dari 3 daerah
terpisah yaitu Borneo, Semenanjung Malaya dan Jawa, dan Thailand. Sub-
klad Borneo terdiri dari satu spesies dari Filipina dan Sumatra. Klad kedua
terdiri dari dua sub-klad yaitu dari Indocina dan lainnya dari Cina bagian
selatan (Vibrissaphora).
6. McLeod (2010) menyusun sistematika spesies kriptik: Limnonectes kuhlii
complex berdasarkan gen 12S rRNA, tRNA-valin, dan 16S rRNA. Hasil
menunjukkan bahwa apa yang selama ini diakui secara historis sebagai satu
spesies ternyata merupakan kompleks lebih dari 22 keturunan evolusi yang
berbeda, 16 di antaranya saat ini dimasukkan ke dalam L. kuhlii. Hasil
penelitian ini juga mendeteksi adanya kasus keturunan simpatrik, dan pada
seluruh kasus simpatrik tersebut, masing-masing spesies tidak berhubungan
dekat.
7. Hamidy et al (2011), meneliti ketidakselarasan hubungan antara morfologi
dan genetik pada dua spesies Leptopbrachium di Borneo. Hasilnya
memperlihatkan bahwa spesies L. hasseltii dan L. chapaense keturunan
Borneo, Filipina dan Sumatera membentuk klad monofiletik bersama spesies
L. lumadorum dari Mindanao sebagai klad basal. Sementara dua spesies
Filipina lainnya dari Pelawan dan Mindoro membentuk klad dan lebih dekat
dengan keturunan Borneo. Hubungan filogenetik tersebut tidak sama dengan
variasi morfologi yang ada. Spesies L. montanum dan salah satu keturunan
dari L. abbotti secara genetik sangat dekat namun memiliki perbedaan yang
sangat jelas pada pola warna ventral.
15
III. METODE PENELITIAN
A. Waktu dan Tempat Penelitian
Penelitian dilakukan pada bulan Januari – April 2012 di Laboratorium
Molekuler Departemen Biologi FMIPA IPB.
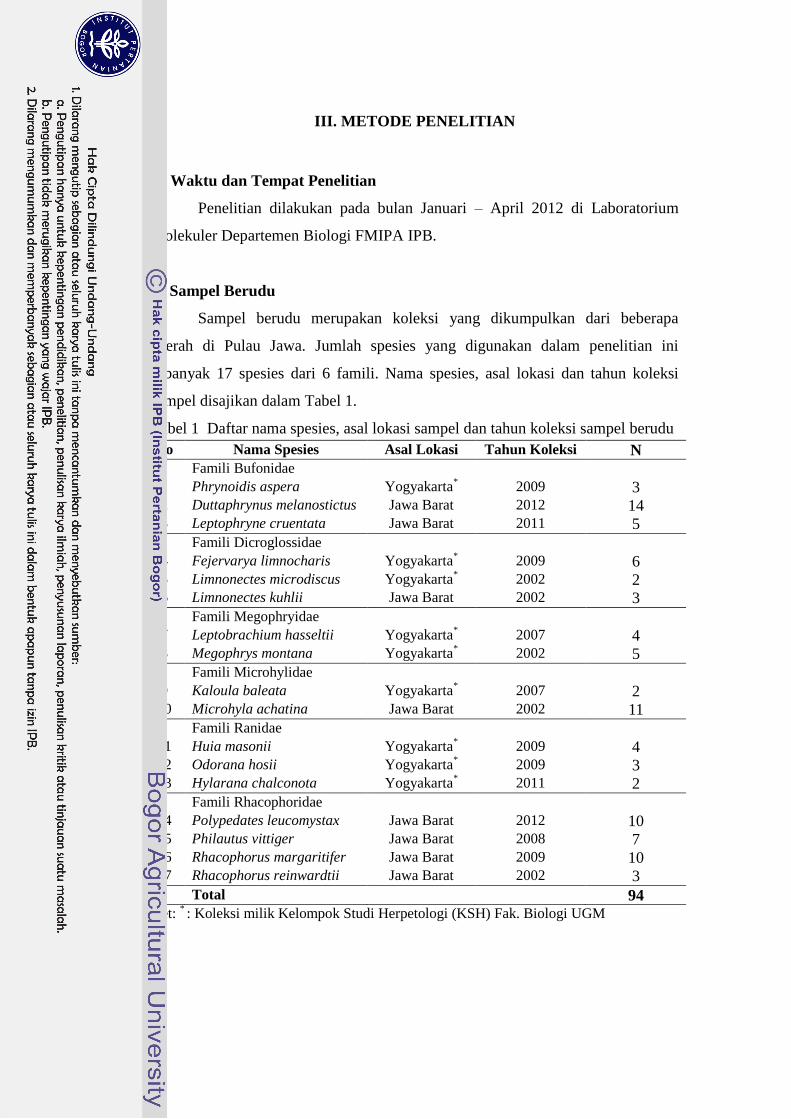
B. Sampel Berudu
Sampel berudu merupakan koleksi yang dikumpulkan dari beberapa
daerah di Pulau Jawa. Jumlah spesies yang digunakan dalam penelitian ini
sebanyak 17 spesies dari 6 famili. Nama spesies, asal lokasi dan tahun koleksi
sampel disajikan dalam Tabel 1.
Tabel 1 Daftar nama spesies, asal lokasi sampel dan tahun koleksi sampel berudu
No Nama Spesies Asal Lokasi Tahun Koleksi N Famili Bufonidae
1 Phrynoidis aspera Yogyakarta*
2009 3 2 Duttaphrynus melanostictus Jawa Barat 2012 14 3 Leptophryne cruentata Jawa Barat 2011 5 Famili Dicroglossidae
4 Fejervarya limnocharis Yogyakarta* 2009 6
5 Limnonectes microdiscus Yogyakarta* 2002 2
6 Limnonectes kuhlii Jawa Barat 2002 3 Famili Megophryidae
7 Leptobrachium hasseltii Yogyakarta* 2007 4
8 Megophrys montana Yogyakarta* 2002 5
Famili Microhylidae 9 Kaloula baleata Yogyakarta
* 2007 2
10 Microhyla achatina Jawa Barat 2002 11 Famili Ranidae
11 Huia masonii Yogyakarta* 2009 4
12 Odorana hosii Yogyakarta* 2009 3
13 Hylarana chalconota Yogyakarta* 2011 2
Famili Rhacophoridae 14 Polypedates leucomystax Jawa Barat 2012 10 15 Philautus vittiger Jawa Barat 2008 7 16 Rhacophorus margaritifer Jawa Barat 2009 10 17 Rhacophorus reinwardtii Jawa Barat 2002 3
Total 94 Ket:
* : Koleksi milik Kelompok Studi Herpetologi (KSH) Fak. Biologi UGM

16
C. Deskripsi Morfologi
Pengamatan morfologi berupa pola warna tubuh dan ekor, letak mata,
nostril, anal, mulut dan geligi dilakukan dengan mikroskop stereo. Pengukuran
morfometri seluruh individu berudu dilakukan untuk mendapatkan variasi ukuran
tubuh berudu. Tahap perkembangan berudu ditentukan berdasarkan Gosner
(1960). Terminologi dan pengukuran morfometri berudu dilakukan berdasarkan
Altig (2007), yaitu: BL - body length, IND - internarial distance, IOD -
interorbital distance, LB - limb bud, MTH - maximum tail height, OD - oral disc,
SP - spiracle, TAL - tail length, TL - total length, TMA – tail muscle axis, TMH -
tail muscle height, TMW - tail muscle width, and VT - vent tube (Gambar 8).
Gambar 8. Terminologi dan morfometrika berudu; (A) dorsal dan (B) lateral
D. Ekstraksi dan Isolasi DNA
Isolasi DNA total dilakukan mengikuti metode Farajallah (2002).
Potongan otot berudu sepanjang ± 2 mm dalam alkohol 70% dicuci dengan 500 µl
buffer TE sebanyak 2 kali. Sampel yang telah dicuci tersebut kemudian
dihancurkan menggunakan gunting dalam buffer 1×STE 300 µl. Penghancuran
protein dilakukan dengan menambahkan enzim Proteinase-K sebanyak 20 µl dan
10% SDS sebanyak 50 µl. Campuran tersebut kemudian diinkubasi pada suhu
550C selama 1 jam sambil dikocok perlahan.
Material DNA dipisahkan dari material organik lainnya dengan metode
ekstraksi fenol, yaitu dengan menambahkan larutan NaCl 5M sebanyak 50 µl,
fenol sebanyak 400 µl dan kloroform-isoamil alkohol (CIAA; 24:1) sebanyak 400

17
µl. Campuran tersebut kemudian diinkubasi pada suhu ruang sambil dikocok
pelan selama 1 jam. Bahan organik yang masuk ke fase fenol dipisahkan dari fase
air dengan sentrifugasi pada kecepatan 5000 rpm selama 5 menit. Fase air yang
terbentuk di lapisan atas kemudian dipindahkan ke dalam tabung baru. DNA
dimurnikan dengan teknik pengendapan alkohol yaitu dengan menambahkan
alkohol absolut sebanyak 2× volume fase air yang dipindahkan dan NaCl 5M
sebanyak 1/10 × volume fase air yang dipindahkan. Campuran diinkubasi pada
suhu 40C selama 24 jam. Molekul-molekul DNA diendapkan dengan sentrifugasi
pada kecepatan 5000 rpm selama 5 menit. Endapan DNA yang diperoleh
kemudian dicuci dengan alkohol 70%. Setelah alkohol pencuci dibuang dan
dievaporasi dalam ruang vakum selama 30 menit, molekul-molekul DNA
disuspensikan dalam buffer TE 80% dan disimpan dalam freezer untuk digunakan
lebih lanjut.
E. Amplifikasi (Perbanyakan) Ruas DNA
Ruas DNA diamplifikasi dengan metode Polymerase Chain Reaction
(PCR) menggunakan mesin Thermo Cycler TaKaRa MP4. Primer yang digunakan
adalah AF05 (5’-ACTGGGATTAGATACCCCACTAT) dan AF08 (5’-
ATGTTTTTGGTAAACAGGCG).
Primer AF05 menempel pada bagian tengah gen 16S rRNA dan AF08
menempel pada bagian akhir gen 12S rRNA. Kedua gen tersebut mengapit ruas
DNA sebesar 1474 pb atau setara dengan posisi nukleotida 505 sampai dengan
1978 genom mitokondria Dogania subplana (Farajallah 2002).
Reaksi PCR dilakukan dalam volume 25µl dengan komposisi 2 µl cetakan
DNA, Distilated water steril 15.3 µl, primer 10 ρmol/ µl masing-masing 1 µl,
dNTP 2.5mM sebanyak 1.5 µl, MgCl2 sebanyak 1.5 µl dan Taq Polymerase 1.25
u/µl sebanyak 0.2 µl beserta bufernya sebanyak 2.5 µl. Amplifikasi DNA
dilakukan dalam kondisi suhu pre-denaturasi 940C selama 4 menit, dilanjutkan
dengan 30 siklus yang terdiri dari denaturasi 940C selama 1 menit, penempelan
550C selama 1 menit 30 detik, pemanjangan 72
0C selama 1 menit dan
pemanjangan akhir 720C selama 7 menit.

18
F. Visualisasi Perbanyakan Ruas DNA
Amplikon diuji menggunakan metode elektroforesis gel poliakrilamid
(PAGE) 6% yang dijalankan pada tegangan 200 V selama 35 menit atau sampai
pewarna bromthymol blue mencapai bagian bawah gel. Setelah pemisahan
elektroforesis, pita-pita DNA divisualisasi dengan pewarnaan perak (silver
staining) (Tegelstrom 1986).
G. Perunutan (Sequencing) DNA Produk PCR
Perunutan merupakan tahap akhir untuk memperoleh data runutan
nukleotida dari ruas DNA amplikon. Perunutan DNA dilakukan menggunakan
jasa pelayanan sekuensing dari PT. Macrogen Inc. Perunutan tersebut dilakukan
menggunakan pasangan primer yang digunakan pada saat PCR.
H. Analisis Data
Morfologi berudu masing-masing spesies disajikan dalam bentuk
deskripsi. Urutan nukleotida yang diperoleh dari proses perunutan diedit secara
manual berdasarkan kromatogram menggunakan program BioEdit Sequence
Alignment Editor 6.32. Runutan nukleotida yang telah diedit kemudian
disejajarkan bersama beberapa spesies Anura lainnya yang diakses dari GenBank
dengan nomor akses terdapat pada Tabel 2. Proses penyejajaran dilakukan dengan
program Clustal W kemudian diedit secara manual.
Analisis jumlah perbedaan nukleotida, jarak genetik dengan model Kimura
2 Parameter (K2P) dan rekonstruksi filogeni dilakukan menggunakan program
MEGA version 4 (Tamura et al. 2007).
Rekonstruksi filogeni dilakukan dengan metode Maximum Parsimony
(MP) berdasarkan gen 12S rRNA, 16S rRNA dan gabungan antara 12S dan 16S
rRNA (selanjutnya disimbolkan dengan gen 12S-16S rRNA). Outgroup yang
digunakan dalam rekonstruksi filogeni adalah Crocodylus porosus. Analisis
bootstrap 1000 kali dilakukan untuk menguji tingkat kepercayaan dari sebuah titik
cabang dalam topologi pohon filogeni.
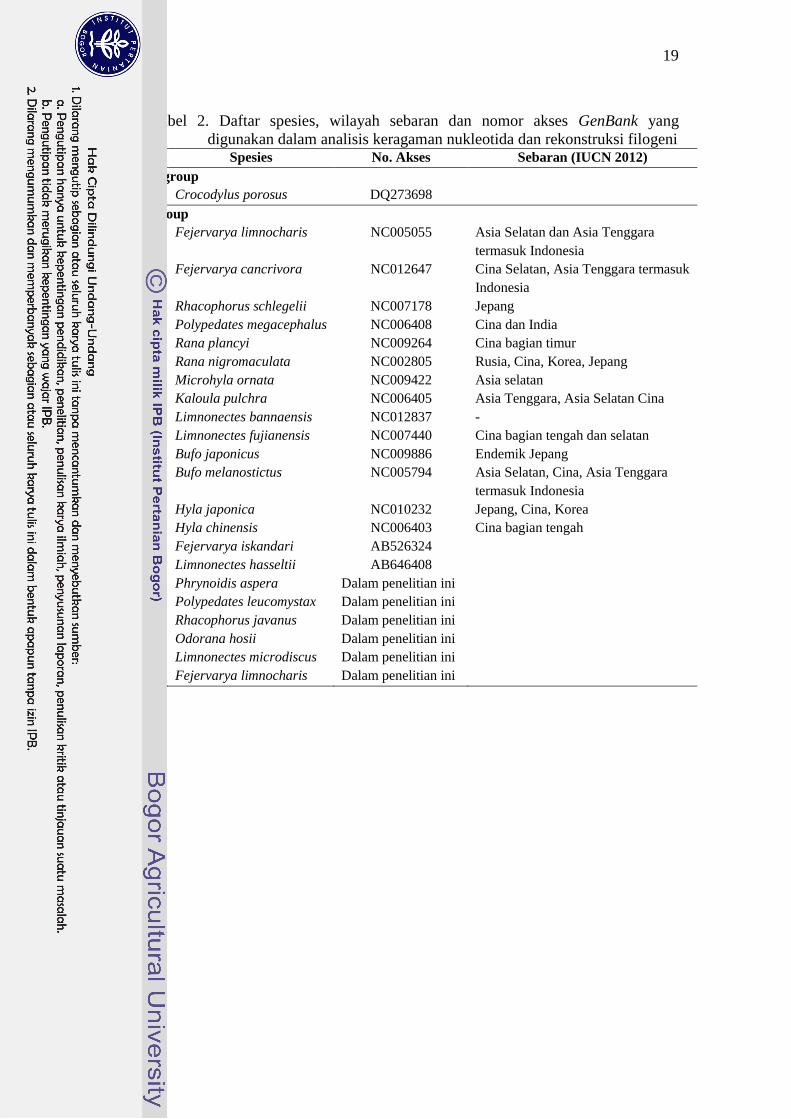
19
Tabel 2. Daftar spesies, wilayah sebaran dan nomor akses GenBank yang
digunakan dalam analisis keragaman nukleotida dan rekonstruksi filogeni No Spesies No. Akses Sebaran (IUCN 2012)
Outgroup
Crocodylus porosus DQ273698
Ingroup
1 Fejervarya limnocharis NC005055 Asia Selatan dan Asia Tenggara
termasuk Indonesia
2 Fejervarya cancrivora NC012647 Cina Selatan, Asia Tenggara termasuk
Indonesia
3 Rhacophorus schlegelii NC007178 Jepang
4 Polypedates megacephalus NC006408 Cina dan India
5 Rana plancyi NC009264 Cina bagian timur
6 Rana nigromaculata NC002805 Rusia, Cina, Korea, Jepang
7 Microhyla ornata NC009422 Asia selatan
8 Kaloula pulchra NC006405 Asia Tenggara, Asia Selatan Cina
9 Limnonectes bannaensis NC012837 -
10 Limnonectes fujianensis NC007440 Cina bagian tengah dan selatan
11 Bufo japonicus NC009886 Endemik Jepang
12 Bufo melanostictus NC005794 Asia Selatan, Cina, Asia Tenggara
termasuk Indonesia
13 Hyla japonica NC010232 Jepang, Cina, Korea
14 Hyla chinensis NC006403 Cina bagian tengah
15 Fejervarya iskandari AB526324
16 Limnonectes hasseltii AB646408
17 Phrynoidis aspera Dalam penelitian ini
18 Polypedates leucomystax Dalam penelitian ini
19 Rhacophorus javanus Dalam penelitian ini
20 Odorana hosii Dalam penelitian ini
21 Limnonectes microdiscus Dalam penelitian ini
22 Fejervarya limnocharis Dalam penelitian ini
20
IV. HASIL
A. Identifikasi Morfologi
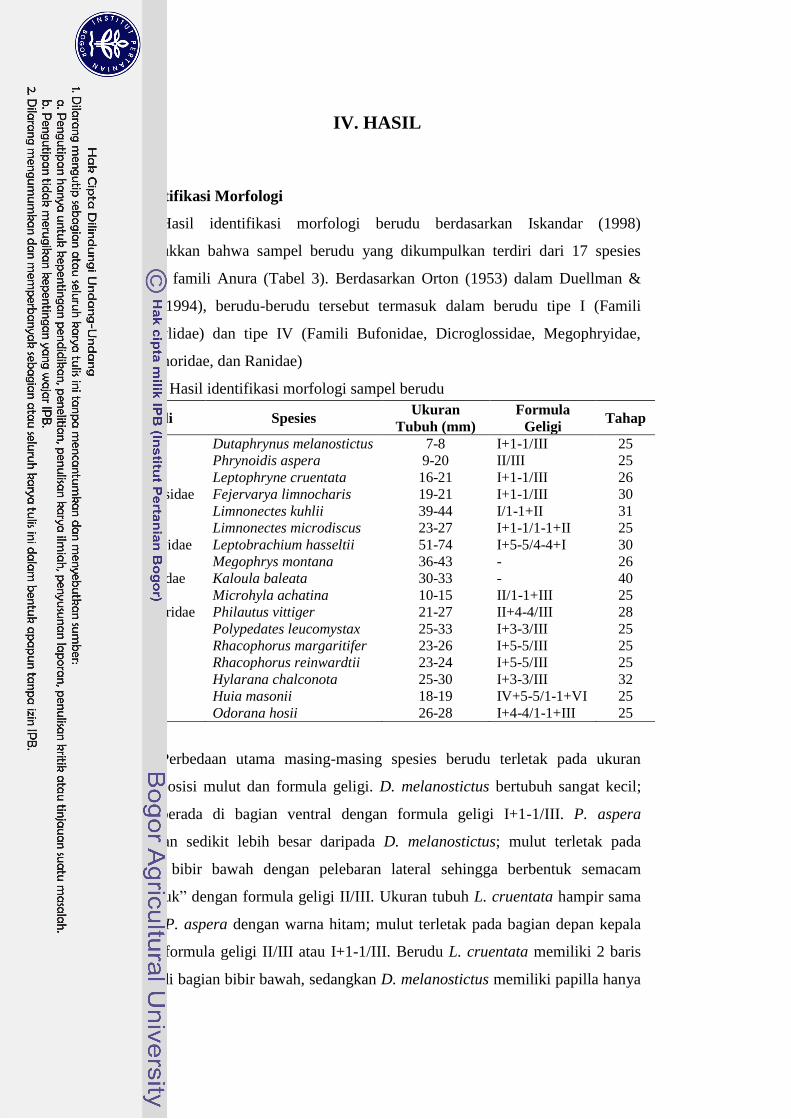
Hasil identifikasi morfologi berudu berdasarkan Iskandar (1998)
menunjukkan bahwa sampel berudu yang dikumpulkan terdiri dari 17 spesies
dalam 6 famili Anura (Tabel 3). Berdasarkan Orton (1953) dalam Duellman &
Trueb (1994), berudu-berudu tersebut termasuk dalam berudu tipe I (Famili
Microhylidae) dan tipe IV (Famili Bufonidae, Dicroglossidae, Megophryidae,
Rhacophoridae, dan Ranidae)
Tabel 3. Hasil identifikasi morfologi sampel berudu
Famili Spesies Ukuran
Tubuh (mm)
Formula
Geligi Tahap
Bufonidae Dutaphrynus melanostictus 7-8 I+1-1/III 25
Phrynoidis aspera 9-20 II/III 25
Leptophryne cruentata 16-21 I+1-1/III 26
Dicroglossidae Fejervarya limnocharis 19-21 I+1-1/III 30
Limnonectes kuhlii 39-44 I/1-1+II 31
Limnonectes microdiscus 23-27 I+1-1/1-1+II 25
Megophryidae Leptobrachium hasseltii 51-74 I+5-5/4-4+I 30
Megophrys montana 36-43 - 26
Microhylidae Kaloula baleata 30-33 - 40
Microhyla achatina 10-15 II/1-1+III 25
Rhacophoridae Philautus vittiger 21-27 II+4-4/III 28
Polypedates leucomystax 25-33 I+3-3/III 25
Rhacophorus margaritifer 23-26 I+5-5/III 25
Rhacophorus reinwardtii 23-24 I+5-5/III 25
Ranidae Hylarana chalconota 25-30 I+3-3/III 32
Huia masonii 18-19 IV+5-5/1-1+VI 25
Odorana hosii 26-28 I+4-4/1-1+III 25
Perbedaan utama masing-masing spesies berudu terletak pada ukuran
tubuh, posisi mulut dan formula geligi. D. melanostictus bertubuh sangat kecil;
mulut berada di bagian ventral dengan formula geligi I+1-1/III. P. aspera
berukuran sedikit lebih besar daripada D. melanostictus; mulut terletak pada
ventral; bibir bawah dengan pelebaran lateral sehingga berbentuk semacam
“mangkuk” dengan formula geligi II/III. Ukuran tubuh L. cruentata hampir sama
dengan P. aspera dengan warna hitam; mulut terletak pada bagian depan kepala
dengan formula geligi II/III atau I+1-1/III. Berudu L. cruentata memiliki 2 baris
papilla di bagian bibir bawah, sedangkan D. melanostictus memiliki papilla hanya

