bioenergetika
12
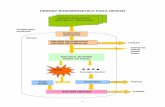
6. BIOENERGITIKA MIKROORGANISME METABOLISME : ENERGI DAN ENZIM 1. Energi dan Kerja a. Energy kemampuan untuk melakukan kerja b. Sel hidup melakukan 3 tipe utama kerja i. Kerja Kimia—mensintesis molekul kompleks ii. Kerja transport pengambilan nutrien, eliminasi sisa, keseimbangan ion iii. Kerja Mekanik internal dan perpindahan eksternal c. Pada ekosistem, photoautotrophs dan chemolithoautotrophs menangkap energi dan menggunakan beberapa untuk transformasi karbondioksida menjadi molekul-molekul organik; molekul organik kemudian menyediakan sumber karbon dan energi untuk chemoheterotrophs, yang mengubah oksidasi molekul organik melalui proses seperti respirasi aerob, melepaskan karbondioksida d. Sumber energi utama pada sel hidup adalah adenosine-5'- triphosphate (ATP) 2. Hukum termodinamika a. Ilmu analisis perubahan energi termodinamika pada pengumpulan materi disebut sistem; semua materi lain di alam disebut lingkungan sekeliling b. Hukum pertama, energi tidak dapat diciptakan dan dimusnahkan i. Total energi di alam cenderung konstan ii. Energi dapat diredistribusikan baik itu dalam sistem atau antara sistem dan lingkungan sekelilingnya iii. Energi diukur dalam kalori dimana 1 kalori adalah sejumlah energi panas yang dibutuhkan untuk meningkatkan suhu 1 gram air dari 14.5°C hingga 15.5°C c. Hukum kedua, proses fisika dan kimia berlangsung dalam beberapa cara dimana kesalahan dalam peningkatan alam semesta hingga kemungkinan maksimum 3. Energi bebas dan reaksi a. Perubahan energi yang dapat terjadi pada reaksi kimia diekpresikan dalam persamaan untuk perubahan energi (DG = DH – T/DS); perubahan energi bebas (DG) adalah sejumlah energi pada sistem yang tersedia untuk melakukan kerja b. Perubahan energi bebas pada reaksi kimia secara langsung berhubungan dengan keseimbangan konstan reaksi i. Standar perubahan energi bebas (DG0´) adalah perubahan dalam energi bebas dibawah kondisi standar konsentrasi, pH, tekanan, dan temperatur ii. Ketika DG0´ negatif, keseimbangan konstan lebih besar daripada satu dan reaksi berlangsung ke penyelesaian seperti yang tertulis; reaksi dikatakan exergonic iii. Ketika DG0´ positif, keseimbangan konstan kurang dari satu dan sedikit produk dibentuk pada saat seimbang; reaksi dikatakan endergonic
-
Upload
lialestari -
Category
Documents
-
view
110 -
download
1
description
bioenergetik
Transcript of bioenergetika

6. BIOENERGITIKA MIKROORGANISME
METABOLISME : ENERGI DAN ENZIM
1. Energi dan Kerjaa. Energy kemampuan untuk melakukan kerjab. Sel hidup melakukan 3 tipe utama kerja
i. Kerja Kimia—mensintesis molekul kompleks ii. Kerja transport pengambilan nutrien, eliminasi sisa, keseimbangan ion
iii. Kerja Mekanik internal dan perpindahan eksternal c. Pada ekosistem, photoautotrophs dan chemolithoautotrophs menangkap energi dan
menggunakan beberapa untuk transformasi karbondioksida menjadi molekul-molekul organik; molekul organik kemudian menyediakan sumber karbon dan energi untuk chemoheterotrophs, yang mengubah oksidasi molekul organik melalui proses seperti respirasi aerob, melepaskan karbondioksida
d. Sumber energi utama pada sel hidup adalah adenosine-5'-triphosphate (ATP)2. Hukum termodinamika
a. Ilmu analisis perubahan energi termodinamika pada pengumpulan materi disebut sistem; semua materi lain di alam disebut lingkungan sekeliling
b. Hukum pertama, energi tidak dapat diciptakan dan dimusnahkani. Total energi di alam cenderung konstan
ii. Energi dapat diredistribusikan baik itu dalam sistem atau antara sistem dan lingkungan sekelilingnya
iii. Energi diukur dalam kalori dimana 1 kalori adalah sejumlah energi panas yang dibutuhkan untuk meningkatkan suhu 1 gram air dari 14.5°C hingga 15.5°C
