
ANALISIS JUMLAH KROMOSOM BIJI UNGU HASIL … · Pipit, Mbak Diy, Dian, Cicin, Teh Pera, Mbak...
19
ANALISIS JUMLAH KROMOSOM BIJI UNGU HASIL PERSILANGAN JAGUNG VARIETAS PULUT DENGAN BC2 PUTATIF PENGINDUKSI HAPLOID DAN HASIL PERLAKUAN KOLKISIN ISNIANI MUFTIARSARI DEPARTEMEN BIOLOGI FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR BOGOR 2010
-
Upload
trankhuong -
Category
Documents
-
view
222 -
download
0
Transcript of ANALISIS JUMLAH KROMOSOM BIJI UNGU HASIL … · Pipit, Mbak Diy, Dian, Cicin, Teh Pera, Mbak...

ANALISIS JUMLAH KROMOSOM BIJI UNGU HASIL PERSILANGAN
JAGUNG VARIETAS PULUT DENGAN BC2 PUTATIF PENGINDUKSI
HAPLOID DAN HASIL PERLAKUAN KOLKISIN
ISNIANI MUFTIARSARI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2010

ABSTRAK
ISNIANI MUFTIARSARI. Analisis Jumlah Kromosom Biji Ungu Hasil Persilangan Jagung
Varietas Pulut dengan BC2 Putatif Penginduksi Haploid dan Hasil Perlakuan Kolkisin. Dibimbing
oleh ENCE DARMO JAYA SUPENA dan IKA MARISKA.
Metode Bulbosum sangat berpotensi untuk menghasilkan tanaman haploid pada tanaman
jagung. Perkembangan metode Bulbosum pada jagung di Indonesia telah dilakukan sampai tahap
inisiasi yakni persilangan tanaman budidaya dengan galur penginduksi haploid. Penelitian ini
bertujuan melakukan analisis kromosom terhadap biji ungu hasil persilangan varietas Pulut dengan
BC2 putatif penginduksi haploid dan hasil perlakuan kolkisin secara in vitro. Jumlah kromosom
biji jagung ungu hasil persilangan tersebut adalah bukan haploid (x=10) tetapi diploid (2n=2x=20).
Pengaruh kolkisin dengan konsentrasi dan lama perlakuan yang berbeda terhadap level ploidi tidak
dapat diamati pada penelitian ini karena kematian terjadi pada hampir seluruh biji dan kecambah
jagung yang digunakan. Penggandaan jumlah kromosom hanya terjadi pada satu individu hasil
perlakuan akar kecambah dengan konsentrasi kolkisin 0,06% selama 6 jam, yakni menjadi
tetraploid (2n=4x=40), tetapi tanaman tidak dapat bertahan hidup sampai penanaman di rumah
kaca. Satu-satunya tanaman yang bertahan hidup setelah perlakuan kolkisin sampai penanaman di
rumah kaca ialah tetap diploid dengan sifat morfologis dan anatomis yang tidak berbeda dari
tanaman kontrol.
ABSTRACT
ISNIANI MUFTIARSARI. Chromosome Number Analysis of Purple Seed from a Cross Between
Pulut Corn and BC2 Putative Haploid Inducer and from Colchicine Treatment. Supervised by
ENCE DARMO JAYA SUPENA and IKA MARISKA.
Bulbosum method is very potential to produce haploid plant of maize. The development
of Bulbosum method in Indonesia was done until initial step, that is cross between local variety
with haploid inducer line. This research was conducted to analyze the chromosome number of
purple seed from a cross between Pulut and BC2 putative haploid inducer and from in vitro
colchicine treatment. The chromosome number of these purple seed were found not haploid (x=10)
but diploid (2n=2x=20). The effect of colchicine with different concentrations and durations of
treatment to ploidy level was not be able to be observed, because death happened at almost all of
the seeds and seedlings treated. Chromosome doubling was found at treatment of seedling root by
colchicine concentration of 0.06% for 6 hours, that is became tetraploid (2n=4x=40), but the plant
could not survive until planting in greenhouse. The only plant survived after colchicine treatment
until planting in greenhouse was still diploid with morphological and anatomical characters which
were not different from control plants.