21
pada ujung-ujung bibir saja. Hal ini yang membedakan jenis berudu L. cruentata
dengan berudu D. melanostictus.
Tubuh F. limnocharis berukuran sedang; mulut berada di bagian ventral
kepala dengan formula geligi I+1-1/III. Deret gigi terakhir pada spesies ini lebih
pendek dari dua deret gigi lainnya. Ukuran tubuh L. kuhlii dua kali lebih besar
daripada ukuran tubuh L. microdiscus. Mulut L. kuhlii dan L. microdiscus berada
pada ujung kepala dengan formula geligi masing-masing adalah I/1-1+II dan I+1-
1/1-1+II.
Ukuran tubuh berudu famili Megophryidae sangat besar. Namun, ukuran
tubuh berudu L. hasseltii 1,5 sampai 2 kali lebih besar daripada berudu M.
montana. Mulut berudu L. hasseltii kecil, berada di bagian ujung ventral kepala
dengan formula geligi I+5-5/4-4+5. Mulut berudu M. montana berada pada ujung
kepala dan memiliki ciri khusus yaitu dengan perpanjangan lateral sehingga
berbetuk seperti “kumis”. Berudu ini tidak memiliki formula geligi tertentu.
Ukuran tubuh berudu K. baleata sedang. Berudu ini tidak memiliki bentuk
mulut seperti berudu pada umumnya, hanya berupa sebuah lubang kecil pada
ujung kepalanya. Berudu M. achatina berukuran sangat kecil; mulut menghadap
ke bawah; bibir bawah melebar dengan formula geligi II/1-1+III.
Berudu P. vittiger berukuran sedang, mulut berada pada bagian ventral
kepala dengan formula geligi II+4-4/III. Berudu P. leucomystax, R. margaritifer
dan R. reinwardtii berukuran sedang sampai besar. Seluruh berudu tersebut
memiliki mulut yang terletak di bagian ventral kepala, menghadap ke bawah.
Formula geligi masing-masing berudu berturut-turut adalah I+3-3/III, I+5-5/III
atau II+4-4/III dan I+5-5/III. Berudu R. margaritifer dan R. reinwardtii dibedakan
berdasarkan jumlah papilla marjinal di bibir bagian bawah, di mana R.
margaritifer memiliki 2 baris papilla dan R. Reinwardtii memiliki 1 baris papilla.
Berudu H. masonii, H. chalconota dan O. hosii berukuran sedang. Ukuran
tubuh H. masonii lebih besar dari ukuran tubuh kedua spesies lainnya. Berudu
jenis ini memiliki ciri khas yaitu pada bagian ventral terdapat bentuk seperti
mangkuk menghadap ke bawah. Mulut ketiga spesies berudu tersebut terletak
pada bagian ventral tubuh dan menghadap ke bawah, dengan formula geligi
masing-masing adalah IV+5-5/1-1+VI, I+3-3/III dan I+4-4/1-1+III.
22
B. Amplifikasi dan Visualisasi DNA
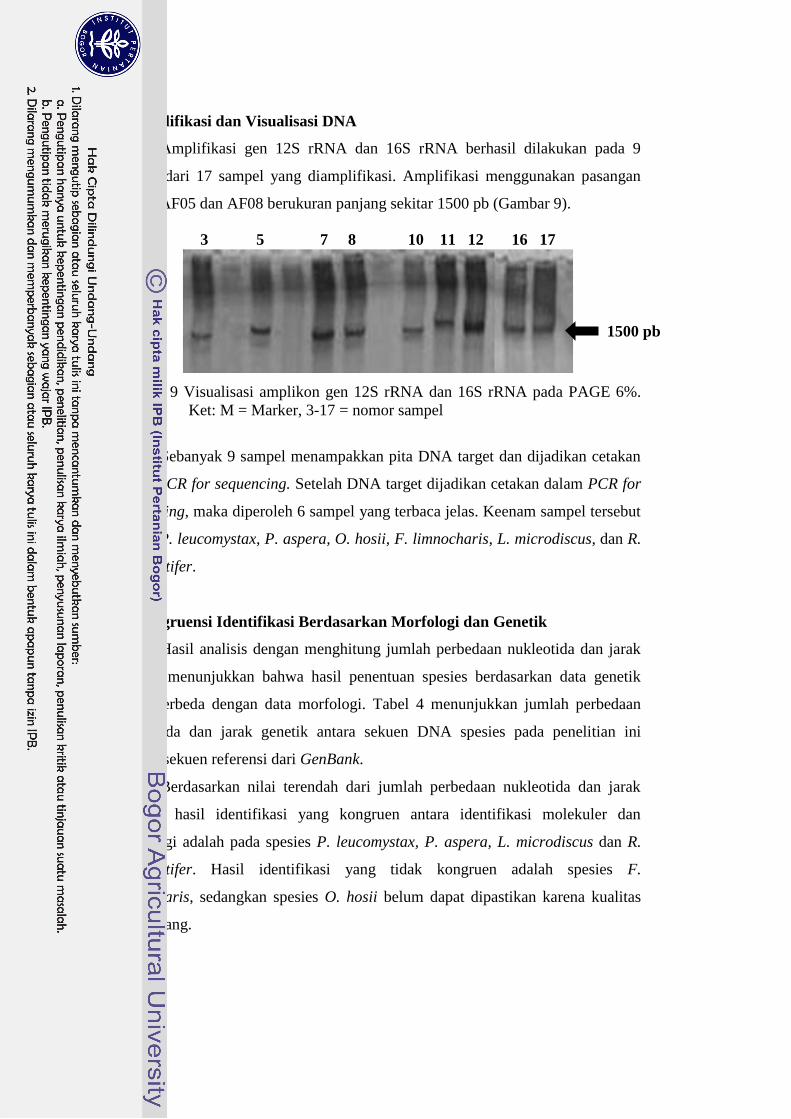
Amplifikasi gen 12S rRNA dan 16S rRNA berhasil dilakukan pada 9
sampel dari 17 sampel yang diamplifikasi. Amplifikasi menggunakan pasangan
primer AF05 dan AF08 berukuran panjang sekitar 1500 pb (Gambar 9).
Gambar 9 Visualisasi amplikon gen 12S rRNA dan 16S rRNA pada PAGE 6%.
Ket: M = Marker, 3-17 = nomor sampel
Sebanyak 9 sampel menampakkan pita DNA target dan dijadikan cetakan
dalam PCR for sequencing. Setelah DNA target dijadikan cetakan dalam PCR for
sequencing, maka diperoleh 6 sampel yang terbaca jelas. Keenam sampel tersebut
adalah P. leucomystax, P. aspera, O. hosii, F. limnocharis, L. microdiscus, dan R.
margaritifer.
C. Kongruensi Identifikasi Berdasarkan Morfologi dan Genetik
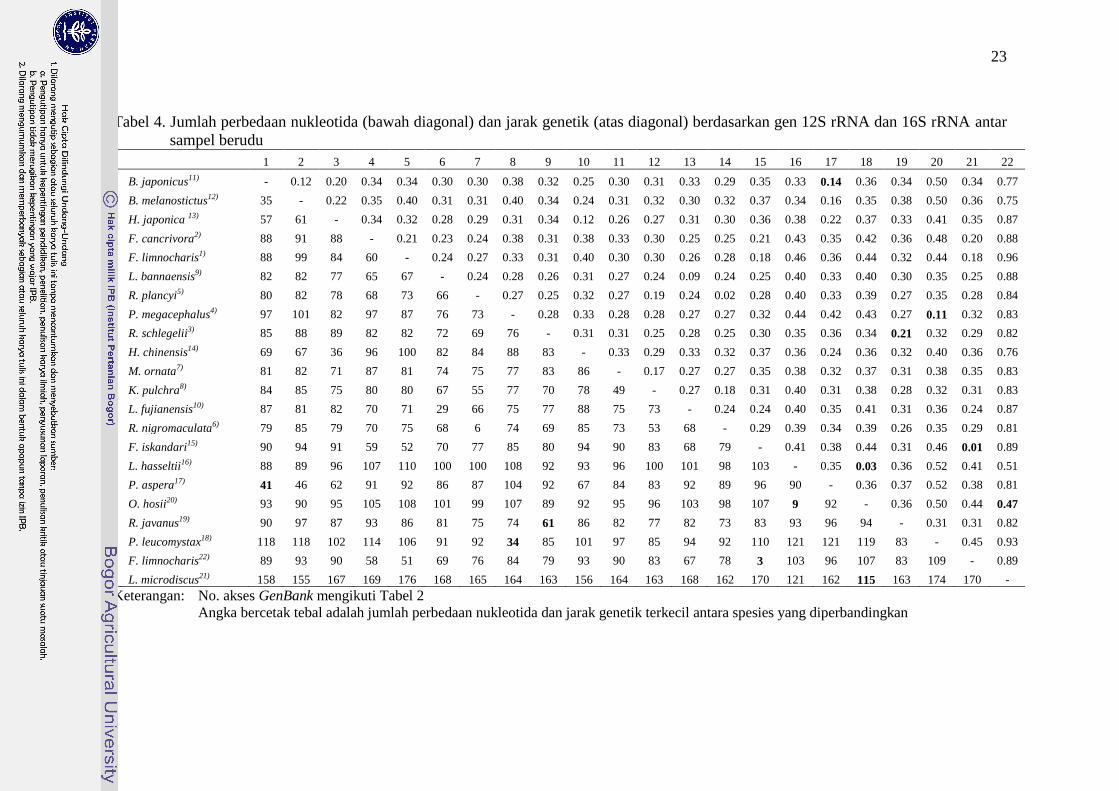
Hasil analisis dengan menghitung jumlah perbedaan nukleotida dan jarak
genetik menunjukkan bahwa hasil penentuan spesies berdasarkan data genetik
dapat berbeda dengan data morfologi. Tabel 4 menunjukkan jumlah perbedaan
nukleotida dan jarak genetik antara sekuen DNA spesies pada penelitian ini
dengan sekuen referensi dari GenBank.
Berdasarkan nilai terendah dari jumlah perbedaan nukleotida dan jarak
genetik, hasil identifikasi yang kongruen antara identifikasi molekuler dan
morfologi adalah pada spesies P. leucomystax, P. aspera, L. microdiscus dan R.
margaritifer. Hasil identifikasi yang tidak kongruen adalah spesies F.
limnocharis, sedangkan spesies O. hosii belum dapat dipastikan karena kualitas
data kurang.
1500 pb
3 5 7 8 10 11 12 16 17
23
Tabel 4. Jumlah perbedaan nukleotida (bawah diagonal) dan jarak genetik (atas diagonal) berdasarkan gen 12S rRNA dan 16S rRNA antar
sampel berudu
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
1 B. japonicus11) - 0.12 0.20 0.34 0.34 0.30 0.30 0.38 0.32 0.25 0.30 0.31 0.33 0.29 0.35 0.33 0.14 0.36 0.34 0.50 0.34 0.77
2 B. melanostictus12) 35 - 0.22 0.35 0.40 0.31 0.31 0.40 0.34 0.24 0.31 0.32 0.30 0.32 0.37 0.34 0.16 0.35 0.38 0.50 0.36 0.75
3 H. japonica 13) 57 61 - 0.34 0.32 0.28 0.29 0.31 0.34 0.12 0.26 0.27 0.31 0.30 0.36 0.38 0.22 0.37 0.33 0.41 0.35 0.87
4 F. cancrivora2) 88 91 88 - 0.21 0.23 0.24 0.38 0.31 0.38 0.33 0.30 0.25 0.25 0.21 0.43 0.35 0.42 0.36 0.48 0.20 0.88
5 F. limnocharis1) 88 99 84 60 - 0.24 0.27 0.33 0.31 0.40 0.30 0.30 0.26 0.28 0.18 0.46 0.36 0.44 0.32 0.44 0.18 0.96
6 L. bannaensis9) 82 82 77 65 67 - 0.24 0.28 0.26 0.31 0.27 0.24 0.09 0.24 0.25 0.40 0.33 0.40 0.30 0.35 0.25 0.88
7 R. plancyi5) 80 82 78 68 73 66 - 0.27 0.25 0.32 0.27 0.19 0.24 0.02 0.28 0.40 0.33 0.39 0.27 0.35 0.28 0.84
8 P. megacephalus4) 97 101 82 97 87 76 73 - 0.28 0.33 0.28 0.28 0.27 0.27 0.32 0.44 0.42 0.43 0.27 0.11 0.32 0.83
9 R. schlegelii3) 85 88 89 82 82 72 69 76 - 0.31 0.31 0.25 0.28 0.25 0.30 0.35 0.36 0.34 0.21 0.32 0.29 0.82
10 H. chinensis14) 69 67 36 96 100 82 84 88 83 - 0.33 0.29 0.33 0.32 0.37 0.36 0.24 0.36 0.32 0.40 0.36 0.76
11 M. ornata7) 81 82 71 87 81 74 75 77 83 86 - 0.17 0.27 0.27 0.35 0.38 0.32 0.37 0.31 0.38 0.35 0.83
12 K. pulchra8) 84 85 75 80 80 67 55 77 70 78 49 - 0.27 0.18 0.31 0.40 0.31 0.38 0.28 0.32 0.31 0.83
13 L. fujianensis10) 87 81 82 70 71 29 66 75 77 88 75 73 - 0.24 0.24 0.40 0.35 0.41 0.31 0.36 0.24 0.87
14 R. nigromaculata6) 79 85 79 70 75 68 6 74 69 85 73 53 68 - 0.29 0.39 0.34 0.39 0.26 0.35 0.29 0.81
15 F. iskandari15) 90 94 91 59 52 70 77 85 80 94 90 83 68 79 - 0.41 0.38 0.44 0.31 0.46 0.01 0.89
16 L. hasseltii16) 88 89 96 107 110 100 100 108 92 93 96 100 101 98 103 - 0.35 0.03 0.36 0.52 0.41 0.51
17 P. aspera17) 41 46 62 91 92 86 87 104 92 67 84 83 92 89 96 90 - 0.36 0.37 0.52 0.38 0.81
18 O. hosii20) 93 90 95 105 108 101 99 107 89 92 95 96 103 98 107 9 92 - 0.36 0.50 0.44 0.47
19 R. javanus19) 90 97 87 93 86 81 75 74 61 86 82 77 82 73 83 93 96 94 - 0.31 0.31 0.82
20 P. leucomystax18) 118 118 102 114 106 91 92 34 85 101 97 85 94 92 110 121 121 119 83 - 0.45 0.93
21 F. limnocharis22) 89 93 90 58 51 69 76 84 79 93 90 83 67 78 3 103 96 107 83 109 - 0.89
22 L. microdiscus21) 158 155 167 169 176 168 165 164 163 156 164 163 168 162 170 121 162 115 163 174 170 -
Keterangan: No. akses GenBank mengikuti Tabel 2
Angka bercetak tebal adalah jumlah perbedaan nukleotida dan jarak genetik terkecil antara spesies yang diperbandingkan
24