c. Hukum kedua, proses fisika dan kimia berlangsung dalam beberapa cara dimana kesalahan dalam peningkatan alam semesta hingga kemungkinan maksimum
3. Energi bebas dan reaksia. Perubahan energi yang dapat terjadi pada reaksi kimia diekpresikan dalam persamaan untuk
perubahan energi (DG = DH – T/DS); perubahan energi bebas (DG) adalah sejumlah energi pada sistem yang tersedia untuk melakukan kerja
b. Perubahan energi bebas pada reaksi kimia secara langsung berhubungan dengan keseimbangan konstan reaksi
i. Standar perubahan energi bebas (DG0´) adalah perubahan dalam energi bebas dibawah kondisi standar konsentrasi, pH, tekanan, dan temperatur
ii. Ketika DG0´ negatif, keseimbangan konstan lebih besar daripada satu dan reaksi berlangsung ke penyelesaian seperti yang tertulis; reaksi dikatakan exergonic
iii. Ketika DG0´ positif, keseimbangan konstan kurang dari satu dan sedikit produk dibentuk pada saat seimbang; reaksi dikatakan endergonic
4. Peran ATP pada Metabolismea. ATP adalah molekul berenergi tinggi; penyingkiran fosfat terminal melalui hidrolisis
berlangsung hampir selesai denga perubahan besar energi bebas standar negatif (yaitu, reaksi sangat exergonic); ATP juga memiliki potensi transfer grup fosfat tinggi
b. Karakteristik tersebut membuat ATP sesuai dalam peranannya sebagai sumber energi; ATP dibentuk dari ADP dan Pi melalui proses penangkapan energi; exergonic memecahkan ATP dipasangkan dengan macam-macam reaksi endergonik untuk memfasilitasi penyelesaian mereka
5. Reaksi Oksidasi-Reduksi dan Pembawa Elektrona. Pelepasan energi selama proses metabolik berlangsung secara normal melibatkan reaksi oksidasi
dan reduksii. Reaksi Oxidasi-reduksi (redox) melibatkan transfer elektron dari donor (agen pereduksi
atau reductant) ke akseptor (agen pengoksidasi atau oxidant) ii. Keseimbangan konstan untuk reaksi oksidasi-reduksi disebut Standar potensial reduksi
(E0) dan mengukur kecenderungan agen pereduksi untuk kehilangan elektron; potensial reduksi lebih negatif, agen pereduksi yang lebih baik adalah sebagai donor elektron
b. Ketika elektron ditransfer dari donor elektron ke akseptor elektron dengan potensial reduksi lebih positif, energi bebas dilepaskan dan dapat digunakan dari ATP
c. Transport Electron penting dalam berbagai proses metabolik (seperti, respirasi dan photosynthesis); sel-sel menggunakan bermacam-macam pemabwa elektron yang diorganisasi dalam rantai untuk memindahkan elektron; pembawa electron membawa antara lain NAD+,

NADP+, flavoproteins, coenzymes, dan cytochromes; pembewa tersebut berbeda dalam hal bagaimana mereka membawa elektron, dan ini berpengaruh kuat dengan bagaimana mereka berfungsi pada rantai transport elektron
6. Enzima. Struktur dan klasifikasi enzim
i. Enzim adalah protein katalis yang sangat spesifik untuk reaksi terkatalisis dan molekul dalam bereaksi
Katalis adalah substansi yang meningkatkan kecepatan reaksi tanpa perubahan secara permanen
Molekul yang bereaksi disebut substrat dan substansi yang dibentuk adalah produk
ii. Enzim dapat hanya mengandung protein atau dapat berupa holoenzyme, terdiri dari komponen protein (apoenzyme) dan komponen nonprotein (cofactor)
Kofaktor Grup Prosthetic yang benar-benar diserang apoenzim Kofaktor Coenzyme yang sedikit diserang apoenzim; ini mungkin memisahkan
dari apoenzim dan membawa satu atau lebih produk reaksi ke enzim lainb. Mekanisme reaksi enzim
i. Enzim meningkatkan kecepatan reaksi, namun tidak merubah keseimbangan konstanii. Enzim menurunkan energi aktivasi yang dibutuhkan untuk membawa bersama molekul
tereaksi dengan benar untuk membentuk komplek keadaan transisi; ketika keadaan transisi telah dicapai reaksi berlangsung secara cepat