ANALISIS JUMLAH KROMOSOM BIJI UNGU HASIL PERSILANGAN
JAGUNG VARIETAS PULUT DENGAN BC2 PUTATIF PENGINDUKSI
HAPLOID DAN HASIL PERLAKUAN KOLKISIN
ISNIANI MUFTIARSARI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2010

Judul : Analisis Jumlah Kromosom Biji Ungu Hasil Persilangan Jagung
Varietas Pulut dengan BC2 Putatif Penginduksi Haploid dan
Hasil Perlakuan Kolkisin
Nama : Isniani Muftiarsari
NIM : G34051891
Disetujui oleh,
Pembimbing I, Pembimbing II,
Dr. Ir. Ence Darmo Jaya Supena, M.Si. Dr. Ir. Ika Mariska, APU
NIP. 19641002 198903 1 002 NIP. 19491027 197903 2 001
Diketahui oleh,
Ketua Departemen Biologi
Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor,
Dr. Ir. Ence Darmo Jaya Supena, M.Si.
NIP. 19641002 198903 1 002
Tanggal Lulus :

PRAKATA
Puji dan syukur penulis panjatkan ke hadirat Allah Swt atas segala rahmat dan karunia-
Nya sehingga karya ilmiah ini berhasil penulis selesaikan. Shalawat dan salam semoga selalu
dilimpahkan kepada Nabi Muhammad SAW, yang karena perjuangan beliau, penulis menempuh
jalan yang benar. Karya ilmiah ini ditulis berdasarkan penelitian yang dilaksanakan bulan Maret
sampai Oktober 2009 di Laboratorium Kultur In Vitro Kelompok Peneliti Biologi Sel dan Jaringan
(Kelti BSJ), Balai Besar Balai Besar Bioteknologi dan Sumberdaya Genetik Pertanian (BB-
Biogen). Penelitian ini termasuk ke dalam rangkaian proyek penelitian Kelti BSJ untuk
mendapatkan tanaman jagung haploid ganda di bawah tanggung jawab Dr. Ir. Ika Mariska, APU.
Penulis menyampaikan terima kasih kepada Dr. Ir. Ence Darmo Jaya Supena, M.Si. dan
Dr. Ir. Ika Mariska, APU atas bimbingan, ilmu, nasehat, waktu, dan perhatian yang diberikan
selama penelitian dan penyusunan karya ilmiah ini, serta Dr. Ir. R. R. Dyah Perwitasari, M.Sc.
selaku penguji yang telah memberikan saran dalam perbaikan karya ilmiah ini, juga Ibu Mia
Kosmiatin, M.Si. dan Teti Mardyatul Khibtiah atas bantuan yang tidak terkira besarnya. Penulis
juga menyampaikan terima kasih kepada Kepala BB-Biogen yang telah memberikan kesempatan
dan fasilitas kepada penulis untuk melaksanakan penelitian ini; serta kepada Pak Joko, Mas Anto,
Pak Iman, Pak Hafidz, Ade, Mawid, Rohmat, teman-teman Biologi UNJ, Reni, Xenia, Shabrina,
Asri, Mbak Retno, dan seluruh staf Laboratorium dan Rumah Kaca Kelti BSJ, Laboratorium Kelti
Mikrobiologi BB-Biogen, dan Laboratorium Biologi Terpadu IPB atas fasilitas dan bantuan
selama penelitian.
Ucapan terima kasih terbesar penulis sampaikan kepada Bunda, Ayah, A Feisal, dan De
Hiban atas cinta, doa, perhatian, dan dukungannya; kepada Bu Dorly, Bu Yudhiwanti, Kak Arif,
Kak Erma, Hapshoh, dan Hida atas saran dan bantuannya; juga kepada Intan, Nani, Uni Iil, Mbak
Pipit, Mbak Diy, Dian, Cicin, Teh Pera, Mbak Nindha, Mbak Maria, Teh Neng, Uni Zikra, Hani,
Citra, Suci, Orie, Dewi, Rinay, Yeni, Izza, Mafri, Upik, Resti, Istirokhah, dan Khairil atas doa dan
dukungannya. Terima kasih juga kepada teman-teman Biologi 41, 42, 43, 44, dan 45 yang selalu
memberikan perhatian dan dukungan.
