
Biokimi Dasar II Asam Lemak Kel 1
12
Pengaktifan Asam Lemak dan Oksidasi Asam Lemak di Mitokondria Biokimi Dasar II Kelompok 1 : 1. Delvan Armandela 2. Filo Ardiansyah 3. Hendru 4. Juliati Fitri 5. Rani Oktavia 6. Rika Mulia 7. Rolly Afrila 8. Septika Suryani 9. Lidya Dosen Pembimbing : Riryn Noviyanti MSi Jurusan Kimia Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Riau Pekanbaru 2014 Kelompok 1
-
Upload
rikamulia95 -
Category
Documents
-
view
53 -
download
2
description
jcjcjcjcj
Transcript of Biokimi Dasar II Asam Lemak Kel 1

Pengaktifan Asam Lemak dan Oksidasi Asam Lemak di Mitokondria
Biokimi Dasar II
Kelompok 1 :1. Delvan Armandela2. Filo Ardiansyah3. Hendru4. Juliati Fitri5. Rani Oktavia6. Rika Mulia7. Rolly Afrila8. Septika Suryani9. Lidya
Dosen Pembimbing : Riryn Noviyanti MSi
Jurusan KimiaFakultas Matematika dan Ilmu Pengetahuan Alam
Universitas RiauPekanbaru
2014Kelompok 1

OKSIDASI ASAM LEMAK
Asam lemak yang ada di dalam tubuh banyak mengalami oksidasi menjadi asetil KoA. Oksidasi
asam lemak ini terjadi di dalam mitokondria. Untuk memasuki mitokondria, asam-asam lemak
pertama-tama harus diubah menjadi suatu bentuk asil-KoA oleh aksi tiokinase dan ATP dalam
mikrosom atau pada permukaan mitokondria. Untuk asam-asam lemak rantai panjang,
biasanya harus diubah terlebih dahulu menjadi asilkarnitin supaya dapat masuk menembus
membran mitokondria. Sesampainya di dalam mitokondria, barulah asam lemak dapat
dioksidasi. Semua proses ini mulai dari masuk ke dalam mitokondria hingga mengalami oksidasi
terjadi dalam 3 tahap.
Sistem oksidasi pada asam lemak melibatkan 3 tahap, yaitu:1. Aktivasi asam lemak yang terjadi di sitoplasma2. Transport asam lemak ke dalam mitokondria3. Proses oksidasi di dalam matriks mitokondria
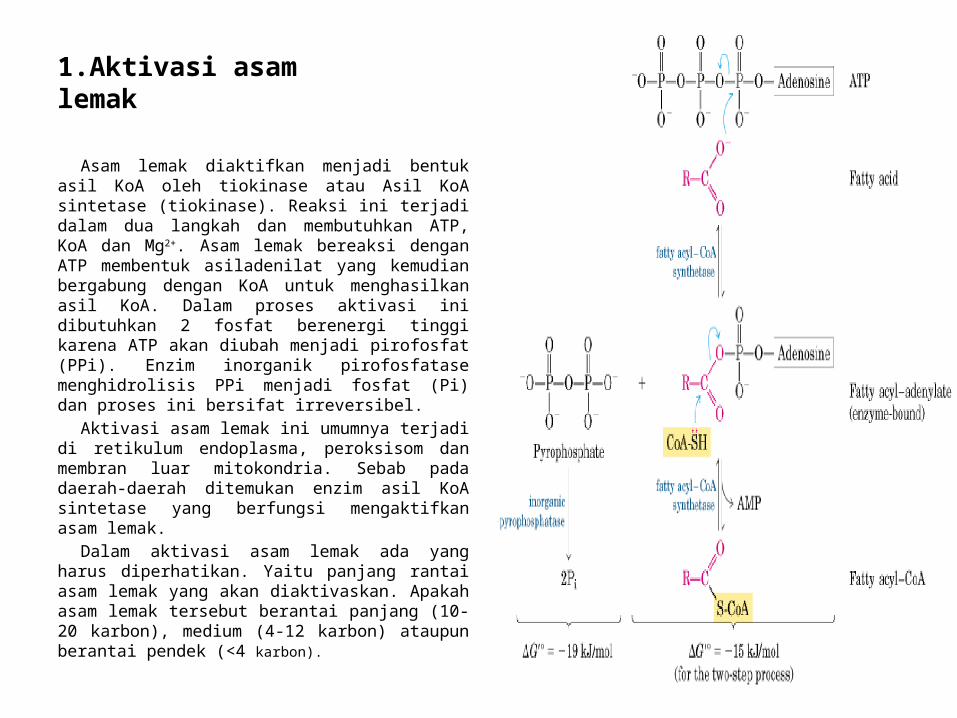
1.Aktivasi asam lemak
Asam lemak diaktifkan menjadi bentuk asil KoA oleh tiokinase atau Asil KoA sintetase (tiokinase). Reaksi ini terjadi dalam dua langkah dan membutuhkan ATP, KoA dan Mg2+. Asam lemak bereaksi dengan ATP membentuk asiladenilat yang kemudian bergabung dengan KoA untuk menghasilkan asil KoA. Dalam proses aktivasi ini dibutuhkan 2 fosfat berenergi tinggi karena ATP akan diubah menjadi pirofosfat (PPi). Enzim inorganik pirofosfatase menghidrolisis PPi menjadi fosfat (Pi) dan proses ini bersifat irreversibel.