iii. Enzim membawa bersama substrat pada sisi aktif untuk membentuk komplek substrat enzim; hal ini dapat menurunkan energi aktivasi dalam beberapa cara:
Konsentrasi lokal substrat ditingkatkan pada sisi aktif (catalytic) enzim Molekul pada sisi aktif diorientasikan secara benar supaya reaksi dapat terjadi
c. Pengaruh lingkungan terhadap aktivitas enzim i. Adanya Sejumlah substrat dapat mempengaruhi kecepatan reaksi, dimana peningkatan
seperti peningkatan konsentrasi substrat hingga semua molekul enzim dapat mengikat substrat dan mengubahnya menjadi produk secepat mungkin
Tidak ada peningkatan kecepatan yang akan terjadi dengan peningkatan subsekuen pada konsentrasi substrat, dan reaksi tersebut berlangsung pada kecepatan maksimal (Vmax)
Konstanta The Michaelis (Km) dari enzim adalah konsentrasi substrat yang dibutuhkan untuk reaksi mencapai setengah kecepatan maksimal dan digunakan seperti mengukur afinitas lawan dari enzim untuk substrat tersebut
ii. Aktivitias Enzim dipengaruhi oleh perubahan pH dan temperatur; setiap enzim memiliki pH and temperature optimum spesifik; keadaan ekstrim pH, temperature, dan faktor lain dapat menyebabkan denaturasi (kehilangan aktivitas berhubungan dengan gangguan struktur)
d. Penghambatan Enzim i. Penghambatan kompetitif terjadi ketika inhibitor mengikat sisi aktif dan biasanya
bersaing dengan substrat (jika inhibitor mengikat, kemudian substrat tidak dapat, dan reaksi tidak terjadi); tipe penghambatan ini dapat diatasi dengan penambahan substrat berlebihan
ii. Penghambatan Noncompetitive terjadi ketika inhibitor mengikat enzim pada beberapa lokasi selain pada sisi aktif dan merubah bentuk enzim sehingga ini tidak aktif atau kurang aktif; tipe penghambatan ini tidak dapat diatasi dengan penambahan substrat berlebihan
7. Alamiah dan Signifikan Regulasi Metabolika. Regulasi penting bagi microorganisme untuk pemenuhan energi dan materi dan untuk mengatur
keseimbangan metabolik meskipun sering berubah pada lingkungan merekab. Proses metabolik dapat diregulasi menjadi tiga cara utama:
i. Saluran Metabolik melokalisasi metabolit dan enzim pada bagian berbeda di selii. Stimulus atau penghambatan enzim pada jalur
iii. Peningkatan atau penurunan jumlah molekul enzim yang ada (regulasi ekspresi gen)8. Penyaluran Metabolik
a. Cara pembagian ruangan adalah mekanisme biasa pada penyaluran metabolik; enzim dan metabolit didistribusikan pada struktur sel atau organel terpisah
b. Penyaluran dapat menghasilkan variasi tertanda pada konsentrasi metabolit dan oleh karena itu secara langsung mempengaruhi aktivitas enzim

9. Kontrol aktivitas Enzima. Regulasi Allosteric dari aktivitas enzim oleh efektor atau modulator, yang mengikat dapat balik
dan tidak kovalen pada sisi regulator enzim; sisi regulator berbeda dengan sisi katalitikb. Modifikasi kovalent dari regulasi enzim aktivitas enzim oleh penambahan kovalen atau
pemindahan grup kimia (seperti, phosphate, grup methyl, asam adenylic) c. Penghambatan Balik
i. Setiap jalur memiliki sedikitnya enzim pacemaker yang mengkatalisis terlambat (terbatasnya kecepatan) reaksi pada jalur; seringkali ini sebagai reaksi pertama pada jalur
ii. Pada penghambatan balik (penghambatan produk akhir), produk akhir dari jalur menghambat enzim pacemaker
iii. Pada jalur bercabang, keseimbangan antara produk akhir diatur melalui penggunaan enzim regulator pada titik cabang; jalur bercabang multipel seringkali menggunakan isoenzim, masing-masing dibawah kontrol terpisah dan bebas
METABOLISME : PEMBENTUKAN ENERGI
1. Ringkasan Metabolismea. Metabolisme (semua reaksi kimia yang terjadi di dalam sel) dapat dibagi menjadi dua bagian
utama: katabolisme dan anabolismei. Katabolisme-pemecahan molekul besar dan komplek menjadi molekul yang lebih kecil