Semoga karya ilmiah ini bermanfaat, baik bagi penulis, pembaca dari kalangan akademis,
maupun masyarakat secara luas. Amin.
Bogor, Juni 2010
Isniani Muftiarsari

RIWAYAT HIDUP
Penulis dilahirkan di Tasikmalaya pada 20 Juli 1987, dari ayah Jojo Nuryanto, M.Hum.
dan ibu Titin Setiartin, M.Pd., dan merupakan anak kedua dari tiga bersaudara.
Penulis mengenyam pendidikan di SMA Negeri 1 Tasikmalaya (2002-2005). Pada tahun
2005, penulis lulus seleksi Undangan Seleksi Masuk IPB (USMI) dan pada tahun 2006, penulis
diterima di Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut
Pertanian Bogor.
Selama mengikuti perkuliahan, penulis aktif dalam berbagai organisasi, yakni Badan
Kerohanian Islam Mahasiswa (BKIM) IPB periode 2006-2007 dan 2007-2008, Himpunan
Mahasiswa Biologi (Himabio) FMIPA-IPB periode 2006-2007 dan 2007-2008, serta Ikatan
Himpunan Mahasiswa Biologi Indonesia (Ikahimbi) periode 2007-2009. Penulis juga menjadi
asisten praktikum Biologi Dasar pada tahun ajaran 2008-2009 dan 2009-2010 serta Anatomi dan
Morfologi Tumbuhan pada tahun ajaran 2009-2010. Penulis melaksanakan kegiatan Praktik
Lapangan pada tahun 2008 di PT Sinar Inesco Perkebunan Teh Sambawa, Taraju, Tasikmalaya.

DAFTAR ISI
Halaman
DAFTAR TABEL .......................................................................................................... viii
DAFTAR GAMBAR ...................................................................................................... viii
DAFTAR LAMPIRAN .................................................................................................. viii
PENDAHULUAN .......................................................................................................... 1
BAHAN DAN METODE ............................................................................................... 1
Waktu dan Tempat .................................................................................................... 1
Bahan Tanaman ........................................................................................................ 2
Metode ...................................................................................................................... 2
Penggandaan Jumlah Kromosom in vitro dengan Kolkisin ...................................... 2
Analisis Sitologi ........................................................................................................ 2
Analisis Morfologi .................................................................................................... 2
Analisis Anatomi ...................................................................................................... 2
HASIL ............................................................................................................................ 3
Jumlah Kromosom dan Daya Hidup ......................................................................... 3
Analisis Morfologi .................................................................................................... 4
Analisis Anatomi ...................................................................................................... 4
PEMBAHASAN ............................................................................................................. 5
SIMPULAN DAN SARAN ............................................................................................ 6
Simpulan ................................................................................................................... 6
Saran ......................................................................................................................... 6
DAFTAR PUSTAKA ..................................................................................................... 6
LAMPIRAN ................................................................................................................... 8

DAFTAR TABEL
Halaman
1 Hubungan perlakuan kolkisin terhadap daya hidup dan level ploidi biji dan
kecambah jagung ....................................................................................................... 3
2 Hubungan perlakuan kolkisin terhadap morfologi tanaman jagung pada umur 7
minggu setelah tanam (MST) ..................................................................................... 4
3 Jumlah dan rasio jumlah biji hasil selfing tanaman jagung ........................................ 5
4 Ukuran dan kerapatan stomata jagung kontrol dan perlakuan biji K0,03%; 24 jam .. 5
DAFTAR GAMBAR
Halaman
1 Kematian biji dan kecambah pasca perlakuan kolkisin ............................................. 4
2 Jumlah kromosom tanaman jagung biji ungu hasil perlakuan akar kecambah
K0,06%; 6 jam sebelum dan setelah perendaman dengan kolkisin ........................... 4
3 Jagung hasil penyerbukan sendiri tanaman diploid ................................................... 5
4 Kerapatan stomata tanaman jagung biji ungu diploid ............................................... 5
DAFTAR LAMPIRAN
Halaman
1 Komposisi media MS (Murashige & Skoog 1962) .................................................. 9
2 Pembuatan larutan kolkisin ....................................................................................... 10
3 Pengujian percobaan monohibrid mendel untuk warna biji jagung hasil perlakuan
K0,03%; 24 jam pada biji ......................................................................................... 11
4 Pengujian percobaan monohibrid mendel untuk warna biji jagung (dati total lima
tanaman kontrol) ....................................................................................................... 11

1
PENDAHULUAN
Kebutuhan akan jagung (Zea mays) di
Indonesia sangat tinggi karena dimanfaatkan
sebagai bahan pangan, pakan, dan berbagai
keperluan industri. Pada tahun 2008, produksi
jagung dalam negeri sebesar 16,32 juta ton
dari 4 juta ha lahan, dengan produktivitas
tanaman sekitar 4 ton/ha (BPS 2009).