Aktivasi asam lemak ini umumnya terjadi di retikulum endoplasma, peroksisom dan membran luar mitokondria. Sebab pada daerah-daerah ditemukan enzim asil KoA sintetase yang berfungsi mengaktifkan asam lemak.
Dalam aktivasi asam lemak ada yang harus diperhatikan. Yaitu panjang rantai asam lemak yang akan diaktivaskan. Apakah asam lemak tersebut berantai panjang (10-20 karbon), medium (4-12 karbon) ataupun berantai pendek (<4 karbon).

2. Transport asil KoA ke dalam mitokondria
Asil KoA (asam lemak yang sudah diaktifkan) yang berantai panjang tidak dapat menembus membran mitokondria dengan mudah. Oleh karena itu, Asil KoA berantai panjang akan diubah dulu menjadi asilkarnitin oleh karnitin parmitoiltransferase-I (karnitin asiltransferase-I) yang ada di sitoplasma agar dapat masuk ke dalam mitokondria.
Semua proses ini terjadi dalam 4 tahap1. Grup Asil pada asilKoA ditransfer ke karnitin yang dibantu oleh karnitin
asiltransferase-I. Pada tahap ini asilKoA akan diubah menjadi Asilkarnitin2. Asilkarnitin akan dibawa masuk menembus membran mitokondria
sampai ke matriks oleh protein carrier.3. Karnitin asiltransferase-II (yang berada di membran dalam mitokondria)
akan mengkonversi asilkarnitin menjadi asilKoA kembali sedangkan karnitin akan dilepas.
4. Karnitin akan dilepaskan ke sitoplasma untuk digunakan kembali.
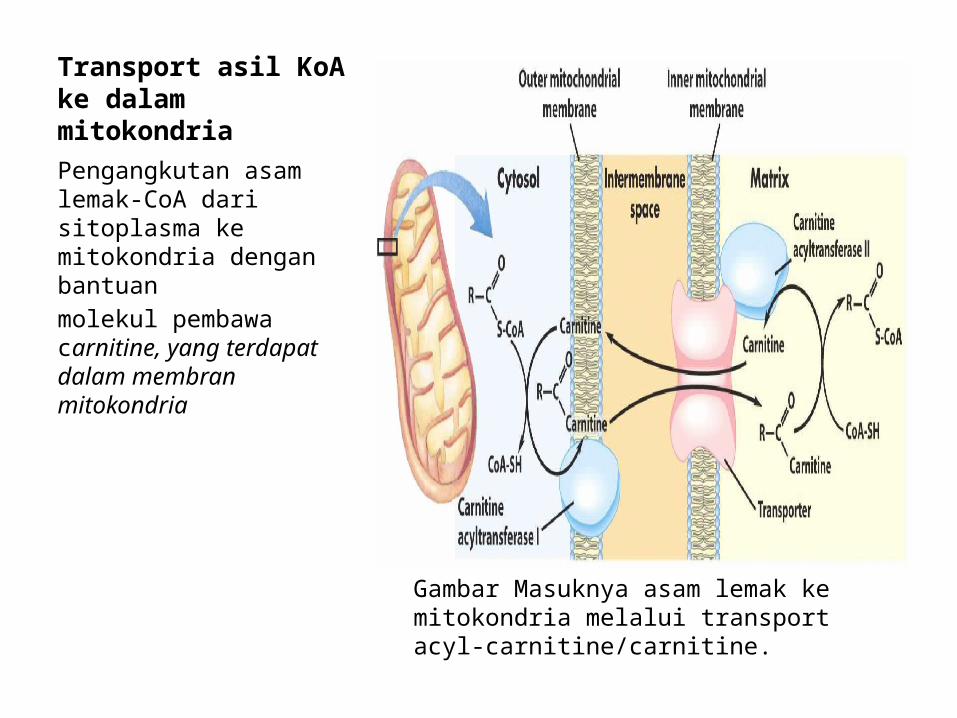
Transport asil KoA ke dalam mitokondriaPengangkutan asam lemak-CoA dari sitoplasma ke mitokondria dengan bantuanmolekul pembawa carnitine, yang terdapat dalam membran mitokondria
Gambar Masuknya asam lemak ke mitokondria melalui transport acyl-carnitine/carnitine.

3. Proses oksidasi di dalam matriks mitokondria
Setiap siklus oksidasi akan membebaskan dua unit karbon asetil KoA dan terjadi dalam 4 urutan reaksi.