serta sederhana, selama proses ini energi dilepaskan, dijebak, dan dapat digunakan untuk melakukan kerja
ii. Anabolisme-sintesis molekul komplek dari yang sederhana dan selama proses ini energi ditambahkan sebagai input
iii. Chemolithotrophy dan fotosintesis termasuk sebagai proses penghasilan energi katabolik, meskipun mereka tidak melibatkan degradasi molekul komplek
b. Mikroorganisme khemotrophic tidak hanya bervariasi dalam sumber energi mereka namun juga akseptor elektron mereka
i. Jika sumber energi dioksidasi dan didegradasi tanpa menggunakan akseptor elektron eksogen, proses disebut fermentasi
ii. Jika sumber energi dioksidasi dan didegradasi menggunakan akseptor elektron eksogen, proses disebut respirasi; pada respirasi aerob akseptor elektron terakhir adalah oksigen, sedangkan pada respirasi anaerob akseptor elektron adalah molekul lain selain oksigen
c. Untuk organisme chemoorganoheterotrophic, katabolisme seringkali proses tiga tahap selama nutrien memasuki jalur degradasi; biasanya jalur fungsi secara katabolik dan anabolik dinamakan ampibolik
2. Pemecahan Glukosa menjadi Piruvata. Jalur glikolisis
i. Juga dikenal sebagai jalur Embden-Meyerhof, ini jalur yang paling biasa ditemukan di semua grup utama mikroorganisme
ii. Fungsi ini berlangsung ada atau tidak ada oksigen dan dibagi menjadi dua bagian: Tahap glukosa gula 6-carbon difosforilasi dua kali menghasilkan fruktosa 1,6-
bisphosphate; ini membutuhkan pengeluaran dua molekul ATP Tahap gula 3 karbon memecah fructose 1,6-bisphosphate menjadi dua
molekul 3 karbon, dimana masing-masing diproses menjadi piruvat; dua molekul ATP diproduksi oleh fosforilasi tingkat substrat dari masing-masing molekul 3 karbon menghasilkan dua molekul ATP; 2 molekul NADH juga diproduksi per molekul glukosa
b. Jalur pentosa fosfat i. Juga dikenal sebagai jalur hexose monophosphate, jalur ini menggunakan kumpulan
reaksi berbeda untuk memproduksi bermacam-macam gula fosfat 3-, 4-, 5-, 6-, dan 7-karbon
ii. Memiliki beberapa fungsi katabolik dan anabolik Produksi NADPH, dimana bertindak sebagai sumber elektron untuk proses
biosintesis Sumber jaringan empat dan lima karbon yang dapat digunakan dalam sintesis
asam amino, asam nukleat, dan makromolekul lain Katabolisme lengkap dari heksosa dan pentosa, menghasilkan ATP dan NADH
(dibuat dengan mengubah NADPH menjadi NADH)

c. Jalur Entner-Doudoroff i. Hubungan Reaksi jalur ini yaitu jalur pentosa fosfat dengan jalur glikolisis dengan
reaksi unikii. Jalur ini memproduksi ATP, NADPH, dan NADH
3. Fermentasia. Fermentasi-proses dimana organisme mengoksidasi NADH yang diproduksi melalui salah satu
jalur di atas dengan menggunakan piruvat atau satu derivatnya sebagai akseptor elektron dan hidrogen, jadi proses ini melibatkan pengggunaan akseptor elektron endogen
b. Banyak tipe fermentasi diketahui i. Fermentasi alkohol memproduksi entanol dan CO2
ii. Fermentasi Asam laktat memproduksi asam laktat (lactate) Fermenter Homolaktik mereduksi hampir semua piruvat menjadi laktat Fermenter Heterolaktik membentuk sejumlah substansi produk selain laktat
iii. Fermentasi asam format memproduksi baik itu asam campuran atau butanediolc. Microorganisme dapat memfermentasi bahan lain selain gula (misalnya, asam amino)
4. Siklus Asam Tricarboxylica. Pyruvat dapat didegradasi menjadi karbondioksida melalui siklus asam tricarboxylic (TCA)
sesudah pertama telah dikonversikan menjadi acetyl CoA; reaksi ini dibarengi dengan hilangnya satu atom karbon seperti karbon dioksida
b. Acetyl-CoA bereaksi dengan oxaloacetate (molekul 4-carbon) untuk memproduksi molekul 6 karbon, yang secara subsekuen dipecah menjadi dua molekul karbondioksida, perbaruan oksaloasetat; selama proses ini, terjadi hal-hal berikut:
i. ATP diproduksi melalui fosforilasi tingkat substratii. Tiga molekul NADH dan satu molekul FADH2 diproduksi
c. Meskipun organisme kurang lengkap siklus TCAnya biasanya memiliki banyak enzim siklus karena salah satu fungsi utama siklus TCA adalah menyediakan jaringan karbon untuk digunakan pada biosintesis
5. Transport Elektron dan Fosforilasi Oksidatifa. Rantai Transport Electron
i. Rantai transport elektron mitokondria menggunakan rangkaian pembawa elektron untuk mentransfer elektron dari NADH dan FADH2 ke O2
Pembawa Electron berlokasi didalam membran dalam mitokondria Selama fosforilasi oksidatif, tiga molekul ATP dapat disintesis ketika sepasang
elektron dilewatkan dari NADH ke O2; dua molekul ATP dapat disintesis ketika elektron dari FADH2 lewat ke O2
ii. Meskipun mereka beroperasi berdasarkan pronsip dasar yang sama, rantai transport elektron bakteri biasanya berbeda pada strukturnya: mereka mungkin bercabang, dapat dibentuk dari pembawa elektron berbeda, atau dapat lebih pendek daripada rantai transport elektron mitokondria; rentai transport elektron berlokasi di membran plasma
b. Oxidative Phosphorylation i. The chemiosmotic hypothesis of oxidative phosphorylation postulates that the energy
released during electron transport is used to establish a protonmotive force (potential energy due to the difference in proton concentration and charge on either side of the membrane), which can be used to drive ATP synthesis, flagellar rotation, and transport of molecules across the membrane
ii. ATP synthesis is catalyzed by the ATP synthase complex, which is thought to behave like a small rotary motor
iii. Inhibitors of ATP synthesis fall into two main categories: Blockers that inhibit the flow of electrons through the system Uncouplers that allow electron flow, but disconnect it from oxidative
phosphorylationc. Produksi ATP melalui Glikolisis dan Respirasi Aerob
i. Produksi ATP melalui glikolisis selama fermentasi adalah 2 ATP ii. Respiraso Aerob menghasilkan antara 2 hingga 38 ATP per molekul glukosa,
tergantung pada ketepatan alamiah sistem transport elektroniii. Efek pasteur adalah fenomena regulator dimana organisme mengurangi kecepatan
katabolisme gula ketika kondisi menyebabkan pergantian dari fermentasi menjadi respirasi aerob; ini terjadi karena respirasi aerob lebih effisien dan menghasilkan energi lebih besar per molekul glukosa
6. Respirasi Anaerob

a. Menggunakan molekul selain oksigen sebagai akseptor elektron terakhir; yang paling biasa digunakan sebagai akseptor elektron alternatif adalah nitrat, sulfat, dan CO2
b. Reduksi nitrat Dissimilato terjadi ketika nitrat digunakan sebagai akseptor elektron terakhir; jika nitrat direduksi menjadi gas nitrogen, proses dinamakan denitrifikasi
c. Respirasi Anaerob tidak seefisien respirasi aerob dalam sintesis ATP karena akseptor elektron alternatif tidak memiliki potensial reduksi sepositif O2; meskipun demikian respirasi anaerob berguna karena lebih effisien daripada fermentasi
7. Katabolisme Karbohidrat dan Polimer Cadangan Intraselulera. Karbohidrat
i. Kebanyakan monosakarida masuk dengan mudah ke dalam jalur glikolisisii. Disaccharides dipecah menjadi monosakarida baik itu melalui hidrolisis atau
fosforolisisiii. Polysaccharida dipecah menjadi molekul lebih kecil baik itu melalui hidrolisis atau
fosforolisis; bagaimanapun kebanyakan tidak mudah didegradasi (misal, sellulosa, agar) iv. Microorganisme juga mampu mendegradasi molekul xenobiotic (bahan asing yang
tidak dibentuk melalui proses alamiah biosintesis) seperti pestisidab. Polimer Cadangan-ketika nutrien tidak ada mikroorganisme mengkatabolis simpanan glikogen,
pati, dll8. Katabolisme Lipid
a. Triglycerida biasanya merupakan sumber energi; mereka dihidrolisis menjadi gliserol dan asam lemak