24
D. Rekonstruksi Filogeni
Urutan basa nukleotida yang analisis berjumlah 1362 nt, terdiri dari 504 nt
gen 12S rRNA dan 858 nt gen 16S rRNA. Pada rekonstruksi filogeni berdasarkan
gen 12S rRNA, spesies P. leucomystax tidak disertakan dalam analisis karena
jumlah nukleotida spesies tersebut dalam penelitian ini sangat pendek (< 20 nt).
Tabel 5 menunjukkan nilai konsistensi dan kestabilan pohon filogeni yang
direkonstruksi berdasarkan masing-masing gen dan gabungan gen 12S dan 16S
rRNA (selanjutnya disimbolkan dengan 12S-16S rRNA). Berdasarkan tabel
tersebut, nilai CI dan RI hasil rekonstruksi filogeni berdasarkan gen 16S rRNA
tertinggi di antara dua rekonstruksi lainnya. Hal ini menujukkan bahwa pohon
filogeni berdasarkan gen 16S rRNA merupakan pohon filogeni yang paling
konsisten dan stabil.
Tabel 5. Nilai CI (Consistency Index), RI (Retention index), RC (Rescaled
consistency index) ketiga pohon filogeni Anura
Gen CI RI RC
12S rRNA 0.4540 0.5702 0.2589
16S rRNA 0.5169 0.6103 0.3155
12S-16S rRNA 0.4786 0.5667 0.2713
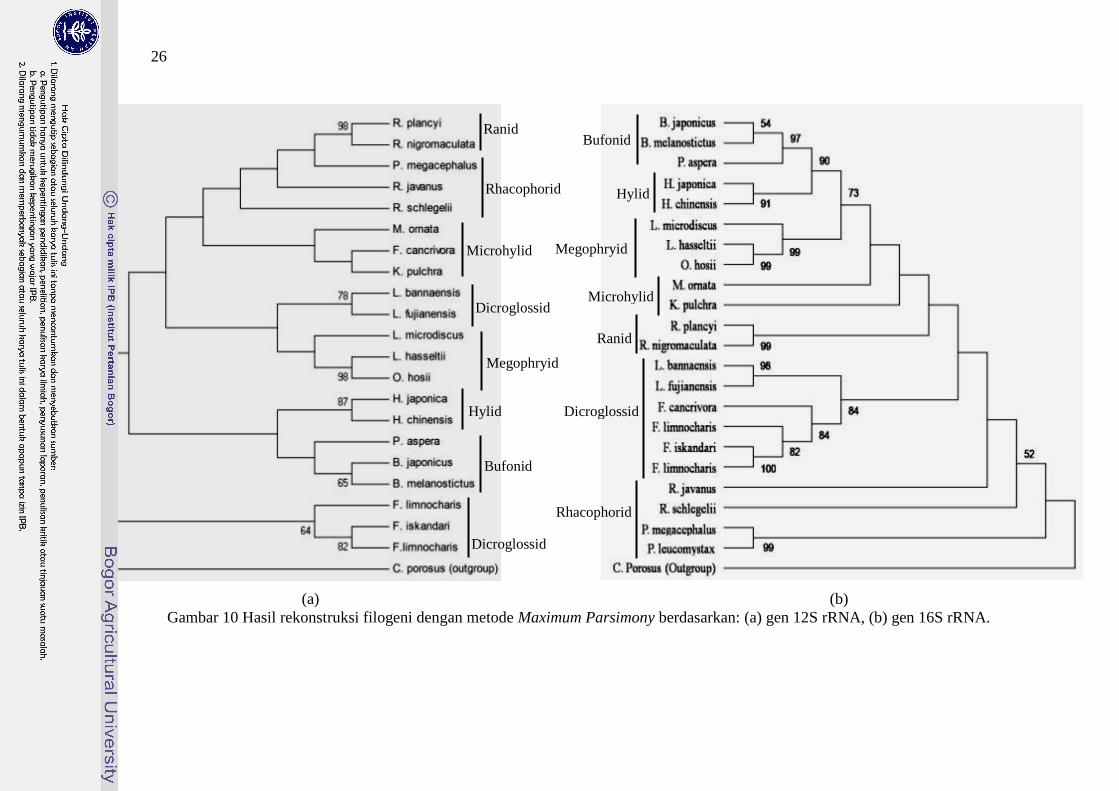
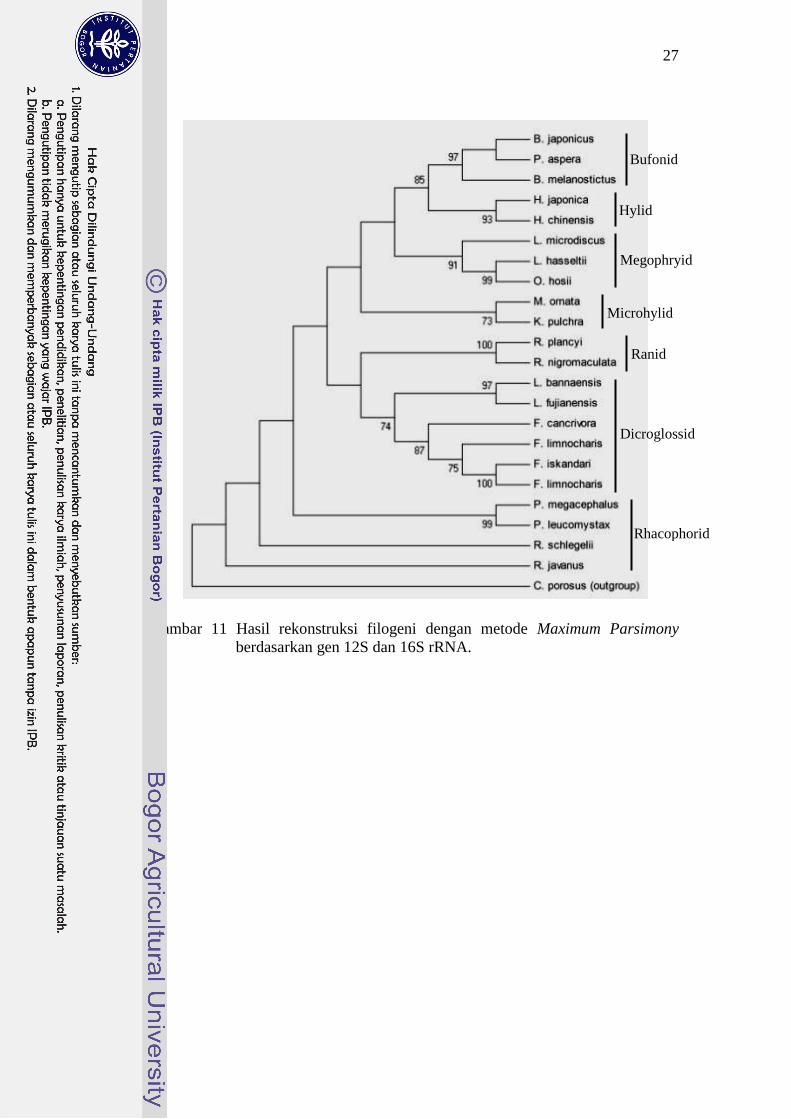
Hasil rekonstruksi filogeni menghasilkan topologi yang yang berbeda pada
gen 12S rRNA (Gambar 10(a)). Topologi pohon berdasarkan gen 16S rRNA
(Gambar 10(b)) sama dengan topologi pohon berdasarkan gen 12S-16S rRNA
(Gambar 11), di mana posisi basal ditempati oleh kelompok Rhacophorid dan
posisi terluar ditempati oleh kelompok Bufonid. Posisi basal topologi pohon
filogeni gen 12S rRNA ditempati oleh kelompok Dicroglossid dan posisi terluar
ditempati oleh kelompok Ranid.
Kesamaan dari ketiga pohon filogeni tersebut adalah Bufonid berkelompok
dengan Hylid dalam satu kelompok monofiletik. Kelompok-kelompok
monofiletik lainnya antara lain Mycrohylid dan Ranid. Dicroglossid merupakan
kelompok monofiletik pada topologi pohon gen 16S rRNA dan gen 12S-16S
rRNA sedangkan berdasarkan gen 12S rRNA kelompok tersebut terpisah pada 2
kelompok, yaitu kelompok genus Fejervarya dan kelompok genus Limnonectes.
Pada ketiga topologi pohon filogeni, kelompok Rhacophorid terpisah secara

25
parafiletik. Genus Polypedates membentuk cabang monofiletik dan berhubungan
dekat dengan Rhacophorus.
Pada ketiga rekonstruksi filogeni, spesies L. microdiscus, L. hasseltii dan
O. hosii (selanjutnya disebut kelompok Megophryid) membentuk kelompok
monofiletik dengan dukungan bootstrap >90%, sedangkan spesies F. limnocharis
menempati posisi lebih dekat dengan F. iskandari pada kelompok Dicroglossid
dengan dukungan bootstrap 100%. Spesies P. aspera menempati posisi lebih
dekat dengan genus Bufo berdasarkan gen 12S rRNA dan gen 16S rRNA,
sedangkan berdasarkan gen 12S-16S rRNA spesies ini menempati posisi lebih
dekat dengan spesies B. japonicus dengan dukungan bootstrap rendah (<50%).
Posisi basal kelompok Rhacophorid berbeda pada ketiga topologi pohon.
Spesies R. schlegelii menempati posisi basal Rhacophorid berdasarkan gen 12S
rRNA, spesies anggota genus Polypedates menempati posisi basal berdasarkan
gen 16S rRNA, sedangkan berdasarkan gen 12S-16S rRNA posisi basal
Rhacophorid ditempati oleh R. javanus.
26
(a) (b)
Gambar 10 Hasil rekonstruksi filogeni dengan metode Maximum Parsimony berdasarkan: (a) gen 12S rRNA, (b) gen 16S rRNA.
Bufonid Ranid
Rhacophorid Hylid
Microhylid
Dicroglossid
Ranid
Bufonid
Dicroglossid
Microhylid
Hylid
Rhacophorid
Dicroglossid
Megophryid
Megophryid
27
Gambar 11 Hasil rekonstruksi filogeni dengan metode Maximum Parsimony
berdasarkan gen 12S dan 16S rRNA.
Bufonid
Hylid
Microhylid
Ranid
Dicroglossid
Rhacophorid
Megophryid

28
V. PEMBAHASAN
A. Kongruensi Identifikasi Morfologi dan Molekuler
Ukuran tubuh sampel berudu O. hosii dan L. microdiscus sekitar 1-2 kali
lebih kecil daripada berudu L. hasseltii pada tahap perkembangan yang sama.
Perbedaan ukuran tubuh tersebut diduga sebagai akibat dari perbedaan peletakan
telur oleh induk betina. Pada kondisi yang tidak memungkinkan untuk meletakkan
telur di genangan air di lantai hutan, katak betina L. hasseltii dapat meletakkan
telur pada tempat lain seperti di tepi sungai berarus lambat atau tenang. Fenomena
seperti ini juga terlihat pada jenis katak pohon (Kusrini et al. 2009). Berudu yang
hidup pada kolam temporer, misal genangan air, akan lebih cepat tumbuh dan
bermetamorfosis dibandingkan dengan berudu yang hidup pada kolam permanen,
misal sungai. Hal ini dikarenakan kolam temporer memiliki kelebihan dalam hal
potensi pakan (Wassersug 1975; Wilbur 1987) dengan lebih sedikit jumlah
predator (Brendonck et al. 2002; Petranka & Kennedy 1999). Sifat sementara dari
kolam menguntungkan dari aspek tersebut namun mengharuskan berudu untuk
bermetamorfosis pada waktu tertentu.
Formula geligi O. hosii berbeda satu baris pada geligi atas dengan L.
hasseltii. Iskandar (1998) menyebutkan bahwa pada berudu terdapat variasi
formula geligi dalam satu spesies yang sama. Formula geligi berudu sempurna
pada awal fase larva, namun proporsi relatif formula geligi dan oral disc terus
berubah selama metamorfosis. Pada berudu Scaphiopus dan kelompok Ranid
jumlah baris geligi bertambah selama metamorfosis (Gosner 1960).
Genus Philautus merupakan salah satu Anura yang seluruh fase
metamorfosisnya terjadi di dalam telur sehingga pada akhir metamorfosis telur
menetaskan katak muda (Inger 1996). Metamorfosis pada spesies P. vittiger
diketahui memiliki berudu yang hidup bebas di dalam air (Kusrini et al. 2008),
sehingga pengelompokkan spesies ini perlu dipelajari lebih lanjut untuk
memastikan posisinya dalam taksonomi Anura.
Pada awal perkembangannya, data molekuler hanya digunakan oleh para
taksonomis sebagai alat untuk mempelajari kekerabatan antar taksa dan proses
evolusi sebuah kelompok organisme. Namun, saat ini data molekuler telah

29
berkembang penggunaannya, salah satunya dalam proses penentuan spesies oleh
para ekologis untuk mempelajari keanekaragaman hayati suatu wilayah tertentu.
Data molekuler dapat dengan cepat dan akurat dalam penentuan spesies, terutama
pada spesies kriptik, spesies sibling dan spesies dengan variasi morfologi akibat
pengaruh lingkungan. Sumber data molekuler tidak hanya dapat diperoleh dari
darah atau jaringan otot saja, melainkan juga dari rambut, feses atau urin. Hal ini
merupakan keuntungan lain dari data molekuler terutama pada kegiatan penelitian
dengan target hewan yang jarang dan/atau sulit ditemukan secara langsung. Data
molekuler merupakan data dasar yang tidak subjektif dan dapat dianalisis kembali.
Data molekuler mempunyai beberapa tantangan dalam penerapannya.
Pertama, memerlukan keahlian khusus terutama pada teknik laboratorium
molekuler, di mana tidak semua peneliti mampu melakukannya. Kedua, biaya
relatif mahal bagi negara berkembang seperti Indonesia, tidak hanya peralatan
mesin PCR dan sekuensing tetapi juga bahan kimia dan komponennya. Ketiga,
jumlah basa minimal yang digunakan pada saat identifikasi spesies (>500 pb) sulit
dicapai pada sampel bangkai atau DNA terdegradasi.
Hasil yang diperoleh dari data molekuler masih lebih besar bila
dibandingkan dengan usaha yang dikeluarkan jika identifikasi spesies dilakukan
dengan pendekatan ini. Selain identifikasi spesies, data molekuler juga dapat
digunakan untuk mengetahui biodiversitas masa lampau (Paleoecology),
mempelajari interaksi inter-spesies, mempelajari evolusi dan kekerabatan antar
taksa, serta mengetahui jenis pakan satwa. Karakter morfologi kadang sudah tidak
mampu lagi untuk menggambarkan proses evolusi suatu organisme karena
karakter yang sangat kompleks. Selain itu, sering terjadi tentangan pada hubungan
filogeni antara taksa berdasarkan karakter morfologi karena perbedaan
pembobotan karakter dalam analisis. Alasan-alasan inilah yang menjadi dasar
mengapa pendekatan molekuler perlu digunakan sebagai pelengkap dan penguat
data morfologi.
B. Filogenetik
Dukungan nilai bootstrap yang rendah, ketidakstabilan topologi dapat
terjadi karena gen yang digunakan dalam analisis hanya satu gen saja dan

30
berukuran lebih pendek (parsial). Cao et al. (1994) telah menunjukkan bahwa set
data gen yang besar, seperti serangkaian gen-gen yang berurutan memberikan
hasil analisis yang lebih kuat dibandingkan dengan analisis yang didasarkan pada
gen tunggal.
Hasil analisis molekular sebelumnya mendukung bahwa kelompok
Rhacophorid merupakan kelompok monofiletik (Li et al. 2008; Yu et al. 2009).
Namun berdasarkan data morfologi, Channing (1989) menyatakan bahwa
Rhacophorinae (Rhacophorus dan Polypedates) merupakan kelompok yang
polifiletik. Hubungan yang dekat antara Rhacophorus dan Polypedates didukung
dengan baik pada penelitian ini dan penelitian sebelumnya baik berdasarkan
morfologi (Wilkinson dan Drewes 2000) maupun molekular (Wilkinson et al.
2002; Frost et al. 2006). Dubois (1986) menggabungkan genus Polypedates ke
dalam genus Rhacophorus karena menganggap karakter morfologi kedua genus
tersebut tidak cukup berbeda untuk dipisahkan. Sedangkan Duellman (1993) dan
Frost et al. (2000) memisahkan kedua genus tersebut berdasarkan karakter
morfologi yang dipakai oleh Channing (1989) karena Rhacophorus dan
Polypedates bukan merupakan sister group. Berdasarkan uraian tersebut, masih
diperlukan penelitian lebih lanjut untuk memastikan hubungan kekerabatan antara
kedua genus tersebut dalam kelompok Rhacophorid.
Rekonstruksi filogeni berdasarkan morfologi menunjukkan bahwa Ranid
dan Dicroglossid dapat membentuk cabang parafiletik (Ford and Cannatella,
1993). Hasil tersebut juga didukung oleh Haas (2003) dengan menggunakan
seluruh karakter morfologi larva Anura. Namun, hasil rekonstruksi filogeni
dengan mengeluarkan 4 karakter morfometri dari analisis menghasilkan
percabangan monofiletik pada kedua taksa tersebut. Optimasi karakter yang
dilakukan tersebut menunjukkan bahwa terdapat 3 karakter synapomorfi pada
Ranidae, yaitu firmisterny, adanya basihyal dan fenestrae parietals. Hubungan
monofiletik kelompok ini didukung dengan kuat berdasarkan DNA mitokondria
(Frost et al. 2006). Kelompok Dicroglossid merupakan kelompok monofiletik
didukung dengan baik berdasarkan data morfologi (Anderson 1871; Emerson and
Berrigan 1993) dan data DNA mitokondria (Emerson et al. 2000; Evans et al.
2003; Frost et al. 2006).