1. Oksidasi: Asil KoA mengalami dehidrogenasi oleh FAD-dependent flavoenzim, asilKoA dehidrogenase. Ikatan doubledibentuk antara karbon ? dan ?.
2. Hidrasi: Enoil KoA hidratase menghidrasi ikatan double tadi sehingga membentuk ?-hidroksiasil KoA.
3. Oksidasi: Dalam tahap ini akan dihasilkan ?-ketoasil KoA.4. Pemecahan (Cleavage): Reaksi terakhir ?-oksidasi akan membebaskan 2 karbon asetil KoA
dari asil KoA.
Pada jalur oksidasi, asam lemak yang jumlah atom karbonnya ganjil, akan membentuk asetil KoA hingga tersisa sebuah residu tiga karbon (propionil KoA). Propionil KoA ini akan diubah menjadi suksinil KoA yang siap memasuki siklus asam sitrat.
Kesimpulan:Pada oksidasi asam lemak, asam lemak akan diubah dalam proses ?-oksidasi menjadi asetil KoA.

3. Proses oksidasi di dalam matriks mitokondria
1. Dehidrogenasi I, yaitu dehidrogenasi Asamlemak-CoA yang sudah berada di dalammitokondrion oleh enzim acyl-CoAdehidrogenase, mengha-silkan senyawaenoyl-CoA. Pada reaksi ini, FAD (flavinadenin dinukleotida) yang bertindaksebagai koenzim direduksi menjadiFADH2. Dengan mekanisme fosforilasibersifat oksidasi melalui rantai pemafasan,suatu molekul FADH2 dapat menghasilkandua molekul ATP.
2. Hidratasi, yaitu ikatan rangkap pada enoyl-CoA dihidratasi menjadi 3-hidroxyacyl-CoA oleh enzim enoyl-CoA hidratase.
3. Dehidrogenase II, yaitu dehidrogenasi 3-hidroxyacyl-CoA oleh enzim ß-hidroxyacyl-CoA dehidrogenase dengan NAD+sebagai koenzimnya menjadi ß-ketoacyl-CoA. NADH yang terbentuk dari NAD+dapat dioksidasi kembali melaluimekanisme fosforilasi oksidatif yangdirangkaikan dengan rantai pernafasanmenghasilkan tiga molekul ATP.
4. Pemecahan molekul dengan enzim ß-ketoacyl-CoA thiolase. Pada reaksi ini satu molekulketoacyl-CoA menghasilkan satu molekul asetyl-CoA dan sisa rantai asam lemak dalambentuk CoA-nya, yang mempunyai rantai dua atom karbon lebih pendek dari semula.