b. Asam Lemak dikatalisis melalui jalur Oksidasi, yang memproduksi acetyl-CoA, NADH, dan FADH2; NADH dan FADH2 dapat dioksidasi melalui ranta transport elektron untuk memproduksi ATP
9. Katabolisme Protein dan Asam Amino a. Protein didegradasi oleh protease menjadi komponen asam amino merekab. Asam Amino pertama dideaminasi dan kemudian sisa jaringan karbon dikonversi menjadi
piruvat, acetyl-CoA, atau siklus intermediet TCA10. Oksidasi Molekul Anorganik
a. Chemolithotrophy – proses metabolik yang menggunakan molekul anorganik sebagai sumber energi,
i. Sumber energi dioksidasi; ii. Akseptor elektron biasanya O2, namun sulfat dan nitrat juga digunakan;
iii. Donor elektron (sumber energi) yang paling umum adalah hydrogen, komponen nitrogen tereduksi, komponen sulfur tereduksi, dan besi ferro (Fe2+)
b. Chemolithotrophs biasanya autotrophs; mereka menggunakan siklus Calvin untuk memperbarui karbondioksida
11. Fotosintesisa. Selama fotosintesis, energi dari cahaya matahari ditangkap dan digunakan untuk produksi ATP
dan NADPH (reaksi terang), yang digunakan untuk mereduksi karbondioksida membentuk karbohidrat (reaksi gelap)
b. Reaksi Terang eukariot dan cyanobakteriI. Molekul Chlorophyll dan bermacam-macam pigmen pelengkap digunakan untuk
membentuk anntena; antennas menjebak photons dan mentransfer mereka ke pusat reaksi klorofil; bagian klorofil khusus ini secara langsung terlibat pada transport elektron
II. Eucaryotes dan cyanobacteria memiliki dua photosystems; masing-masing, elektron dari pusat reaksi cahaya berenergi ditransfer ke rantai transport elektron gabungan
Photosystem I dapat melakukan fotofosforilasi siklik, memproduksi ATP Photosystems I dan II, bekerja bersama, dapat melakukan fosforilasi nonsiklik,
memproduksi ATP dan NADPH; elektron untuk fosforilasi nonsiklik didapatkan dari air, yang dioksidasi menjadi O2 (fotosintesis oksigenik)
c. Reaksi Terang bakteri hijau dan ungu i. Bakteri hijau dan ungu dapat melakukan fotosintesis anoksigenik (mereka tidak
menggunakan air sebagai sumber elektron, sehingga tidak memproduksi O2), dan mereka memiliki pigmen fotosintesis yang berbeda disebut bacteriochlorophylls
ii. Banyak perbedaan dalam reaksi terang bakteri hijau dan ungu disebabkan mereka hanya memiliki satu fotosistem
iii. Bakteri hijau dan Ungu biasanya autotrophs yang menggunakan NADH atau NADPH untuk fiksasi karbon dioksida; tiga metode untuk membuat NADH:

Reduksi NAD+ secara langsung oleh gas hidrogen Membalikkan aliran elektron Bentuk sederhana aliran elektron nonsiklik
METABOLISME : PENGGUNAAN ENERGI
1. Pendahuluana. ANABOLISME – pembentukan melalui sintesis molekul komplek dari yang sederhana; ini
membutuhkan input energib. PEMBALIKAN – degradasi secara terus menerus dan resintesis bahan selulerc. KECEPATAN BIOSINTESIS adalah Perkiraan Diseimbangkan dengan katabolisme,
disebabkan regulasi hati-hati dari proses metabolik2. Prinsip-Prinsip Pengaturan Biosintesis
a. Metabolisme biosintesis berlangsung berdasarkan beberapa prinsip berikut: i. Sintesis molekul besar komplek (makromolekul) dari sejumlah unit struktur sederhana
(monomers) yang terbatas menyimpan melebih kapasitas penyimpanan genetik, materi dasar biosintesis, dan energi
ii. Penggunaan banyak enzim yang sama untuk katabolisme dan anabolisme menyimpan penambahan materi dan energi
iii. Banyak jalur biosintesis merupakan kebalikan jalur katabolik; banyak langkah jalur dikatalisis oleh enzim yang berpartisipasi baik aktivitas katabolik maupun anabolik; bagaimanapun, beberapa langkah dikatalisis oleh dua enzim berbeda: satu berfungsi pada pengaturan katabolik dan kedua yang berfungsi pada pengaturan biosintesis; ini memperbolehkan regulasi bebas katabolisme dan anabolisme