31
Fejervarya limnocharis merupakan spesies dengan penyebaran yang
sangat luas. Morfologi antar populasi spesies tersebut terlalu sulit untuk
dibedakan karena variasi morfologinya kurang dikenali dan tidak adanya diagnosa
spesifik antara spesies pada populasi lokal dan spesies pada populasi lainnya
sehingga seluruh spesies ini dinamai dengan nama yang sama, yaitu F.
limnocharis (Dubois, 1984, 1987; Inger and Voris, 2001). Veith et al. (2001)
berhasil mendeskripsikan satu spesies kriptik yang dideteksi oleh Toda et al.
(1998) sebagai spesies baru, yaitu F. iskandari, berdasarkan data sekuen
Allozyme dan DNA mitokondria. Data molekuler tersebut menunjukkan bahwa
tidak ada aliran gen antara F. iskandari dan F. limnocharis.
Fejervarya iskandari dideskripsi pertama kali di Bandung dan Sukabumi,
Jawa Barat namun diperkirakan spesies ini menyebar lebih luas di Pulau Jawa
(Iskandar dan Mumpuni 2004) sehingga sangat mungkin jika sampel yang
digunakan dalam penelitian ini adalah berudu F. iskandari.
Bufonid merupakan kelompok monofiletik didukung dengan baik oleh
bootstrap pada penelitian ini. Hubungan filogenetik tersebut juga dihasilkan oleh
Liu et al. (2000) dan Pramuk et al. (2001) berdasarkan data sekuen DNA
mitokondria dan Haas (2003) berdasarkan data morfologi larva. Posisi spesies P.
aspera pada topologi pohon filogeni berdasarkan gen 12S rRNA dan 16S rRNA
berbeda dengan posisi spesies tersebut pada topologi pohon berdasarkan gen 12S-
16S rRNA. Phrynoidis aspera dipindahkan dari sinonimnya, yaitu Bufo asper
berdasarkan data DNA inti dan DNA mitokondria (Frost et al. 2006). Hasil
penelitian tersebut menunjukkan bahwa B. asper berhubungan lebih dekat dengan
genus Pedostibes dibandingkan dengan spesies Bufo lainnya. Frost et al. (2006)
memasukkan B. asper ke dalam genus Phrynoidis juga berdasarkan pada karakter
sinapomorfi yang dimiliki oleh genus Phrynoidis yang menguatkan percabangan
kelompok B. asper.

32
VI. KESIMPULAN DAN SARAN
A. Kesimpulan
Identifikasi berudu berdasarkan morfologi dapat dilakukan berdasarkan
perbedaan bentuk mulut dan formula geligi. Penentuan spesies dengan
menggunakan karakter morfologi dapat berbeda dari penentuan spesies dengan
menggunakan data sekuen DNA. Berdasarkan karakter molekuler, spesies O. hosii
memiliki kemiripan yang tinggi dengan L. hasseltii, dan spesies F. limnocharis
memiliki kemiripan yang tinggi dengan F. iskandari. Untuk spesies yang
menunjukkan kongruensi identifikasi morfologi dan molekuler, maka identifikasi
di lapang cukup dilakukan dengan karakter morfologi, seperti jenis P.
leucomystax, P. aspera, L. microdiscus dan R. margaritifer.
Hasil analisis filogenetik menunjukkan bahwa sebagian besar taksa Anura
membentuk kelompok monofiletik kecuali pada kelompok Rhacophorid
berdasarkan gen 12S rRNA.
B. Saran
Penggunaan karakter ukuran tubuh sebaiknya tidak digunakan dalam
identifikasi spesies karena sangat dipengaruhi oleh variasi lingkungan.
Penggunaan gen tunggal, yaitu 16S rRNA dapat dilakukan dalam analisis
filogenetik Anura untuk memperpendek data sekuen yang dianalisis serta
meningkatkan peluang keberhasilan dalam proses ampllifikasi.

33
DAFTAR PUSTAKA
Anderson J. 1871. A list of the reptilian accession to the Indian Museum, Calcutta
from 1865 to 1870, with a description of some new species. Journal of the
Asiatic Society of Bengal 40: 12-39.
Altig R. 2007. A Primer for the Morphology of Anuran Tadpoles. Herpetological
Conservation and Biology 2(1):71-74.
Brendonck L, Michels E, DeMeester L, Riddoch B. 2002. Temporary ponds are
not “enemy free”. Hydrobiologica 486: 147-159
Cao Y, Adachi A, Janke A, Paabo S, Hasegawa M. 1994. Phylogenetic
relationships among eutherian orders estimated from inferred protein
sequences of mithocondrial proteins: instability of a tree based on single
gene. J Mol Evol 39: 519-527
Che J, Pang J, Zhao H, Wu G, Zhao E, Zhang Y. 2007. Phylogeny of Raninae
(Anura: Ranidae) inferred from mitochondrial and nuclear sequences.
Molecular Phylogenetics and Evolution 43: 1–13
Cogger HG, Zweifel RG. 2003. Encyclopedia of Reptiles and Amphibians Second
Edition. San Fransisco: Fog City Press
Dubois A. 1984. Note preliminaire sur le groupe de Rana limnocharis
Gravenhorst, 1929 (Amphibiens, Anaures). Alytes 3: 143–159
Dubois A .1987. Miscellanea taxomica batrachologica (1). Alytes 5: 7–95
Duellman WE, Trueb L. 1994. Biology of Amphibians. New York: McGraw-Hill.
Emerson SB, Inger RF, Iskandar DT. 2000. Molecular systematics and
biogeography of the fanged frogs of Southeast Asia. Molecular
Phylogenetics and Evolution 16(1): 131–142
Emerson SB, Berrigan D. 1993. Systematics of Southeast Asia ranids: Multiple
origin of voicelessness in the subgenus Limnonectes (Fitzinger).
Herpetologica 49(1): 22-31.
Evans BJ, Brown RF, McGuire JA, Supriatna J, Andayani N, Diesmos A,
Iskandar DT, Melnick DJ, Cannatella DC. 2003. Phylogenetics of Fanged
Frogs: Testing Biogeographical Hypotheses at the Interface of the Asian and
Australian Faunal Zones. Syst. Biol. 52(6): 794–819.
Farajallah A. 2002. Karakterisasi genom mitokondria labi-labi Dogania subplana
(Trionychidae, Testudines, Reptilia) [Disertasi]. Bogor. Program
Pascasarjana, Institut Pertanian Bogor.

34
Frost, DR, Grant T, Faivovich J, Bain RH, Haas A, Haddad CFB, De Sá RO,
Channing A, Wilkinson M, Donnellan SC, Raxworthy CJ, Campbell JA,
Blotto BL, Moler P, Drewes RC, Nussbaum RA, Lynch JD, Green DM,
Wheeler WC, 2006. The amphibian tree of life. Bull. Am. Mus. Nat. Hist.
297: 1–370
Ford LS, Cannatella DC. 1993. The major clades of frogs. Herpetological
monographs 7: 94-117
Gosner LK. 1960. A Simplified Table for Staging Anuran Embryos and Larvae
with Notes on Identification. Herpetologica 16(3): 183-190
Gosner LK, Rossman DA. 1960. Eggs and larval development of the tree frogs
Hyla crucifer and Hyla ocularis. Herpetologica 16(4): 225-232.
Haas A. 2003. Phylogeny of frogs as inferred from primarily larval characters
(Amphibia: Anura). Cladistics 19: 23–89.
Hamidy A, Matsui M, Shimada T, Nishikawa K, Yambun P, Sudin A, Kusrini
MD, Kurniati H. 2011. Morphological and genetic discordance in two
species of Bornean Leptobrachium (Amphibia, Anura, Megophryidae).
Molecular Phylogenetics and Evolution 61: 904–913.
Hillis DM, Wilcox TP. 2005. Phylogeny of the New World true frogs (Rana).
Molecular Phylogenetics and Evolution 34: 299–314.
Hofrichter R. 1999. The Encyclopedia of Amphibian. Canada: Key Porter Books
Limited.
Inger RF. 1966. The systematic and zoogeography of the amphibian of Borneo.
Fieldiana: Zoology Volume 52. Field Museum of Natural History. United
States of America: Field Museum Press
Inger RF. 1972. Bufo of Eurasia. Di dalam W.F. Blair (editor), Evolution in the
genus Bufo: 102–118. Austin: University of Texas Press.
Inger RF, Voris HK. 2001. Biogeographical relations of the frog and snake of
Sundaland. J Biogeogr 28: 863–891
Iskandar DT. 1998. Amfibi Jawa dan Bali – Seri Panduan Lapangan. Bogor:
Puslitbang – LIPI.
Iskandar DT, Mumpuni 2004. Fejervarya iskandari. In: IUCN 2011. IUCN Red
List of Threatened Species. Version 2011.2. <www.iucnredlist.org>.
[13 May 2012].
Kitching IJ, Forey PL, Humphries CJ, Williams DM. 1998. Cladistics second
edition: The Theory and Practice of Parsimony Analysis. New York: Oxford
University Press Inc

35
Kusrini MD, Lubis ML, Darmawan B. 2008. The Tree Frog of Chevron
Geothermal Concession, Mount Halimun Salak National Park Indonesia.
Technical report submitted to the Wildlife Trust – Peka Foundation.
Kusrini MD, Lubis MI, Darmawan B, Rahman NL, Hypananda W. 2009. Final
Report: Integrated Ecological Research of Conservation of the Tree Frog in
West Java, Indonesia. Technical report submitted to the Wildlife Truts –
Peka Foundation.
Leong TM, Chou LM. 2000. Tadpole of the Celebes Toad Bufo Celebensis
Gunther (Amphibia: Anura: Bufonidae) from Northeast Sulawesi. The
Raffles Bulletin of Zoology 48(2): 297-30.
Li J, Che J, Bain RH, Zhao E, Zhang Y. 2008. Molecular phylogeny of
Rhacophoridae (Anura): A framework of taxonomic reassignment of species
within the genera Aquixalus, Chiromantis, Rhacophorus, and Philautus.
Molecular Phylogenetics and Evolution 48: 302–312.
Lipscomb D. 1998. Basics of cladistic analysis. Washington DC: George
Washington Univ.
Liu W, Lathrop A, Fu J, Yang D, Murphy RW. 2000. Phylogeny of East Asian
Bufonids Inferred from Mitochondrial DNA Sequences (Anura: Amphibia).
Molecular Phylogenetics and Evolution 14(3): 423–435.
Matsui M, Hamidy A, Murphy RW, Khonsue W, Yambun P, Shimada T, Ahmad
N, Belabut DM, Jiang JP. 2010. Phylogenetic relationships of megophryid
frogs of the genus Leptobrachium (Amphibia, Anura) as revealed by
mtDNA gene sequences. Molecular Phylogenetics and Evolution 56: 259–
272.
McLeod DS. 2010. Of Least Concern? Systematics of a cryptic species complex:
Limnonectes kuhlii (Amphibia: Anura: Dicroglossidae). Molecular
Phylogenetics and Evolution 56: 991–1000.
Mistar. 2003. Panduan Lapang Amfibi Kawasan Ekosistem Leuser. Bogor:
PILINGO Movement.
Ningsih WD. 2011. Struktur komunitas berudu anura di Sungai Cibeureum
Taman Nasional Gunung Gede Pangrango, Jawa Barat [Skripsi]
Departemen Konservasi Sumberdaya Hutan Dan Ekowisata Fakultas
Kehutanan IPB. Bogor. Tidak dipublikasi
Page RDM, Holmes EC. 1998. Molecular Evolution: A Phylogenetic Approach.
United Kingdom: Blackwell Science.
Petranka JW, Kennedy CA. 1999. Pond tadpoles with generalised morphology: is
it time to reconsider their functional roles in aquatic communities?
Oecologia 120: 621-631

36
Pramuk JB, Hass CA, Hedges SB. 2001. Molecular Phylogeny and Biogeography
of West Indian Toads (Anura: Bufonidae). Molecular Phylogenetics and
Evolution 20(2): 294–301.
Quicke DLJ. 1993. Principles and Techniques of Contemporary Taxonomy.
United Kingdom: Blackie Academic & Professional.
Starrett P. 1960. Descriptions of Tadpoles of Middle American Frogs. Museum of
Zoology, University of Michigan 110: 5-37
Tamura K, Dudley J, Nei M dan Kumar S. 2007. MEGA4: Molecular
evolutionary genetics analysis (MEGA) software version 4.0. Molecular
Biology and Evolution 24: 1596-1599
Tegelstrom H. 1986. Mitochondrial DNA in natural population: An improved
routine for screening of genetic variation based on sensitive silver staining.
Electrophoresis 7(5): 226-229
Tjong DH, Iskandar DT, Gusman D. 2010. Hubungan filogenetik spesies
Limnonectes (Ranidae: Amphibia) asal Sumatera Barat dan asal Asia
Tenggara berdasarkan gen 16S ribosomal RNA. Makara sains 14(1): 79-87.
Toda M, Matsui M, Nishida M, Ota H. 1998. Genetic divergence among
Southeast and East Asian population of Rana limnocharis (Amphibia:
Anura), with species reference to sympatric cryptic species in Java. Zool Sci
14: 607–613
Van Buskirk, J, McCollum, SA. 1999. Plasticity and selection explain variation in
tadpole phenotype between ponds with different predator composition.
Oikos 85: 31-39
Veith M, Kosuch J, Ohler A, Dubois A. 2001. Systematics of Fejervarya
limnocharis (Gravenhorst, 1829) (Amphibia, Anura, Ranidae) and related
species. 2. Morphological and molecular variation in frogs from the Greater
Sunda Islands (Sumatra, Java, Borneo) with the definition of two species.
Alytes 19: 5–28
Verma PK, Pande N. 2002. Learning Amphibia through Latest Portfolio of Theory
and Practice. New Delhi: Dominant Publishers and Distributors.
Wassersug RJ. 1975. The adaptive significance of the tadpole stage with
comments on the maintenance of complex life cycles in anurans. American
Zoology 15: 405-417
Wilbur H. 1987. Regulation of structure in complex systems: experimental
temporary pond communities. Ecology 68: 1437-1452
Wilkinson JA, Drewes RC. 2000. Character assessment, genus level boundaries,
and phylogenetic analyses of the family Rhacophoridae: a review and
present day status. Cont. Herpetol. 2000, 2

37
Wilkinson JA, Drewes RC, Tatum OL. 2002. A molecular phylogenetic analysis
of the family Rhacophoridae with an emphasis on the Asian and African
genera. Mol. Phylogenet. Evol. 24: 265–273.
Yu G, Rao D, Zhang M, Yang J. 2009. Re-examination of the phylogeny of
Rhacophoridae (Anura) based on mitochondrial and nuclear DNA.
Molecular Phylogenetics and Evolution 50: 571–579.