Produktivitas ini jauh di bawah potensi
produktivitas jagung hibrida, yakni sebesar
10-11 ton/ha (Iriany & Takdir 2007).
Departemen Pertanian terus melakukan upaya
peningkatan produktivitas melalui
penggunaan benih unggul bermutu terutama
benih hibrida serta pemanfaatan pupuk
berimbang dan pupuk organik (Ditjentan
2008). Penggunaan benih hibrida di Indonesia
baru sekitar 40% dari total lahan jagung
nasional (Depkominfo 2008).
Benih hibrida merupakan hasil persilangan
antara dua galur murni. Kelebihan benih
jagung hibrida dibandingkan benih bersari
bebas (open pollinated) terutama memiliki
potensi hasil yang lebih tinggi, responsif
terhadap nitrogen, toleran terhadap kepadatan
tanaman yang tinggi, dan resisten terhadap
penyakit dan hama (Dhillon 1998) dengan
kualitas bulir yang lebih baik (Delorit et al.
1974).
Tanaman galur murni sebagai tetua hibrida
dapat diperoleh dari tanaman haploid yang
telah digandakan kromosomnya menjadi
haploid ganda kemudian dilakukan
penyerbukan sendiri (selfing) (Eder & Chalyk
2002). Terdapat empat metode penting dalam
pembentukan tanaman haploid secara in vitro,
yaitu kultur antera, kultur mikrospora, kultur
ovari, dan penyelamatan embrio dari hasil
persilangan jauh (metode Bulbosum). Metode
Bulbosum dapat dilakukan melalui
persilangan antara dua jenis tanaman yang
hubungan kekerabatannya jauh (berbeda
spesies) atau antara dua tanaman dari spesies
yang sama tetapi tetua jantannya memiliki
kemampuan untuk menginduksi terbentuknya
tanaman haploid.
Metode Bulbosum pertama kali
dikembangkan untuk memperoleh tanaman
haploid barley dari hasil persilangan Hordeum
vulgare (betina) dan H. bulbosum (jantan)
dengan adanya proses eliminasi kromosom H.
bulbosum (Chahal & Gosal 2006). Pada
jagung, metode ini dikembangkan sejak tahun
2000 di Australia dan Jerman dari persilangan
dua tanaman jagung dengan tetua jantan yang
memiliki kemampuan menginduksi haploid.
Untuk mengidentifikasi haploid, digunakan
sistem gen penanda R1-nj yang ekspresinya
menyebabkan warna ungu pada biji hasil
persilangan tersebut (Nanda & Chase 1966).
Persilangan varietas Bisma yang berbiji
kuning sebagai tetua betina dengan galur
penginduksi haploid (nomor aksesi 3490)
yang berbiji putih sebagai tetua jantan
menghasilkan biji berwarna kuning dan putih.