iv. Rangkaian reaksi biosintesis dengan pemecahan ATP (atau nukleosida trifosfat lain) mengatur jalur anabolik tidak dapat balik dalam pengaturan biosintesis
v. Pada sel eucariot, reaksi anabolik dan katabolic melibatkan komponen yang sama secara bertahap dilokasikan pada kompartemen terpisah secara simultan namun operasi bebas
vi. Jalur Katabolik dan anaboli mengunakan kofaktor berbeda: katabolik oksidasi memproduksi NADH, dimana substrat untuk transport elektron , sementara NADPH bertindak sebagai reduktan untuk jalur anabolik
b. Ketika makromolekul telah dibentuk dari prekusor sederhana, struktur sel (misalnya, ribosom) membentuk secara spontan dari makromolekul melalui proses yang dikenal sebagai SELF-ASSEMBLY (PENGGABUNGAN SENDIRI)
3. Fiksasi Karbondioksida Fotosintesisa. Tiga Proses berbeda untuk perubahan karbondioksida menjadi karbon organik dikenal
i. Siklus Calvin (siklus reduksi pentosa fosfat)-ditemui pada fotosintesis eukariot dan banyak bakteri fotosintetik
ii. Siklus Reduksi TCA –digunakan oleh beberapa archaea dan bakteri iii. Jalur Acetyl-CoA –digunakan oleh methanogens, sulfate reducers, dan acetogens
b. Siklus Kalvini. Terdiri dari tiga fase
a. Fase carboksilasi - enzim ribulose 1,5-bisphosphate carboxylase mengkatalisis penambahan carbon dioxida menjadi ribulose 1,5-bisphosphate, membentuk dua molekul 3-phosphoglycerate
b. The reduction phase - 3-phosphoglycerate is reduced to glyceraldehyde 3-phosphate
c. The regeneration phase - a series of reactions is used to regenerate ribulose 1,5-bisphosphate and to produce carbohydrates such as fructose and glucose; this phase is similar to the pentose phosphate pathway and involves transketolase and transaldolase reactions
ii. Each carbon dioxide takes three ATP molecules and two NADPH molecules, thus the formation of a single glucose molecule requires six turns through the cycle with an expenditure of 18 ATP molecules and 12 NADPH molecules; sugars formed in the Calvin cycle can then be used to synthesize other essential molecules
4. Sintesis Gula dan Polisakaridaa. Heterotrophs mensintesis glukosa dari prekusor nonkarbohidrat dalam proses disebut
gluconeogenesis; Jalur gluconeogenic berfungsi pembalikan glikolisis-ini membagi tujuh enzim

dengan jalur glikolitik, pembalikan pengaturan katabolik, dan menggunakan beberapa enzim tertentu atau sistem multienzim untuk langkah katalisis yang tidak dapat secara langsung dibalik
b. Ketika glukosa dan fruktosa disintesis melalui gluconeogenesis, gula lain dibentuk; beberapa dari gula lain disintesis ketika diserang pada nukleosida difosfat
c. Produksi Polisakarida juga membutuhkan penggunaan gula nuklosida difosfat sebagai prekursor5. Assimilasi Senyawa Fosfor, Sulfur, dan Nitrogen Anorganik
a. Assimilasi Senyawa Fosfor i. Fosfat anorganik bergabung pada pembentukan ATP melalui fosforilasi, fosforilasi
oksidatif, dan fosforilasi tingkat substratii. Fosfat Organik didapatkan dari sekeliling dihidrolisis untuk melepaskan fosfat
anorganik oleh enzim yang yang disebut fosfataseb. Assimilasi Sulfur
i. Sulfur Organik dalam bentuk sistein dan metionin didapatkan dari sumber luarii. Reduksi sulfat pengassimilasi digunakan untuk mereduksi sulfat anorganik sebelum ini
bergabung menjadi sisteinc. Assimilasi Nitrogen
i. Penggabungan Ammonia a. Banyak Mikroorganisme menggunakan aminasi tereduksi untuk membentuk
alanin dan glutamat, dimana kemudian digunakan sebagai sumber grup amino; grup amino ditransfer dari alanin atau glutamat ke jaringan karbon lainnya melalui reaksi transaminasi
b. Mikroorganisme lain menggunakan enzim glutamin synthetase dan glutamate synthase untuk mensintesis glutamat, yang kemudian berperan sebagai donor grup amino pada reaksi transaminasi