JALUR MINOR DEGRADASI ASAM LEMAK
Jalur utama degradasi asam lemak adalah ß-oksidasi, yaitu untuk asam lemak jenuh beratom C genap. Akan tetapi ada juga jalur-jalur khusus yang lain yaitu untuk degradasi asam lemak tak jenuh, degradasi asam lemak dengan atom C ganjil, serta α oksidasi dan Ѡ-oksidasi.
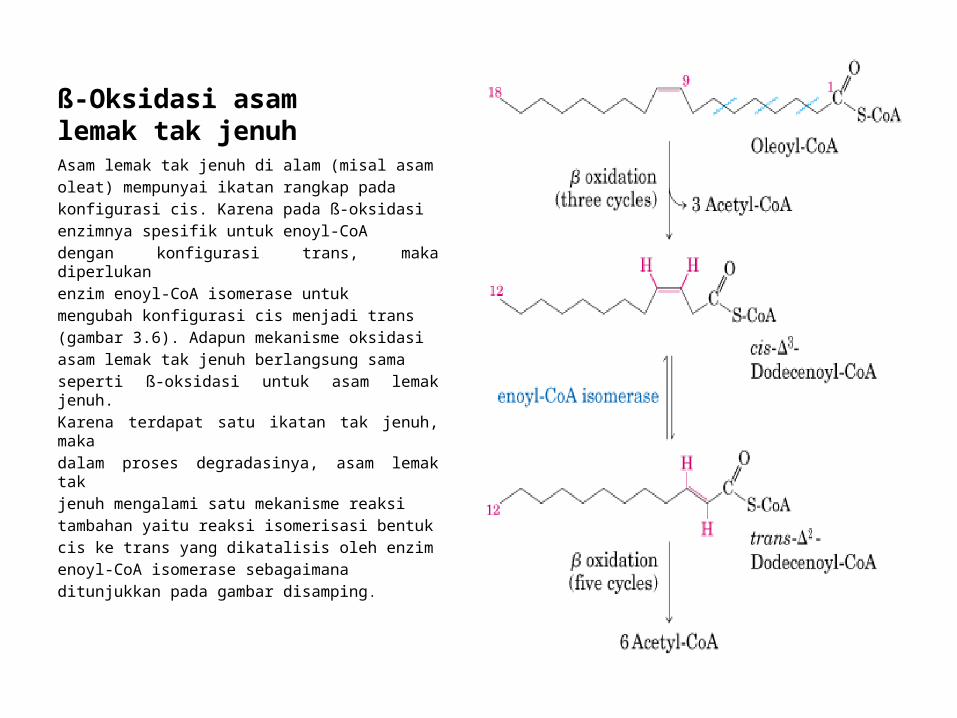
ß-Oksidasi asam lemak tak jenuhAsam lemak tak jenuh di alam (misal asamoleat) mempunyai ikatan rangkap padakonfigurasi cis. Karena pada ß-oksidasienzimnya spesifik untuk enoyl-CoAdengan konfigurasi trans, maka diperlukanenzim enoyl-CoA isomerase untukmengubah konfigurasi cis menjadi trans(gambar 3.6). Adapun mekanisme oksidasiasam lemak tak jenuh berlangsung samaseperti ß-oksidasi untuk asam lemak jenuh.Karena terdapat satu ikatan tak jenuh, makadalam proses degradasinya, asam lemak takjenuh mengalami satu mekanisme reaksitambahan yaitu reaksi isomerisasi bentukcis ke trans yang dikatalisis oleh enzimenoyl-CoA isomerase sebagaimanaditunjukkan pada gambar disamping.