Biji kuning hasil persilangan ini disilangkan
secara backcross dengan tetua jantan sehingga
menghasilkan biji BC1 yang berwarna ungu
sebanyak 1,5% dan sisanya berwarna kuning
dan putih. Berdasarkan analisis sitologi,
semua biji ungu terbukti haploid, sedangkan
biji yang berwarna kuning dan putih adalah
diploid (Mariska et al. 2007). Selanjutnya,
dilakukan backcross lagi tanaman yang
berasal dari biji kuning BC1 terhadap tetua
jantan (3490) sehingga menghasilkan BC2
yang memiliki biji dengan komposisi warna
yang sama dengan BC1, dengan persentase
biji yang berwarna ungu sebesar 12,5%. Biji
BC2 yang berwarna kuning kemudian
digunakan sebagai tetua jantan dalam
persilangan dengan varietas Pulut yang
berwarna putih. Persilangan ini menghasilkan
biji yang berwarna ungu dan berwarna putih.
Biji yang berwarna ungu diduga merupakan
biji haploid, dengan asumsi BC2 telah
memiliki sifat penginduksi dari tetuanya. Biji
putatif haploid ini berpotensi untuk digunakan
dalam menghasilkan galur haploid ganda
melalui perlakuan kolkisin (diploidisasi).
Perlakuan kolkisin diketahui dapat
menggandakan jumlah kromosom. Level
ploidi yang meningkat akibat perlakuan
kolkisin menyebabkan morfologi tanaman
menjadi lebih tinggi (Rober et al. 2005) dan
fertil (Eder & Chalyk 2002). Selain itu,
tanaman memiliki stomata yang lebih besar
dengan kerapatan lebih rendah dibandingkan
tanaman dengan level ploidi yang lebih
rendah (Wongpiyasatid et al. 2005).
Penelitian ini bertujuan melakukan analisis
kromosom terhadap biji ungu hasil
persilangan jagung varietas Pulut dengan BC2
putatif penginduksi haploid dan hasil
perlakuan kolkisin secara in vitro.
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilaksanakan pada bulan Maret
sampai Oktober 2009 di Laboratorium Kultur
In Vitro Kelompok Peneliti Biologi Sel dan
Jaringan, Balai Besar Bioteknologi dan
Sumberdaya Genetik Pertanian, Bogor.

2
Bahan Tanaman
Biji jagung ungu hasil persilangan varietas
Pulut berbiji putih x biji kuning BC2 (((Bisma
x 3490) x 3490) x 3490) yang diduga dapat
menginduksi haploid.
Metode
Perlakuan penggandaan kromosom
diberikan terhadap biji dan kecambah jagung
dengan konsentrasi kolkisin 0; 0,03; 0,06; dan
0,1% (Gayen et al. 1994), dan lama perlakuan
6, 16, dan 24 jam. Pada perlakuan biji,
digunakan masing-masing 10 biji untuk
kombinasi perlakuan waktu dan konsentrasi
kolkisin. Sedangkan pada perlakuan akar
kecambah hanya digunakan masing-masing 2
kecambah karena keterbatasan bahan.
Penggandaan Jumlah Kromosom in vitro
dengan Kolkisin
Biji jagung ungu disterilisasi dengan
alkohol 96 dan 70% selama masing-masing 5
menit dengan dua kali pengulangan, kemudian
disterilisasi dalam larutan kloroks (5% Na-
hipoklorit) berturut-turut dalam konsentrasi 30
dan 20% masing-masing 15 menit, dan
dibilas dengan akuades steril. Biji direndam di
dalam akuades steril selama satu malam. Agar
perlakuan efektif, biji dipotong menjadi
setengah dengan membuang perikarp dan
sebagian endosperma. Biji ditanam pada
media MS dan diberi perlakuan kolkisin
dengan kombinasi konsentrasi dan lama
perlakuan, sedangkan kontrol ditanam tanpa
diberi kolkisin. Biji yang telah diberi
perlakuan dibilas dengan akuades steril dan
ditanam pada media MS (Murashige & Skoog
1962) (Lampiran 1) yang diperkaya GA3 10
mg/l, kemudian diinkubasi tanpa cahaya di
ruang kultur dengan temperatur ruang 25oC
sampai berkecambah.
Pada perlakuan akar kecambah, biji yang
telah disterilisasi ditanam terlebih dahulu
sampai berkecambah kemudian bagian
akarnya direndam dalam kolkisin dengan
kombinasi perlakuan yang sama. Setelah itu,
kecambah hasil perlakuan ditanam pada media
MS sampai tumbuh akar yang baru.