ii. Reduksi nitrat pengasimilasi-melibatkan reduksi nitrat menjadi nitrit, kemudian menjadi hydroxylamine, dan akhirnya menjadi ammonia, yang kemudian dapat bergabung melalui rute yang digambarkan di atas
iii. Fiksasi Nitrogen – reduksi nitrogen atmosfer menjadi ammonia; ini dikatalisis oleh enzim nitrogenase, yang ditemukan hanya pada sedikit spesies bakteri; fiksasi nitrogen membutuhkan pengeluaran 16 molekul ATP; amonia diproduksi dapat bergabung menjadi molekul organik melalui proses yang digambarkan diatas
6. Sintesis Asam Aminoa. Melibatkan penyerangan grup amino ke jaringan karbonb. Jaringan Karbon diturunkan dari acetyl-CoA dan dari intermediet siklus TCA, glikolisis, dan
jalur pentosa fosfat7. Reaksi Anaplerotik
a. Fungsi Biosintesis siklus TCA juga sangat penting dimana banyak dari intermediet harus disintesis meski ketika siklus TCA tidak berfungsi untuk mengkatabolis piruvat atau menyediakan NADH untuk transport elektron
b. Reaksi Anaplerotic melengkapi kembali intermediet siklus TCA sehingga biosintesis dapat terjadi; dua tipe utama reaksi anapleoritik telah dapat diamati
i. Fiksasi karbondioksida Anaplerotic (misalnya, reaksi karboksilasi piruvat) ii. Siklus Glioksilat cycle-digunakan oleh mikroorganisme yang dapat tumbuh pada asetat
sebagai satu-satunya sumber karbon; ini dimodifikasi siklus TCA8. Sintesis Purin, Pirimidin, dan Nukleotida
a. Molekul ini sangat penting untuk semua sel karena mereka digunakan pada sintesis ATP, beberapa kofaktor, RNA, dan DNA
b. Biosintesis Purin-jalur sangat komplek dimana tujuh molekul berbeda (termasuk asam folat) mengkontribusi bagian akhir kerangka purin; produk pertama purin adalah asam inosinik nukleotida, dari semua nukleotida purin lainnya dapat dibentuk
c. Biosintesis Pirimidin-asm aspartic dan fosfat karbmoksil membentuk produk pirimidin inisial (asam orotic), yang kemudian dapat dikonversikan menjadi nukleotida pirimidin
9. Sintesis Lipida. Sintesis asam lemak dikatalisis oleh asam lemak synthetase menggunakan substrat acetyl-CoA
dan malonyl-CoA, reduktan NADPH, dan protein kecil disebut acyl pembawa protein, yang membawa pertumbuahan rantai asam lemak; asam lemak dipanjangkan dengan penambahan dua karbon pada saat dimana karboksil berakhir
b. Triacylglycerols dibentuk dari reduksi dihydroxyacetone phosphate (a intermediet jalur glikolisis) menjadi glycerol 3-phosphate, yang kemudian menyebabkan esterifikasi dengan dua

asam lemak untuk membentuk asam fosfatidik; ini kemudian dapat digunakan untuk memproduksi triacylglycerol
c. Fosfolipid juga dapat diproduksi dari asam fosfatidik menggunakan pembawa cytidine diphosphate (CDP)
10. Sintesis Peptidoglikana. Prose multilangkah yang melibatkan dua pembawa: uridine diphosphate dan bactoprenol; selama
proses berulang-ulang unit peptidoglycan dibentuk dan diserang menjadi rantai peptidoglikan tumbuh setelah ditransport melalui membran sitoplasmik; crosslinks kemudian dibentuk oleh transpeptidasi
b. Sintesis Peptidoglikan sangat mudah diserang oleh gangguan agen antimikroba, antibiotik yang termasuk seperti penicillin; penghambatan banyak langkah pada proses pelemahan dinding sel dan menyebabkan lisis
11. Pola Pembentukan Dinding Sela. Autolisin menyebabkan digesti terbatas peptidoglikan, dan menyediakan akseptor akhir untuk
penambahan unit baru peptidoglikanb. Dua pola umum aktivitas sintesis dinding sel telah dapat diamati
1. Kebanyakan kokus gram-positive hanya memiliki satu atau sedikit daerah pertumbuhan, biasanya pada sisi pembentukan septum
2. Bakteri berbentuk batang biasanya memiliki sisi pertumbuhan tersebar sepanjang bagian silindrikal sel sama baiknya pada sisi pembentukan septum
http://file.upi.edu/Direktori/FPMIPA/JUR._PEND._BIOLOGI/196805091994031-KUSNADI/BUKU_COMMON_TEXT_MIKROBIOLOGI,_Kusnadi,dkk/REV._BAB_5.pdf