38
LAMPIRAN
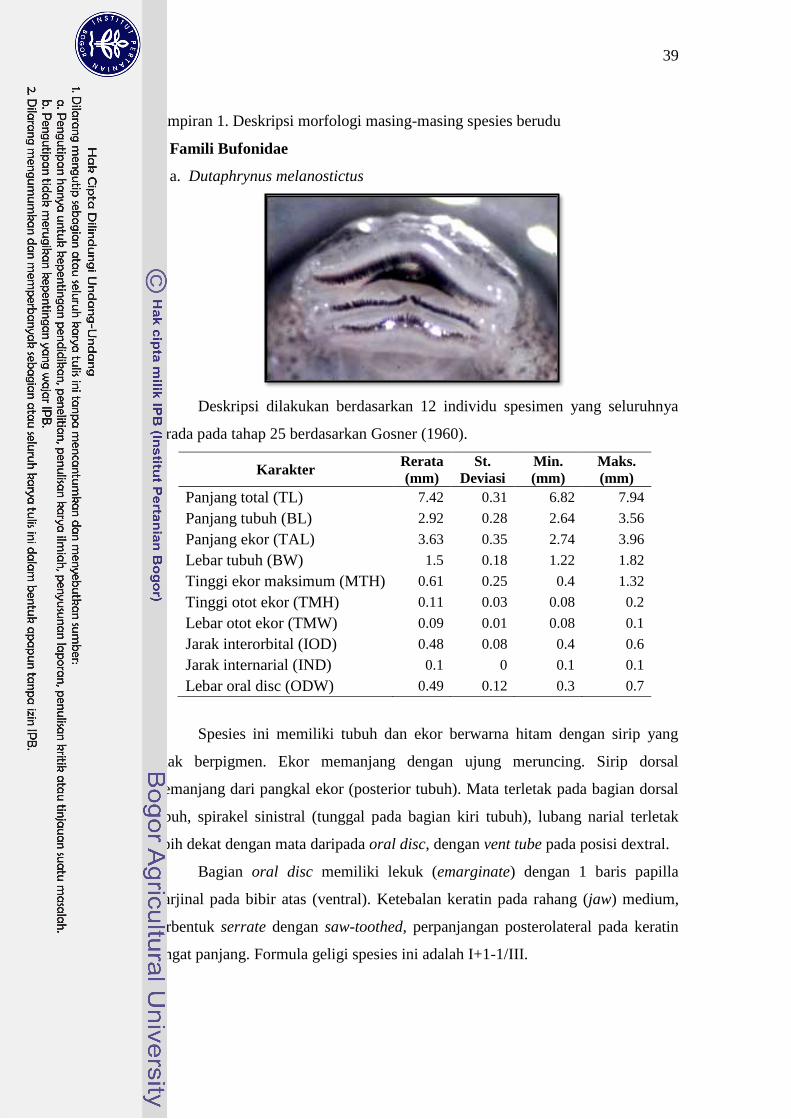
39
Lampiran 1. Deskripsi morfologi masing-masing spesies berudu
1. Famili Bufonidae
a. Dutaphrynus melanostictus
Deskripsi dilakukan berdasarkan 12 individu spesimen yang seluruhnya
berada pada tahap 25 berdasarkan Gosner (1960).
Karakter Rerata
(mm)
St.
Deviasi
Min.
(mm)
Maks.
(mm)
Panjang total (TL) 7.42 0.31 6.82 7.94
Panjang tubuh (BL) 2.92 0.28 2.64 3.56
Panjang ekor (TAL) 3.63 0.35 2.74 3.96
Lebar tubuh (BW) 1.5 0.18 1.22 1.82
Tinggi ekor maksimum (MTH) 0.61 0.25 0.4 1.32
Tinggi otot ekor (TMH) 0.11 0.03 0.08 0.2
Lebar otot ekor (TMW) 0.09 0.01 0.08 0.1
Jarak interorbital (IOD) 0.48 0.08 0.4 0.6
Jarak internarial (IND) 0.1 0 0.1 0.1
Lebar oral disc (ODW) 0.49 0.12 0.3 0.7
Spesies ini memiliki tubuh dan ekor berwarna hitam dengan sirip yang
tidak berpigmen. Ekor memanjang dengan ujung meruncing. Sirip dorsal
memanjang dari pangkal ekor (posterior tubuh). Mata terletak pada bagian dorsal
tubuh, spirakel sinistral (tunggal pada bagian kiri tubuh), lubang narial terletak
lebih dekat dengan mata daripada oral disc, dengan vent tube pada posisi dextral.
Bagian oral disc memiliki lekuk (emarginate) dengan 1 baris papilla
marjinal pada bibir atas (ventral). Ketebalan keratin pada rahang (jaw) medium,
berbentuk serrate dengan saw-toothed, perpanjangan posterolateral pada keratin
sangat panjang. Formula geligi spesies ini adalah I+1-1/III.
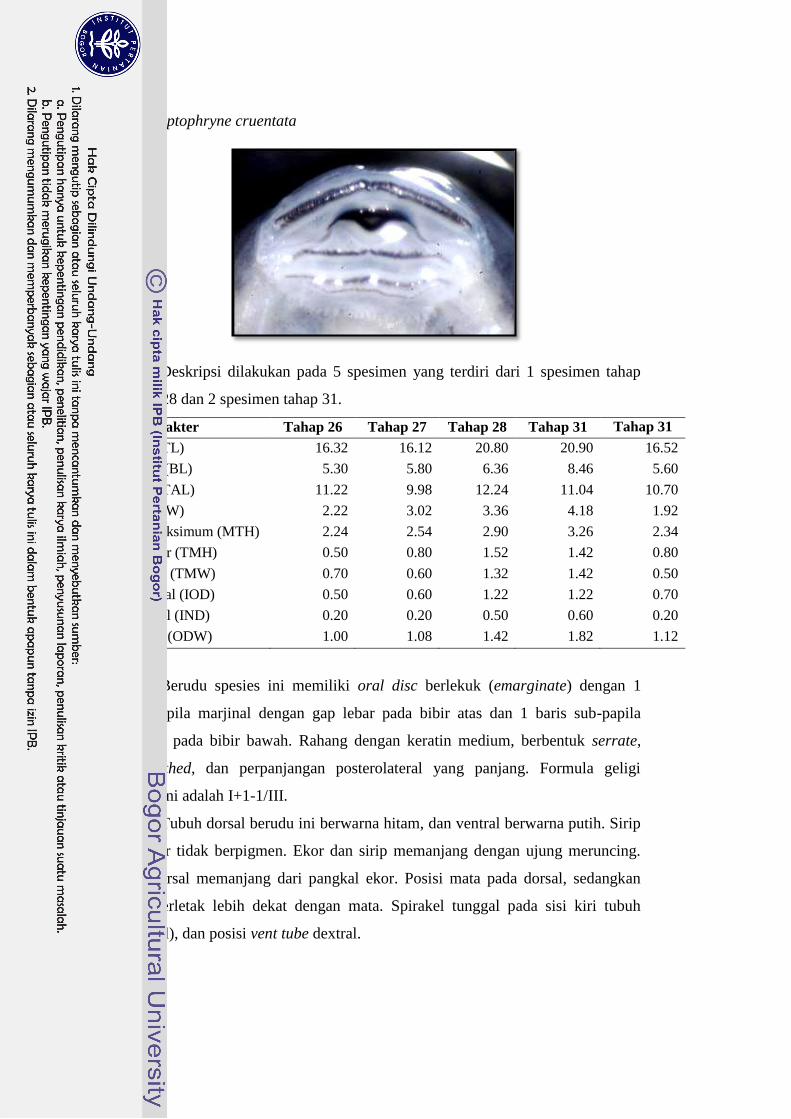
40
b. Leptophryne cruentata
Deskripsi dilakukan pada 5 spesimen yang terdiri dari 1 spesimen tahap
26, 27, 28 dan 2 spesimen tahap 31.
Karakter Tahap 26 Tahap 27 Tahap 28 Tahap 31 Tahap 31
Panjang total (TL) 16.32 16.12 20.80 20.90 16.52
Panjang tubuh (BL) 5.30 5.80 6.36 8.46 5.60
Panjang ekor (TAL) 11.22 9.98 12.24 11.04 10.70
Lebar tubuh (BW) 2.22 3.02 3.36 4.18 1.92
Tinggi ekor maksimum (MTH) 2.24 2.54 2.90 3.26 2.34
Tinggi otot ekor (TMH) 0.50 0.80 1.52 1.42 0.80
Lebar otot ekor (TMW) 0.70 0.60 1.32 1.42 0.50
Jarak interorbital (IOD) 0.50 0.60 1.22 1.22 0.70
Jarak internarial (IND) 0.20 0.20 0.50 0.60 0.20
Lebar oral disc (ODW) 1.00 1.08 1.42 1.82 1.12
Berudu spesies ini memiliki oral disc berlekuk (emarginate) dengan 1
baris papila marjinal dengan gap lebar pada bibir atas dan 1 baris sub-papila
marjinal pada bibir bawah. Rahang dengan keratin medium, berbentuk serrate,
saw-toothed, dan perpanjangan posterolateral yang panjang. Formula geligi
berudu ini adalah I+1-1/III.
Tubuh dorsal berudu ini berwarna hitam, dan ventral berwarna putih. Sirip
dan ekor tidak berpigmen. Ekor dan sirip memanjang dengan ujung meruncing.
Sirip dorsal memanjang dari pangkal ekor. Posisi mata pada dorsal, sedangkan
narial terletak lebih dekat dengan mata. Spirakel tunggal pada sisi kiri tubuh
(sinistral), dan posisi vent tube dextral.
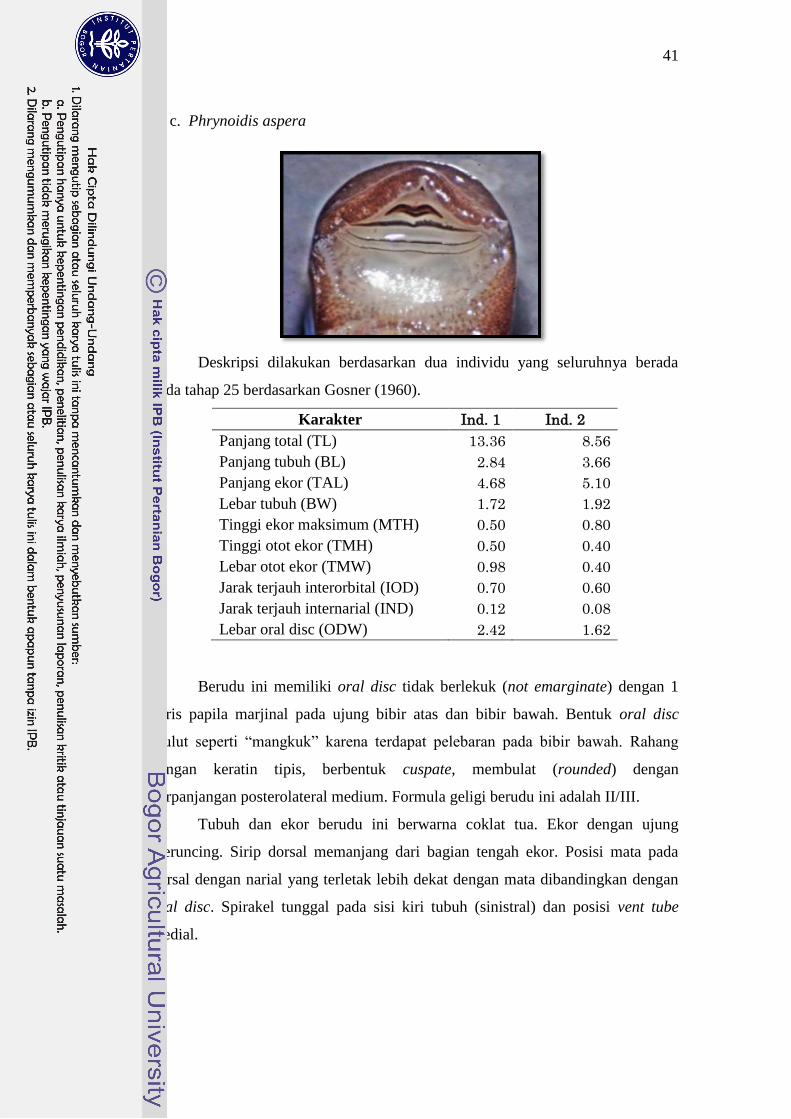
41
c. Phrynoidis aspera
Deskripsi dilakukan berdasarkan dua individu yang seluruhnya berada
pada tahap 25 berdasarkan Gosner (1960).
Karakter Ind. 1 Ind. 2
Panjang total (TL) 13.36 8.56
Panjang tubuh (BL) 2.84 3.66
Panjang ekor (TAL) 4.68 5.10
Lebar tubuh (BW) 1.72 1.92
Tinggi ekor maksimum (MTH) 0.50 0.80
Tinggi otot ekor (TMH) 0.50 0.40
Lebar otot ekor (TMW) 0.98 0.40
Jarak terjauh interorbital (IOD) 0.70 0.60
Jarak terjauh internarial (IND) 0.12 0.08
Lebar oral disc (ODW) 2.42 1.62
Berudu ini memiliki oral disc tidak berlekuk (not emarginate) dengan 1
baris papila marjinal pada ujung bibir atas dan bibir bawah. Bentuk oral disc
mulut seperti “mangkuk” karena terdapat pelebaran pada bibir bawah. Rahang
dengan keratin tipis, berbentuk cuspate, membulat (rounded) dengan
perpanjangan posterolateral medium. Formula geligi berudu ini adalah II/III.
Tubuh dan ekor berudu ini berwarna coklat tua. Ekor dengan ujung
meruncing. Sirip dorsal memanjang dari bagian tengah ekor. Posisi mata pada
dorsal dengan narial yang terletak lebih dekat dengan mata dibandingkan dengan
oral disc. Spirakel tunggal pada sisi kiri tubuh (sinistral) dan posisi vent tube
medial.
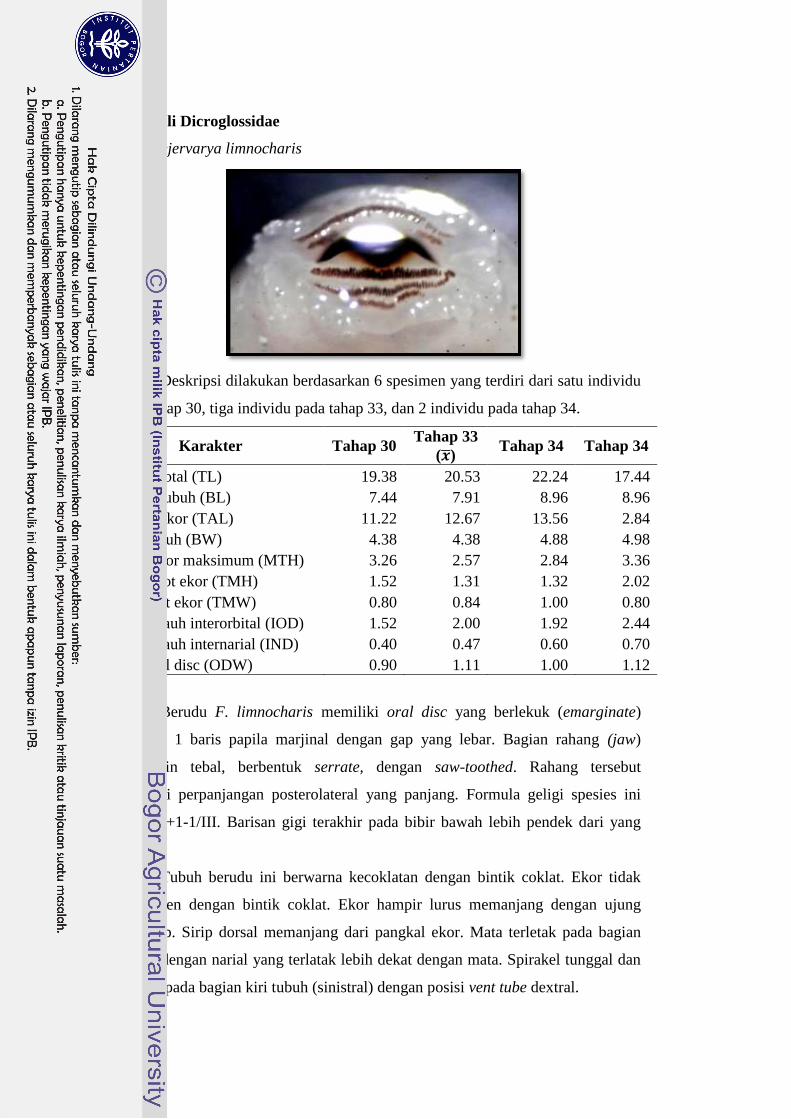
42
2. Famili Dicroglossidae
a. Fejervarya limnocharis
Deskripsi dilakukan berdasarkan 6 spesimen yang terdiri dari satu individu
pada tahap 30, tiga individu pada tahap 33, dan 2 individu pada tahap 34.
Karakter Tahap 30 Tahap 33
(𝒙 ) Tahap 34 Tahap 34
Panjang total (TL) 19.38 20.53 22.24 17.44
Panjang tubuh (BL) 7.44 7.91 8.96 8.96
Panjang ekor (TAL) 11.22 12.67 13.56 2.84
Lebar tubuh (BW) 4.38 4.38 4.88 4.98
Tinggi ekor maksimum (MTH) 3.26 2.57 2.84 3.36
Tinggi otot ekor (TMH) 1.52 1.31 1.32 2.02
Lebar otot ekor (TMW) 0.80 0.84 1.00 0.80
Jarak terjauh interorbital (IOD) 1.52 2.00 1.92 2.44
Jarak terjauh internarial (IND) 0.40 0.47 0.60 0.70
Lebar oral disc (ODW) 0.90 1.11 1.00 1.12
Berudu F. limnocharis memiliki oral disc yang berlekuk (emarginate)
dengan 1 baris papila marjinal dengan gap yang lebar. Bagian rahang (jaw)
berkeratin tebal, berbentuk serrate, dengan saw-toothed. Rahang tersebut
memiliki perpanjangan posterolateral yang panjang. Formula geligi spesies ini
adalah I+1-1/III. Barisan gigi terakhir pada bibir bawah lebih pendek dari yang
lainnya.
Tubuh berudu ini berwarna kecoklatan dengan bintik coklat. Ekor tidak
berpigmen dengan bintik coklat. Ekor hampir lurus memanjang dengan ujung
melancip. Sirip dorsal memanjang dari pangkal ekor. Mata terletak pada bagian
dorsal, dengan narial yang terlatak lebih dekat dengan mata. Spirakel tunggal dan
terletak pada bagian kiri tubuh (sinistral) dengan posisi vent tube dextral.
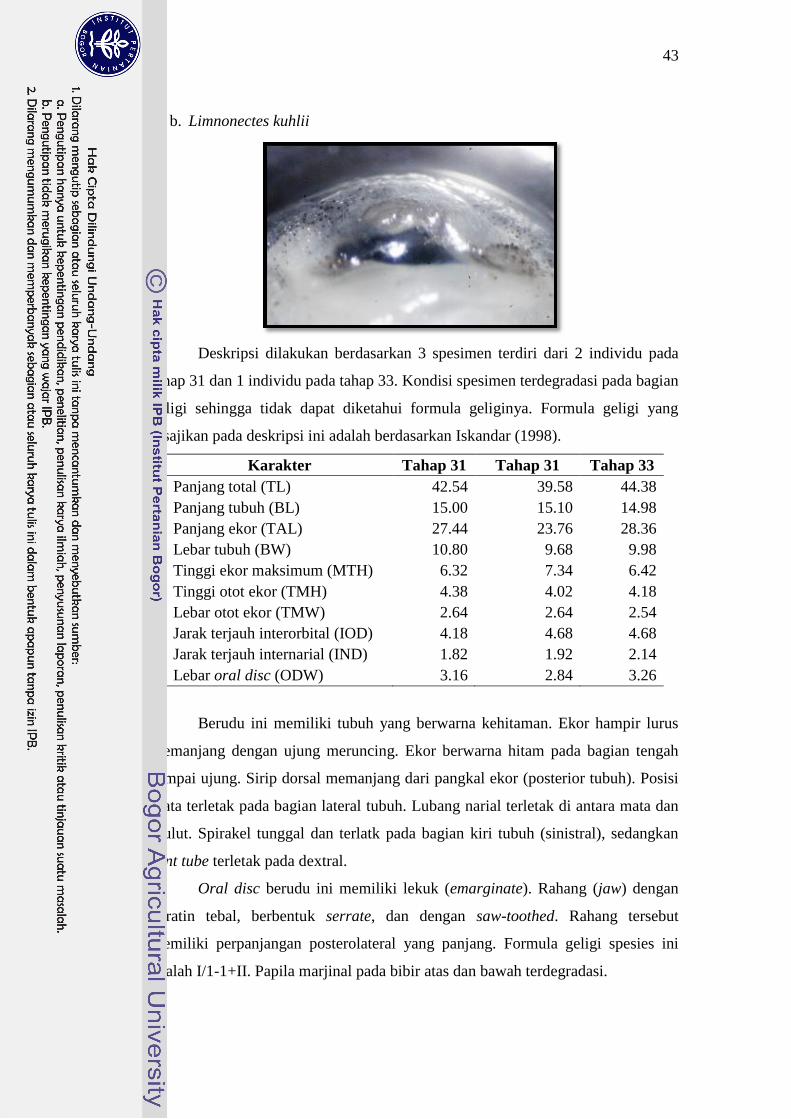
43
b. Limnonectes kuhlii
Deskripsi dilakukan berdasarkan 3 spesimen terdiri dari 2 individu pada
tahap 31 dan 1 individu pada tahap 33. Kondisi spesimen terdegradasi pada bagian
geligi sehingga tidak dapat diketahui formula geliginya. Formula geligi yang
disajikan pada deskripsi ini adalah berdasarkan Iskandar (1998).
Karakter Tahap 31 Tahap 31 Tahap 33
Panjang total (TL) 42.54 39.58 44.38
Panjang tubuh (BL) 15.00 15.10 14.98
Panjang ekor (TAL) 27.44 23.76 28.36
Lebar tubuh (BW) 10.80 9.68 9.98
Tinggi ekor maksimum (MTH) 6.32 7.34 6.42
Tinggi otot ekor (TMH) 4.38 4.02 4.18
Lebar otot ekor (TMW) 2.64 2.64 2.54
Jarak terjauh interorbital (IOD) 4.18 4.68 4.68
Jarak terjauh internarial (IND) 1.82 1.92 2.14
Lebar oral disc (ODW) 3.16 2.84 3.26
Berudu ini memiliki tubuh yang berwarna kehitaman. Ekor hampir lurus
memanjang dengan ujung meruncing. Ekor berwarna hitam pada bagian tengah
sampai ujung. Sirip dorsal memanjang dari pangkal ekor (posterior tubuh). Posisi
mata terletak pada bagian lateral tubuh. Lubang narial terletak di antara mata dan
mulut. Spirakel tunggal dan terlatk pada bagian kiri tubuh (sinistral), sedangkan
vent tube terletak pada dextral.
Oral disc berudu ini memiliki lekuk (emarginate). Rahang (jaw) dengan
keratin tebal, berbentuk serrate, dan dengan saw-toothed. Rahang tersebut
memiliki perpanjangan posterolateral yang panjang. Formula geligi spesies ini
adalah I/1-1+II. Papila marjinal pada bibir atas dan bawah terdegradasi.
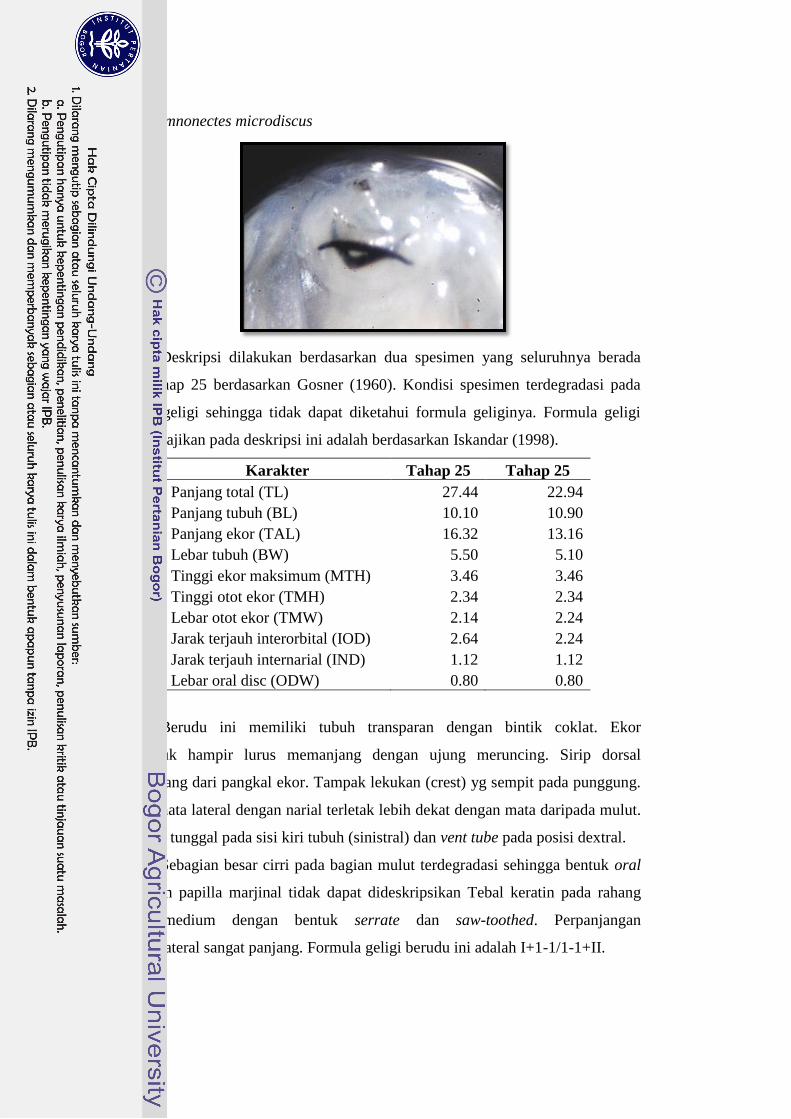
44
c. Limnonectes microdiscus
Deskripsi dilakukan berdasarkan dua spesimen yang seluruhnya berada
pada tahap 25 berdasarkan Gosner (1960). Kondisi spesimen terdegradasi pada
bagian geligi sehingga tidak dapat diketahui formula geliginya. Formula geligi
yang disajikan pada deskripsi ini adalah berdasarkan Iskandar (1998).
Karakter Tahap 25 Tahap 25
Panjang total (TL) 27.44 22.94
Panjang tubuh (BL) 10.10 10.90
Panjang ekor (TAL) 16.32 13.16
Lebar tubuh (BW) 5.50 5.10