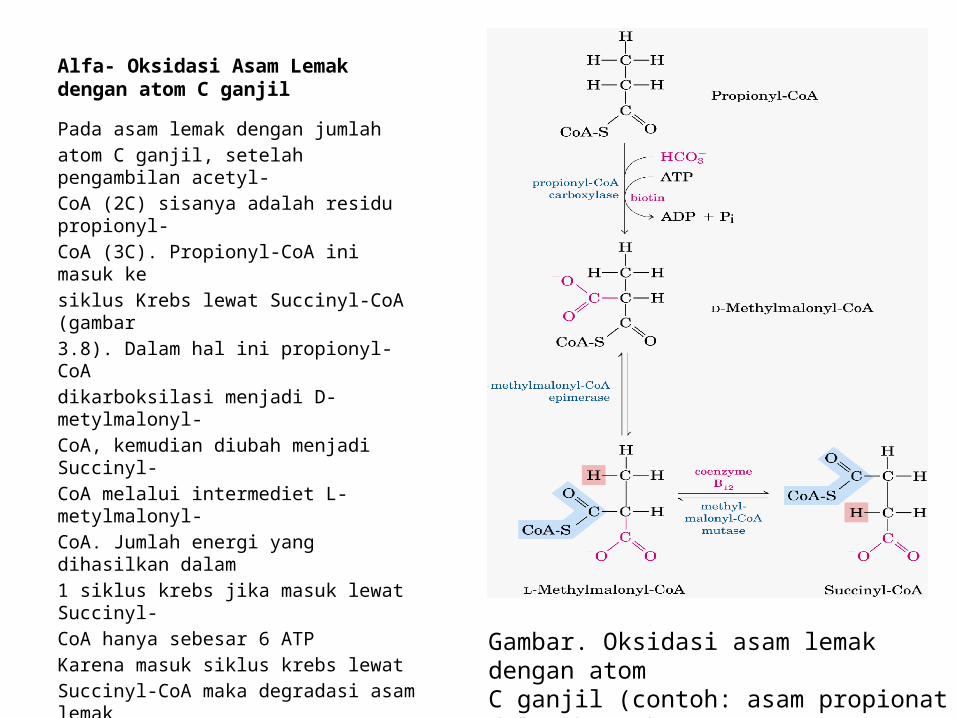
Alfa- Oksidasi Asam Lemak dengan atom C ganjil
Pada asam lemak dengan jumlahatom C ganjil, setelah pengambilan acetyl-CoA (2C) sisanya adalah residu propionyl-CoA (3C). Propionyl-CoA ini masuk kesiklus Krebs lewat Succinyl-CoA (gambar3.8). Dalam hal ini propionyl-CoAdikarboksilasi menjadi D-metylmalonyl-CoA, kemudian diubah menjadi Succinyl-CoA melalui intermediet L- metylmalonyl-CoA. Jumlah energi yang dihasilkan dalam1 siklus krebs jika masuk lewat Succinyl-CoA hanya sebesar 6 ATPKarena masuk siklus krebs lewatSuccinyl-CoA maka degradasi asam lemakdengan atom C ganjil lebih cepatdibandingkan dengan degradasi asamlemak dengan atom C genap. Hal inipenting untuk memberikan konsumyi padaorang atau makhluk hidup yangmembutuhkan energi dengan cepat, misalorang Eskimo. Gambar. Oksidasi asam lemak dengan atom
C ganjil (contoh: asam propionat dalam bentukPropionyl-CoA)

KESIMPULAN
Asam lemak jenuh didegedasi dalam 3 tahapan oksidasi. Tahap pertama, ß-oksidasi,dilakukan dalam siklus yang berkesinambungan dengan hasil akhir sebagai acetyl-CoA. Tiap siklus terdiri atas 4 tahap reaksi, yaitu (1) dehidrogenasi 1, (2) hidratasi, (3) dehidroenasi 2,dan (4) tiolasi. Pada tahap kedua tiap acetyl-CoA dioksidasi menghasilkan 2 CO2 dan 8 elektron dalam siklus TCA. Pada tahap ketiga, elektron yang dihasilkan dari tahap 1 dan 2 masuk ke rantai respirasi mitokondria dengan menghasilkan energi untuk sintesis ATP dengan forforilasi oksidatif. Oksidasi asam lemak tidak jenuh memerlukan 2 enzim tambahan: enoyl-CoA isomerase dan 2,4-dienoyl-CoA reductase. Asam lemak beratom C ganjil dioksidasi ß menghasilkan acetyl-CoA dan propionyl-CoA. Propionyl-CoA dikarboksilasi menjadi Lmethylmalonyl- CoA yang kemudian diisomerisasi menjadi succinyl-CoA untuk dioksidasi menjadi CO2 dalam siklus TCA.





