Tinggi ekor maksimum (MTH) 3.46 3.46
Tinggi otot ekor (TMH) 2.34 2.34
Lebar otot ekor (TMW) 2.14 2.24
Jarak terjauh interorbital (IOD) 2.64 2.24
Jarak terjauh internarial (IND) 1.12 1.12
Lebar oral disc (ODW) 0.80 0.80
Berudu ini memiliki tubuh transparan dengan bintik coklat. Ekor
berbentuk hampir lurus memanjang dengan ujung meruncing. Sirip dorsal
memanjang dari pangkal ekor. Tampak lekukan (crest) yg sempit pada punggung.
Posisi mata lateral dengan narial terletak lebih dekat dengan mata daripada mulut.
Spirakel tunggal pada sisi kiri tubuh (sinistral) dan vent tube pada posisi dextral.
Sebagian besar cirri pada bagian mulut terdegradasi sehingga bentuk oral
disc, dan papilla marjinal tidak dapat dideskripsikan Tebal keratin pada rahang
(jaw) medium dengan bentuk serrate dan saw-toothed. Perpanjangan
posterolateral sangat panjang. Formula geligi berudu ini adalah I+1-1/1-1+II.
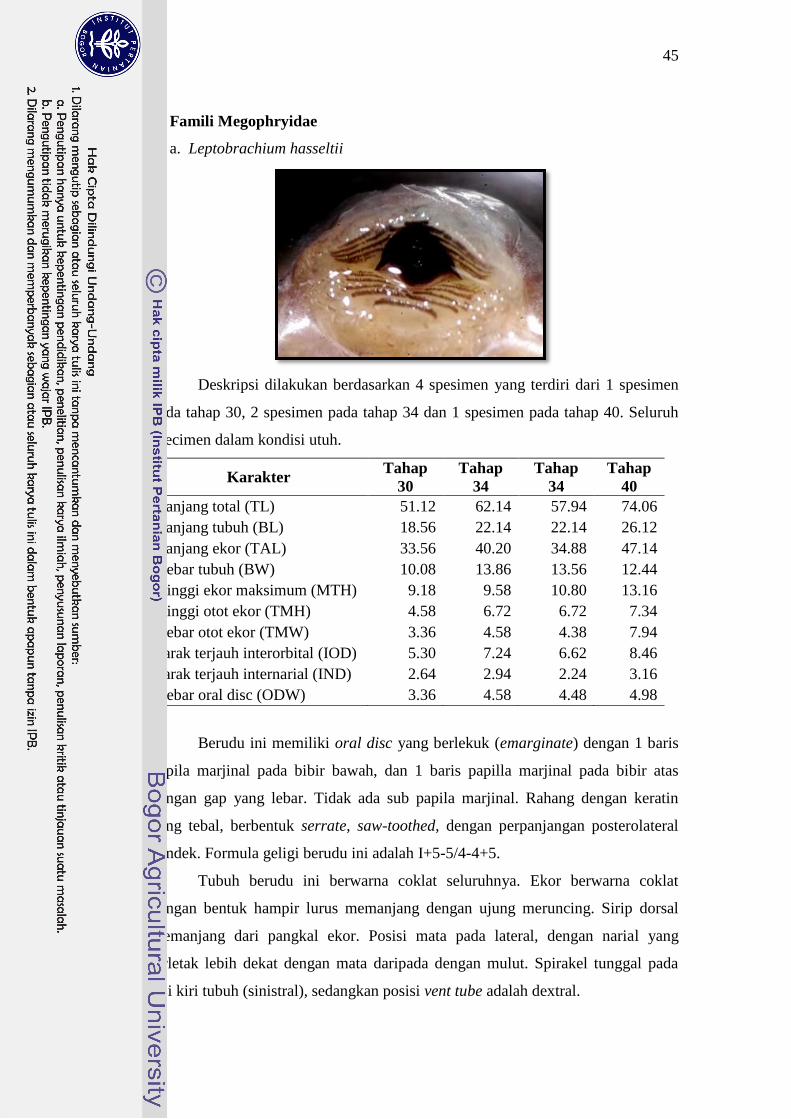
45
3. Famili Megophryidae
a. Leptobrachium hasseltii
Deskripsi dilakukan berdasarkan 4 spesimen yang terdiri dari 1 spesimen
pada tahap 30, 2 spesimen pada tahap 34 dan 1 spesimen pada tahap 40. Seluruh
specimen dalam kondisi utuh.
Karakter Tahap
30
Tahap
34
Tahap
34
Tahap
40
Panjang total (TL) 51.12 62.14 57.94 74.06
Panjang tubuh (BL) 18.56 22.14 22.14 26.12
Panjang ekor (TAL) 33.56 40.20 34.88 47.14
Lebar tubuh (BW) 10.08 13.86 13.56 12.44
Tinggi ekor maksimum (MTH) 9.18 9.58 10.80 13.16
Tinggi otot ekor (TMH) 4.58 6.72 6.72 7.34
Lebar otot ekor (TMW) 3.36 4.58 4.38 7.94
Jarak terjauh interorbital (IOD) 5.30 7.24 6.62 8.46
Jarak terjauh internarial (IND) 2.64 2.94 2.24 3.16
Lebar oral disc (ODW) 3.36 4.58 4.48 4.98
Berudu ini memiliki oral disc yang berlekuk (emarginate) dengan 1 baris
papila marjinal pada bibir bawah, dan 1 baris papilla marjinal pada bibir atas
dengan gap yang lebar. Tidak ada sub papila marjinal. Rahang dengan keratin
yang tebal, berbentuk serrate, saw-toothed, dengan perpanjangan posterolateral
pendek. Formula geligi berudu ini adalah I+5-5/4-4+5.
Tubuh berudu ini berwarna coklat seluruhnya. Ekor berwarna coklat
dengan bentuk hampir lurus memanjang dengan ujung meruncing. Sirip dorsal
memanjang dari pangkal ekor. Posisi mata pada lateral, dengan narial yang
terletak lebih dekat dengan mata daripada dengan mulut. Spirakel tunggal pada
sisi kiri tubuh (sinistral), sedangkan posisi vent tube adalah dextral.
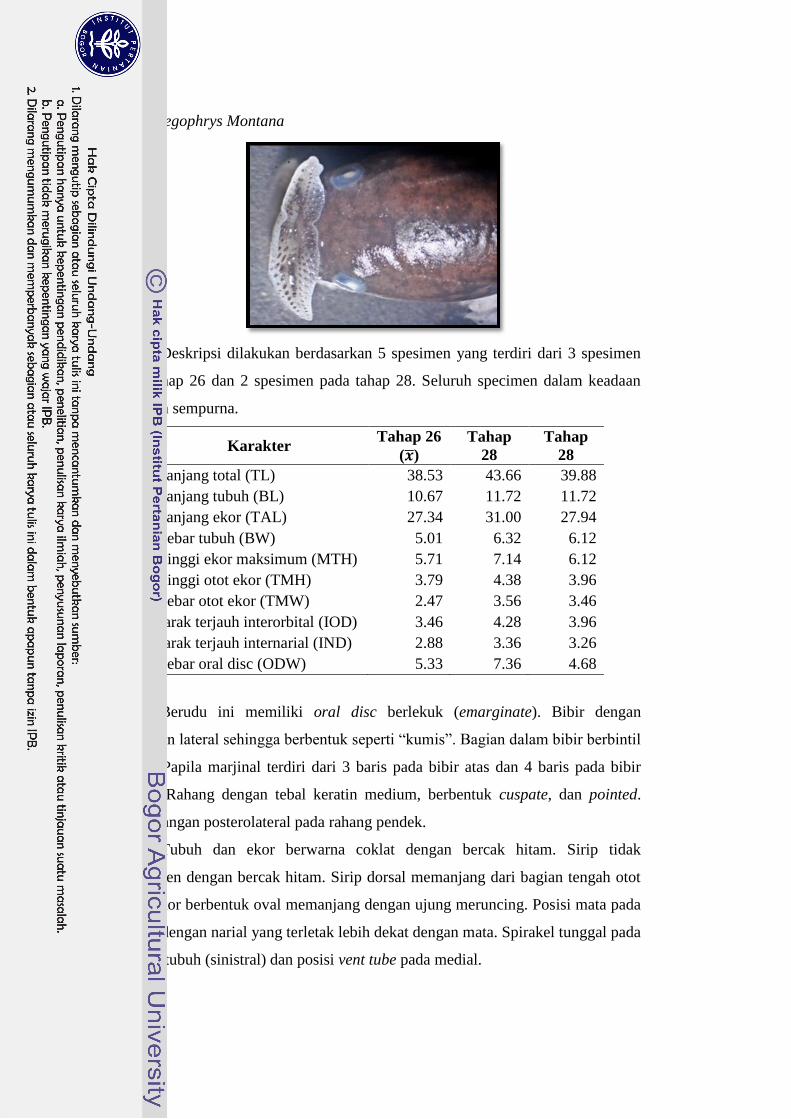
46
b. Megophrys Montana
Deskripsi dilakukan berdasarkan 5 spesimen yang terdiri dari 3 spesimen
pada tahap 26 dan 2 spesimen pada tahap 28. Seluruh specimen dalam keadaan
utuh dan sempurna.
Karakter Tahap 26
(𝒙 )
Tahap
28
Tahap
28
Panjang total (TL) 38.53 43.66 39.88
Panjang tubuh (BL) 10.67 11.72 11.72
Panjang ekor (TAL) 27.34 31.00 27.94
Lebar tubuh (BW) 5.01 6.32 6.12
Tinggi ekor maksimum (MTH) 5.71 7.14 6.12
Tinggi otot ekor (TMH) 3.79 4.38 3.96
Lebar otot ekor (TMW) 2.47 3.56 3.46
Jarak terjauh interorbital (IOD) 3.46 4.28 3.96
Jarak terjauh internarial (IND) 2.88 3.36 3.26
Lebar oral disc (ODW) 5.33 7.36 4.68
Berudu ini memiliki oral disc berlekuk (emarginate). Bibir dengan
pelebaran lateral sehingga berbentuk seperti “kumis”. Bagian dalam bibir berbintil
coklat. Papila marjinal terdiri dari 3 baris pada bibir atas dan 4 baris pada bibir
bawah. Rahang dengan tebal keratin medium, berbentuk cuspate, dan pointed.
Perpanjangan posterolateral pada rahang pendek.
Tubuh dan ekor berwarna coklat dengan bercak hitam. Sirip tidak
berpigmen dengan bercak hitam. Sirip dorsal memanjang dari bagian tengah otot
ekor. Ekor berbentuk oval memanjang dengan ujung meruncing. Posisi mata pada
lateral, dengan narial yang terletak lebih dekat dengan mata. Spirakel tunggal pada
sisi kiri tubuh (sinistral) dan posisi vent tube pada medial.
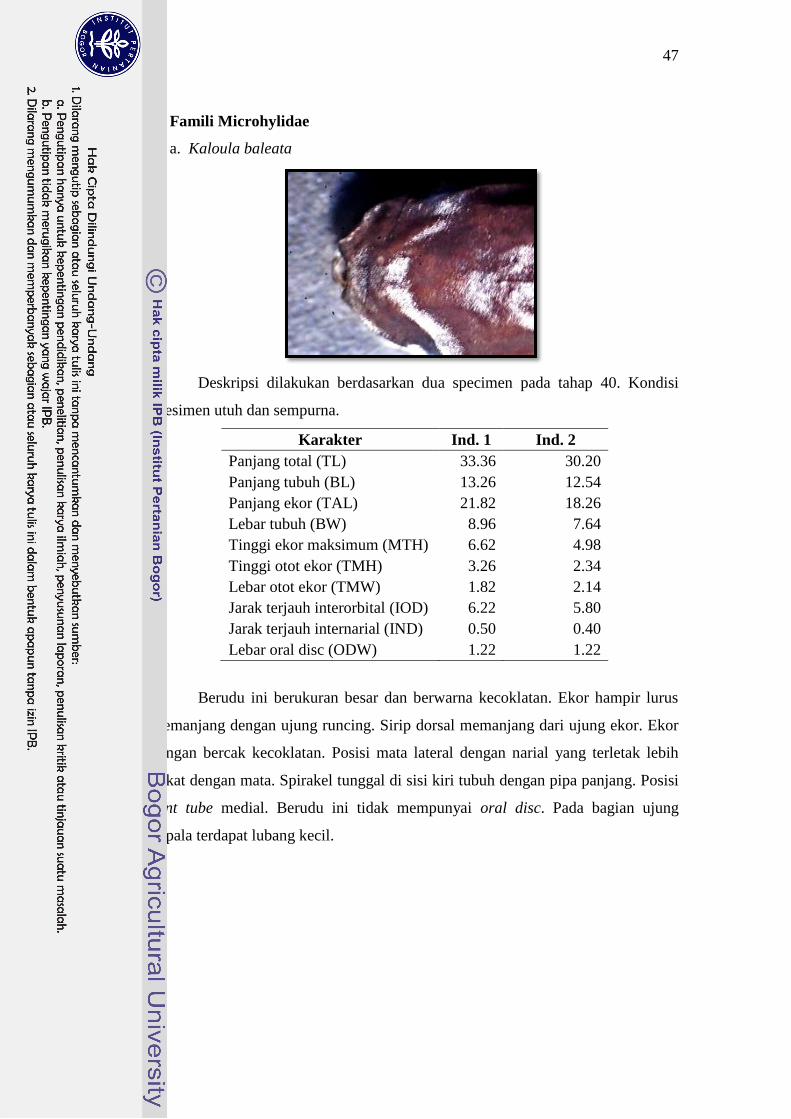
47
4. Famili Microhylidae
a. Kaloula baleata
Deskripsi dilakukan berdasarkan dua specimen pada tahap 40. Kondisi
spesimen utuh dan sempurna.
Karakter Ind. 1 Ind. 2
Panjang total (TL) 33.36 30.20
Panjang tubuh (BL) 13.26 12.54
Panjang ekor (TAL) 21.82 18.26
Lebar tubuh (BW) 8.96 7.64
Tinggi ekor maksimum (MTH) 6.62 4.98
Tinggi otot ekor (TMH) 3.26 2.34
Lebar otot ekor (TMW) 1.82 2.14
Jarak terjauh interorbital (IOD) 6.22 5.80
Jarak terjauh internarial (IND) 0.50 0.40
Lebar oral disc (ODW) 1.22 1.22
Berudu ini berukuran besar dan berwarna kecoklatan. Ekor hampir lurus
memanjang dengan ujung runcing. Sirip dorsal memanjang dari ujung ekor. Ekor
dengan bercak kecoklatan. Posisi mata lateral dengan narial yang terletak lebih
dekat dengan mata. Spirakel tunggal di sisi kiri tubuh dengan pipa panjang. Posisi
vent tube medial. Berudu ini tidak mempunyai oral disc. Pada bagian ujung
kepala terdapat lubang kecil.
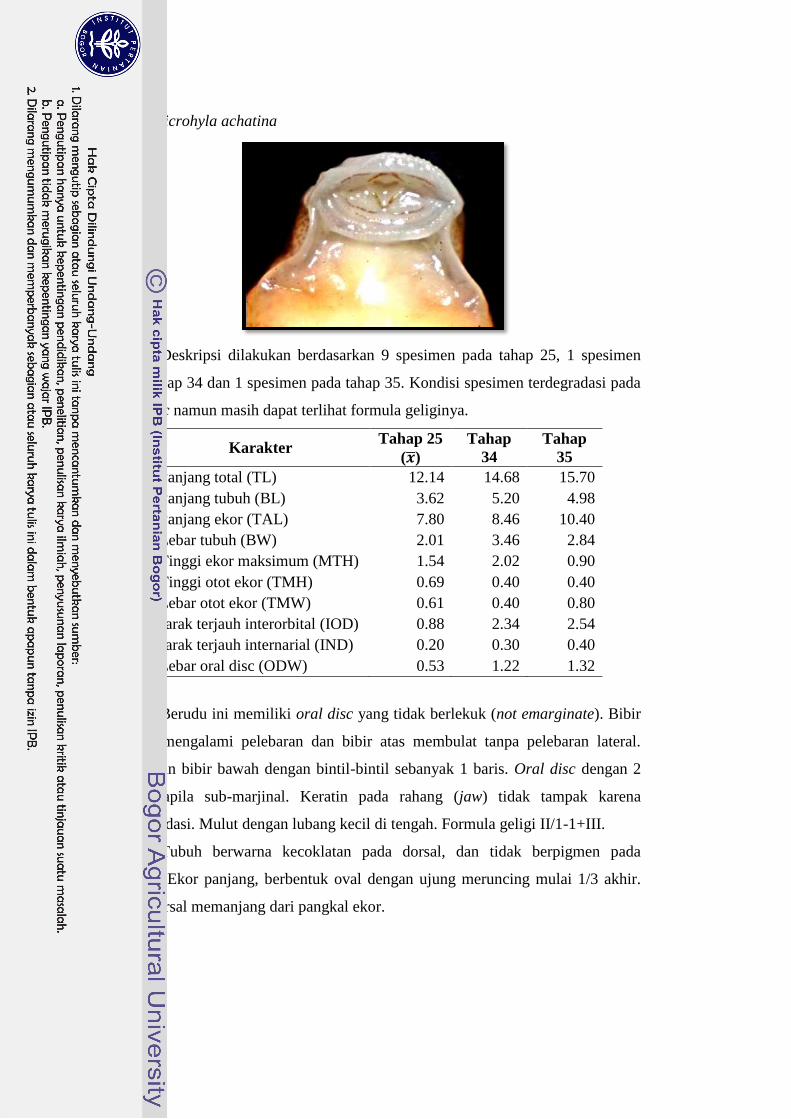
48
b. Microhyla achatina
Deskripsi dilakukan berdasarkan 9 spesimen pada tahap 25, 1 spesimen
pada tahap 34 dan 1 spesimen pada tahap 35. Kondisi spesimen terdegradasi pada
oral disc namun masih dapat terlihat formula geliginya.
Karakter Tahap 25
(𝒙 )
Tahap
34
Tahap
35
Panjang total (TL) 12.14 14.68 15.70
Panjang tubuh (BL) 3.62 5.20 4.98
Panjang ekor (TAL) 7.80 8.46 10.40
Lebar tubuh (BW) 2.01 3.46 2.84
Tinggi ekor maksimum (MTH) 1.54 2.02 0.90
Tinggi otot ekor (TMH) 0.69 0.40 0.40
Lebar otot ekor (TMW) 0.61 0.40 0.80
Jarak terjauh interorbital (IOD) 0.88 2.34 2.54
Jarak terjauh internarial (IND) 0.20 0.30 0.40
Lebar oral disc (ODW) 0.53 1.22 1.32
Berudu ini memiliki oral disc yang tidak berlekuk (not emarginate). Bibir
bawah mengalami pelebaran dan bibir atas membulat tanpa pelebaran lateral.
Pelebaran bibir bawah dengan bintil-bintil sebanyak 1 baris. Oral disc dengan 2
baris papila sub-marjinal. Keratin pada rahang (jaw) tidak tampak karena
terdegradasi. Mulut dengan lubang kecil di tengah. Formula geligi II/1-1+III.
Tubuh berwarna kecoklatan pada dorsal, dan tidak berpigmen pada
ventral. Ekor panjang, berbentuk oval dengan ujung meruncing mulai 1/3 akhir.
Sirip dorsal memanjang dari pangkal ekor.
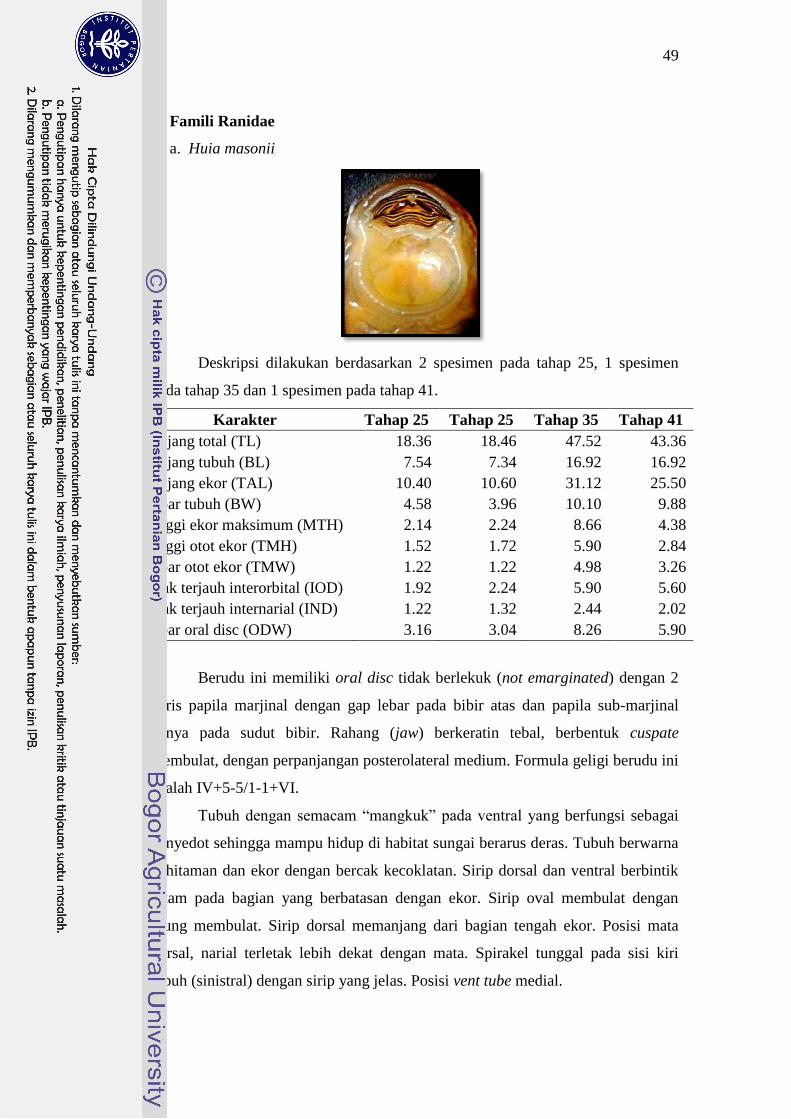
49
5. Famili Ranidae
a. Huia masonii
Deskripsi dilakukan berdasarkan 2 spesimen pada tahap 25, 1 spesimen
pada tahap 35 dan 1 spesimen pada tahap 41.
Karakter Tahap 25 Tahap 25 Tahap 35 Tahap 41
Panjang total (TL) 18.36 18.46 47.52 43.36
Panjang tubuh (BL) 7.54 7.34 16.92 16.92
Panjang ekor (TAL) 10.40 10.60 31.12 25.50
Lebar tubuh (BW) 4.58 3.96 10.10 9.88
Tinggi ekor maksimum (MTH) 2.14 2.24 8.66 4.38
Tinggi otot ekor (TMH) 1.52 1.72 5.90 2.84
Lebar otot ekor (TMW) 1.22 1.22 4.98 3.26
Jarak terjauh interorbital (IOD) 1.92 2.24 5.90 5.60
Jarak terjauh internarial (IND) 1.22 1.32 2.44 2.02
Lebar oral disc (ODW) 3.16 3.04 8.26 5.90
Berudu ini memiliki oral disc tidak berlekuk (not emarginated) dengan 2
baris papila marjinal dengan gap lebar pada bibir atas dan papila sub-marjinal
hanya pada sudut bibir. Rahang (jaw) berkeratin tebal, berbentuk cuspate
membulat, dengan perpanjangan posterolateral medium. Formula geligi berudu ini
adalah IV+5-5/1-1+VI.
Tubuh dengan semacam “mangkuk” pada ventral yang berfungsi sebagai
penyedot sehingga mampu hidup di habitat sungai berarus deras. Tubuh berwarna
kehitaman dan ekor dengan bercak kecoklatan. Sirip dorsal dan ventral berbintik
hitam pada bagian yang berbatasan dengan ekor. Sirip oval membulat dengan
ujung membulat. Sirip dorsal memanjang dari bagian tengah ekor. Posisi mata
dorsal, narial terletak lebih dekat dengan mata. Spirakel tunggal pada sisi kiri
tubuh (sinistral) dengan sirip yang jelas. Posisi vent tube medial.

50
b. Hylarana chalconota
Deskripsi dilakukan berdasarkan 2 spesimen yang terdiri dari 1 spesimen
pada tahap 32 dan 1 spesimen pada tahap 34.
Karakter Tahap 32 Tahap 34
Panjang total (TL) 25.08 30.60
Panjang tubuh (BL) 9.28 10.40
Panjang ekor (TAL) 17.64 19.48
Lebar tubuh (BW) 5.08 5.60
Tinggi ekor maksimum (MTH) 4.58 5.20
Tinggi otot ekor (TMH) 2.64 2.16
Lebar otot ekor (TMW) 2.24 2.02
Jarak terjauh interorbital (IOD) 2.84 3.56
Jarak terjauh internarial (IND) 1.32 1.42
Lebar oral disc (ODW) 2.02 2.34
Berudu ini memiliki oral disc tidak berlekuk (not-emarginate) dengan 1
baris papila marjinal hanya pada sudut mulut dan 1 baris papila sub-marjinal.
Rahang dengan keratin tebal, berbentuk serrate, saw-toothed dan dengan
perpanjangan posterolateral panjang. Formula geligi adalah I+3-3/III.
Tubuh kuning kecoklatan sedangkan ekor tidak berpigmen. Ekor oval
memanjang dengan ujung meruncing. Sirip dorsal memanjang dari pangkal ekor.
Posisi mata lateral, narial terletak di antara mata dan oral disc. Spirakel tunggal
(sinistral) dan vent tube medial.
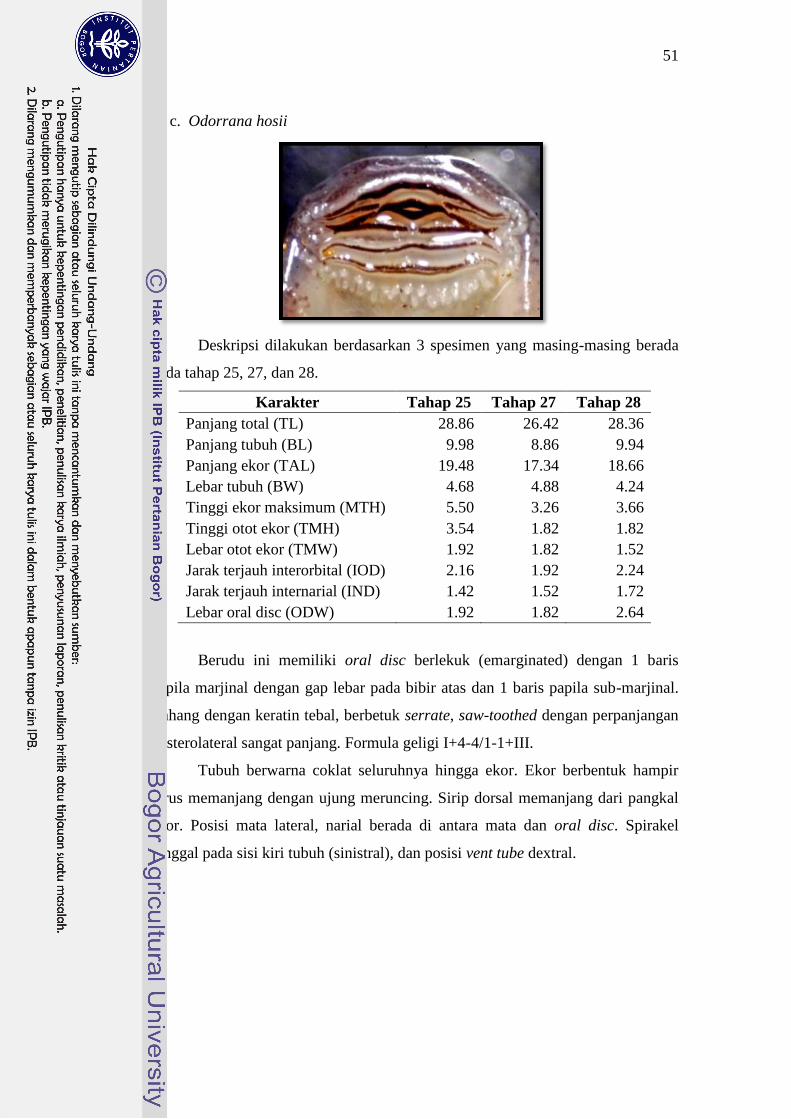
51
c. Odorrana hosii
Deskripsi dilakukan berdasarkan 3 spesimen yang masing-masing berada
pada tahap 25, 27, dan 28.
Karakter Tahap 25 Tahap 27 Tahap 28
Panjang total (TL) 28.86 26.42 28.36
Panjang tubuh (BL) 9.98 8.86 9.94
Panjang ekor (TAL) 19.48 17.34 18.66
Lebar tubuh (BW) 4.68 4.88 4.24
Tinggi ekor maksimum (MTH) 5.50 3.26 3.66
Tinggi otot ekor (TMH) 3.54 1.82 1.82
Lebar otot ekor (TMW) 1.92 1.82 1.52
Jarak terjauh interorbital (IOD) 2.16 1.92 2.24
Jarak terjauh internarial (IND) 1.42 1.52 1.72
Lebar oral disc (ODW) 1.92 1.82 2.64
Berudu ini memiliki oral disc berlekuk (emarginated) dengan 1 baris
papila marjinal dengan gap lebar pada bibir atas dan 1 baris papila sub-marjinal.
Rahang dengan keratin tebal, berbetuk serrate, saw-toothed dengan perpanjangan
posterolateral sangat panjang. Formula geligi I+4-4/1-1+III.
Tubuh berwarna coklat seluruhnya hingga ekor. Ekor berbentuk hampir
lurus memanjang dengan ujung meruncing. Sirip dorsal memanjang dari pangkal
ekor. Posisi mata lateral, narial berada di antara mata dan oral disc. Spirakel
tunggal pada sisi kiri tubuh (sinistral), dan posisi vent tube dextral.

52
6. Famili Rhacophoridae
a. Philautus vittiger
Deskripsi dilakukan berdasarkan 7 spesimen yang terdiri dari 1 spesimen
tahap 28, 2 spesimen pada tahap 29, 2 spesimen pada tahap 35, 1 spesimen pada
tahap 36, dan 1 spesimen pada tahap 41.
Karakter Tahap
28
Tahap
29
Tahap
29
Tahap
35
Tahap
35
Tahap
36
Tahap
41
Panjang total (TL) 24.68 24.78 24.38 27.24 21.32 25.40 24.98
Panjang tubuh (BL) 8.46 8.46 8.74 9.88 8.66 9.88 7.24
Panjang ekor (TAL) 16.62 16.42 16.42 18.96 11.42 17.54 16.92
Lebar tubuh (BW) 4.84 5.40 5.90 5.60 5.30 5.40 4.98
Tinggi ekor maksimum (MTH) 3.96 3.26 3.16 4.28 4.68 3.26 4.68
Tinggi otot ekor (TMH) 1.32 1.92 2.24 2.14 2.24 2.34 2.34
Lebar otot ekor (TMW) 1.22 1.52 1.52 1.42 1.72 1.92 1.82
Jarak terjauh interorbital (IOD) 3.36 3.16 2.94 3.66 3.86 3.66 3.66
Jarak terjauh internarial (IND) 1.32 1.22 1.32 1.32 1.22 1.32 1.32
Lebar oral disc (ODW) 2.44 2.94 3.16 2.84 2.54 3.04 2.24
Berudu ini memiliki oral disc berlekuk (emarginate) dengan 2 baris papila
marginal dengan gap yang lebar pada bibir atas dan 3 baris papilla sub-marjinal
pada bibir bawah. Rahang berkeratin medium, berbentuk cuspate, rounded dengan
perpanjangan posterolateral medium. Formula geligi spesies ini II+4-4/III.
Pola warna tubuh coklat kemerahan dengan bintik hitam, ekor dan sirip
dorsal hitam dan menjelang ujung ekor berpola. Sirip dorsal memanjang dari
pangkal otot ekor (posterior tubuh). Ekor oval dengan ujung meruncing. Posisi
mata lateral, narial lebih dekat dengan mata. Spirakel tunggal pada sisi kiri tubuh
(sinistral) dan vent tube dextral.
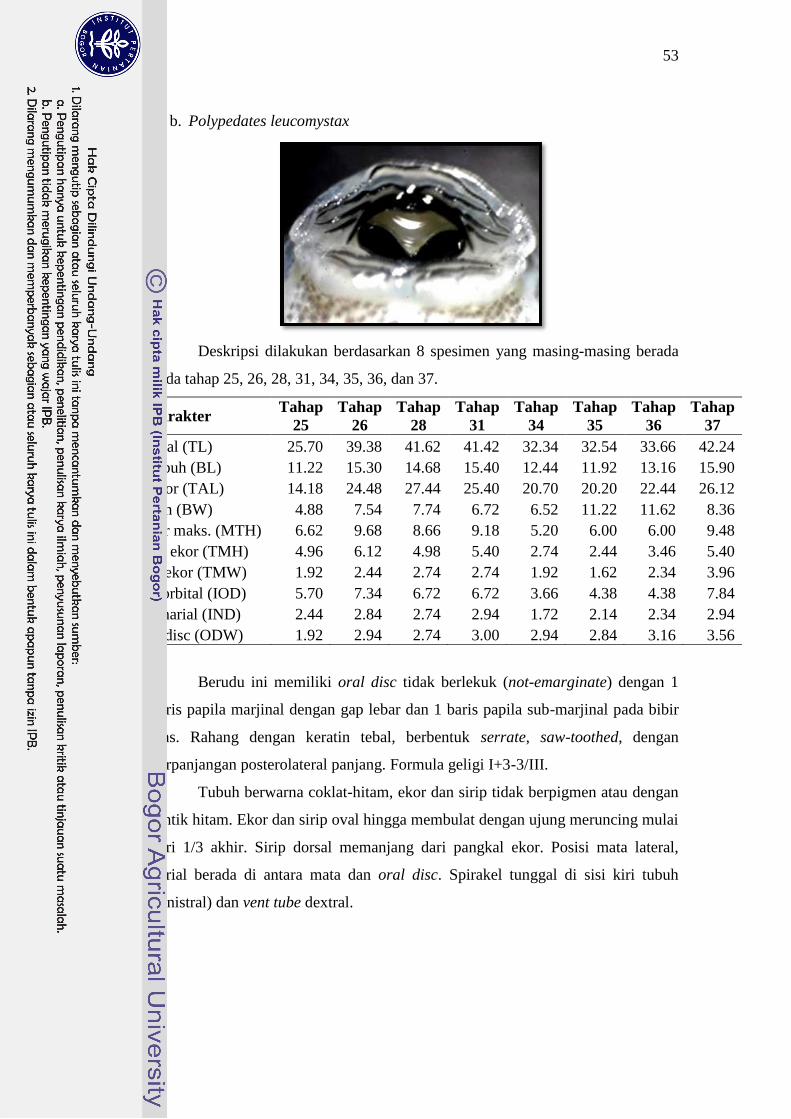
53
b. Polypedates leucomystax
Deskripsi dilakukan berdasarkan 8 spesimen yang masing-masing berada
pada tahap 25, 26, 28, 31, 34, 35, 36, dan 37.
Karakter Tahap
25
Tahap
26
Tahap
28
Tahap
31
Tahap
34
Tahap
35
Tahap
36
Tahap
37
Panjang total (TL) 25.70 39.38 41.62 41.42 32.34 32.54 33.66 42.24
Panjang tubuh (BL) 11.22 15.30 14.68 15.40 12.44 11.92 13.16 15.90
Panjang ekor (TAL) 14.18 24.48 27.44 25.40 20.70 20.20 22.44 26.12
Lebar tubuh (BW) 4.88 7.54 7.74 6.72 6.52 11.22 11.62 8.36
Tinggi ekor maks. (MTH) 6.62 9.68 8.66 9.18 5.20 6.00 6.00 9.48
Tinggi otot ekor (TMH) 4.96 6.12 4.98 5.40 2.74 2.44 3.46 5.40
Lebar otot ekor (TMW) 1.92 2.44 2.74 2.74 1.92 1.62 2.34 3.96
Jarak interorbital (IOD) 5.70 7.34 6.72 6.72 3.66 4.38 4.38 7.84
Jarak internarial (IND) 2.44 2.84 2.74 2.94 1.72 2.14 2.34 2.94
Lebar oral disc (ODW) 1.92 2.94 2.74 3.00 2.94 2.84 3.16 3.56
Berudu ini memiliki oral disc tidak berlekuk (not-emarginate) dengan 1
baris papila marjinal dengan gap lebar dan 1 baris papila sub-marjinal pada bibir
atas. Rahang dengan keratin tebal, berbentuk serrate, saw-toothed, dengan
perpanjangan posterolateral panjang. Formula geligi I+3-3/III.
Tubuh berwarna coklat-hitam, ekor dan sirip tidak berpigmen atau dengan
bintik hitam. Ekor dan sirip oval hingga membulat dengan ujung meruncing mulai
dari 1/3 akhir. Sirip dorsal memanjang dari pangkal ekor. Posisi mata lateral,
narial berada di antara mata dan oral disc. Spirakel tunggal di sisi kiri tubuh
(sinistral) dan vent tube dextral.
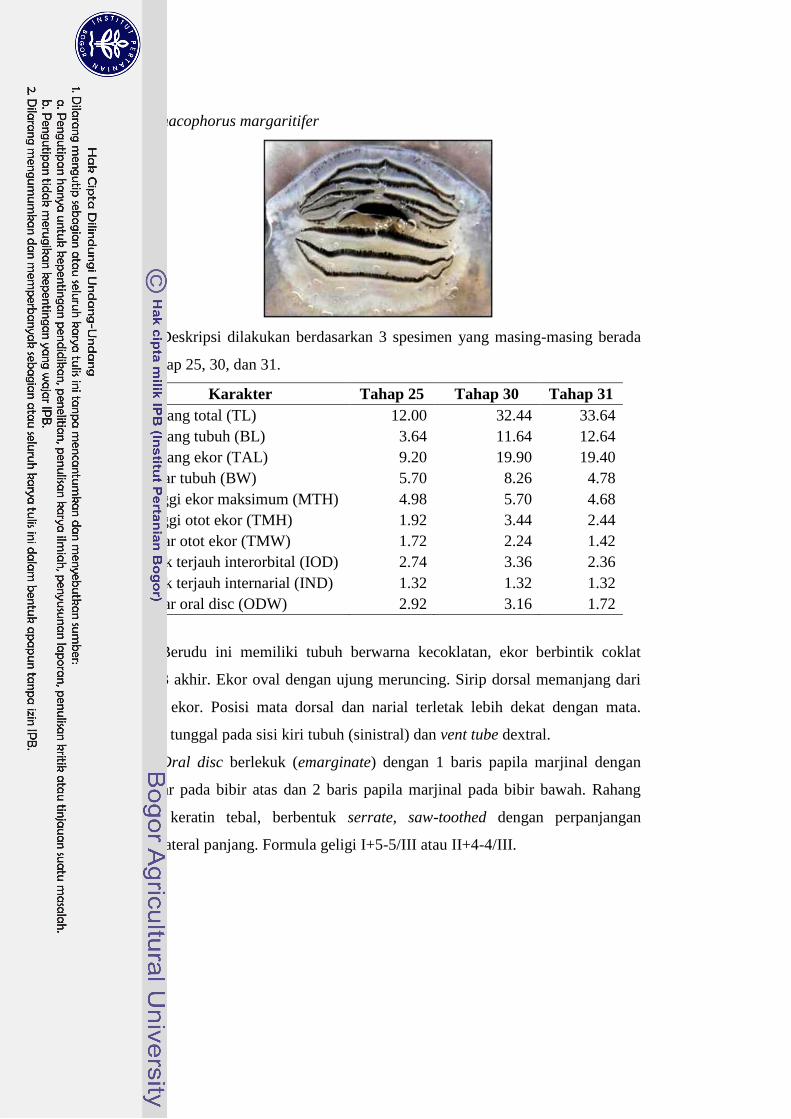
54
c. Rhacophorus margaritifer
Deskripsi dilakukan berdasarkan 3 spesimen yang masing-masing berada
pada tahap 25, 30, dan 31.
Karakter Tahap 25 Tahap 30 Tahap 31
Panjang total (TL) 12.00 32.44 33.64
Panjang tubuh (BL) 3.64 11.64 12.64
Panjang ekor (TAL) 9.20 19.90 19.40
Lebar tubuh (BW) 5.70 8.26 4.78
Tinggi ekor maksimum (MTH) 4.98 5.70 4.68
Tinggi otot ekor (TMH) 1.92 3.44 2.44
Lebar otot ekor (TMW) 1.72 2.24 1.42
Jarak terjauh interorbital (IOD) 2.74 3.36 2.36
Jarak terjauh internarial (IND) 1.32 1.32 1.32
Lebar oral disc (ODW) 2.92 3.16 1.72
Berudu ini memiliki tubuh berwarna kecoklatan, ekor berbintik coklat
pada 1/3 akhir. Ekor oval dengan ujung meruncing. Sirip dorsal memanjang dari
pangkal ekor. Posisi mata dorsal dan narial terletak lebih dekat dengan mata.
Spirakel tunggal pada sisi kiri tubuh (sinistral) dan vent tube dextral.
Oral disc berlekuk (emarginate) dengan 1 baris papila marjinal dengan
gap lebar pada bibir atas dan 2 baris papila marjinal pada bibir bawah. Rahang
dengan keratin tebal, berbentuk serrate, saw-toothed dengan perpanjangan
posterolateral panjang. Formula geligi I+5-5/III atau II+4-4/III.
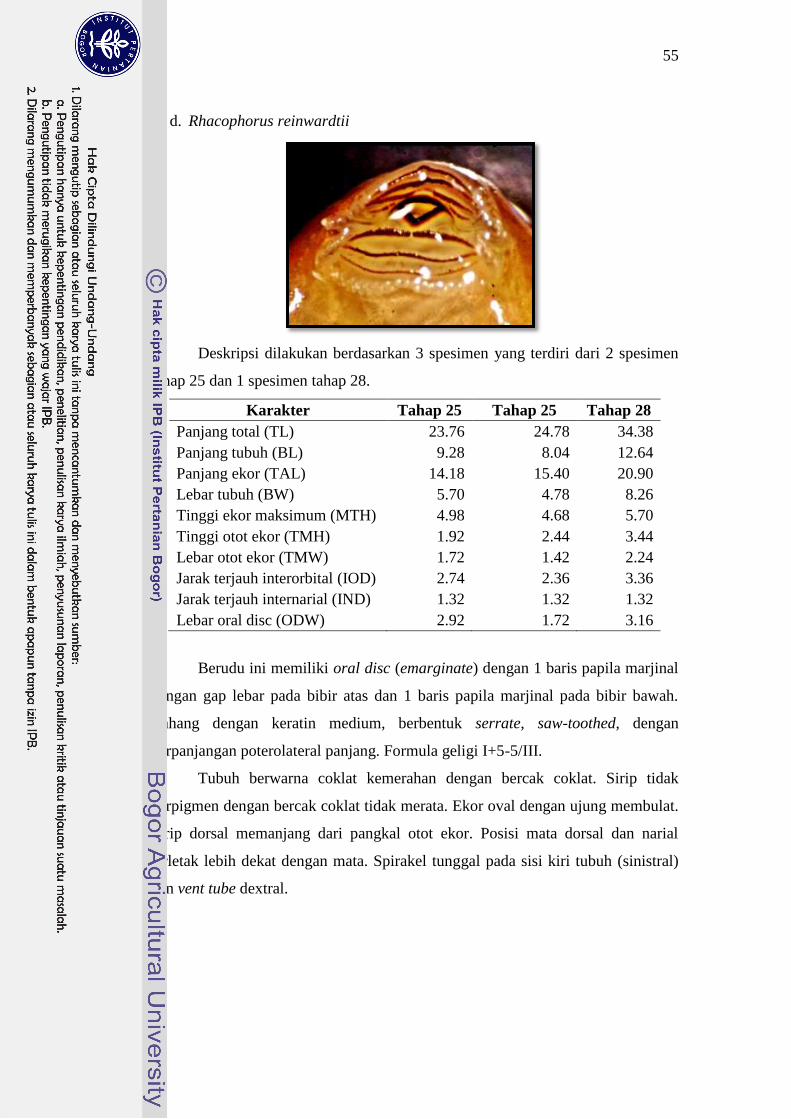
55
d. Rhacophorus reinwardtii
Deskripsi dilakukan berdasarkan 3 spesimen yang terdiri dari 2 spesimen
tahap 25 dan 1 spesimen tahap 28.
Karakter Tahap 25 Tahap 25 Tahap 28
Panjang total (TL) 23.76 24.78 34.38
Panjang tubuh (BL) 9.28 8.04 12.64
Panjang ekor (TAL) 14.18 15.40 20.90
Lebar tubuh (BW) 5.70 4.78 8.26
Tinggi ekor maksimum (MTH) 4.98 4.68 5.70
Tinggi otot ekor (TMH) 1.92 2.44 3.44
Lebar otot ekor (TMW) 1.72 1.42 2.24
Jarak terjauh interorbital (IOD) 2.74 2.36 3.36
Jarak terjauh internarial (IND) 1.32 1.32 1.32
Lebar oral disc (ODW) 2.92 1.72 3.16
Berudu ini memiliki oral disc (emarginate) dengan 1 baris papila marjinal
dengan gap lebar pada bibir atas dan 1 baris papila marjinal pada bibir bawah.
Rahang dengan keratin medium, berbentuk serrate, saw-toothed, dengan
perpanjangan poterolateral panjang. Formula geligi I+5-5/III.
Tubuh berwarna coklat kemerahan dengan bercak coklat. Sirip tidak
berpigmen dengan bercak coklat tidak merata. Ekor oval dengan ujung membulat.
Sirip dorsal memanjang dari pangkal otot ekor. Posisi mata dorsal dan narial
terletak lebih dekat dengan mata. Spirakel tunggal pada sisi kiri tubuh (sinistral)
dan vent tube dextral.




