Baik biji maupun kecambah hasil
perlakuan kolkisin kemudian diamati daya
hidupnya sampai penanaman di rumah kaca.
Analisis Sitologi
Penghitungan jumlah kromosom dilakukan
dengan menggunakan metode Darnaedi
(1991). Sebanyak dua akar dari tiap kecambah
dipotong masing-masing sepanjang ±1 cm dan
dicuci dengan akuades. Ujung akar direndam
dalam hydroxyquinolin 0,3 g/l dan disimpan
pada suhu 20oC selama 3-5 jam dalam kondisi
tanpa cahaya, sedangkan kecambah
diaklimatisasi ke media tanah dalam polibag.
Ujung akar difiksasi dengan larutan asam
asetat 45% selama 10 menit, kemudian
dimaserasi dalam larutan HCl 1N : asam
asetat 45% = 3:1 selama 1-3 menit pada suhu
60oC dan dicuci dengan akuades. Pewarnaan
dilakukan dengan perendaman di dalam aceto
orcein 2% selama 15 menit. Bagian ujungnya
dipotong sepanjang 1-2 mm dan diletakkan
pada gelas objek, diberi 1-2 tetes aceto orcein
lagi dan ditutup dengan gelas penutup.
Preparat di-squash menggunakan ujung pensil
berkaret, kemudian dilewatkan di atas api
sebanyak 2-3 kali. Preparat diamati
menggunakan mikroskop cahaya dengan
perbesaran 400x dan perbesaran 1000x untuk
menghitung jumlah kromosom. Pengambilan
gambar dilakukan menggunakan mikroskop
Olympus yang dilengkapi dengan kamera
DP20 untuk mempermudah penghitungan
jumlah kromosom.
Analisis Morfologi
Sebelum aklimatisasi, akar tanaman dicuci
hingga tidak ada media agar yang menempel
dan direndam di dalam fungisida Dithane 1
mg/l selama 5 menit. Tanaman ditanam di
dalam polibag berisi tanah : pupuk kandang =
1:1 yang telah disterilisasi dan ditutup dengan
plastik. Untuk mengurangi resiko kematian,
dilakukan praaklimatisasi, yakni tanaman
disimpan di ruang kultur dengan temperatur
ruang 25oC hingga tumbuh tegar (sekitar satu
minggu). Tanaman dipindahkan ke rumah
kaca dengan sistem buka-tutup sungkup setiap
pagi dengan lama pembukaan meningkat
setiap harinya, agar dapat beradaptasi dengan
kondisi luar. Setelah tumbuh tegar ketika
sungkup dibuka, tanaman dipindahkan ke
tanah. Pengamatan morfologi dilakukan pada
semua tanaman yang berhasil hidup sampai
penanaman di rumah kaca. Tanaman diamati
pada umur 7 minggu setelah tanam (MST),
meliputi panjang tajuk, jumlah daun, dan rasio
jumlah biji hasil penyerbukan sendiri.
Analisis Anatomi
Analisis stomata meliputi kerapatan,
panjang, dan lebar stomata pada umur 4 MST.
Pengamatan stomata adalah daun ketiga dari
ujung tajuk tanaman, dan sampel stomata
diambil dari bagian tengah daun tersebut.
Kuteks dioleskan pada bagian abaksial,
kemudian diletakkan pada gelas objek dan
ditutup dengan gelas penutup. Analisis
3
dilakukan dengan mikroskop cahaya pada
lima titik bidang pandang yang berbeda.
Kerapatan stomata dihitung pada perbesaran
400x, sedangkan panjang dan lebar stomata
diukur pada perbesaran 1000x.
HASIL
Jumlah Kromosom dan Daya Hidup
Semua biji kontrol dapat berkecambah dan
semua tanaman dapat hidup sampai
aklimatisasi, tetapi hanya 5 dari 10 tanaman
yang hidup sampai penanaman di rumah kaca
(Tabel 1). Berdasarkan penghitungan jumlah
kromosom, biji ini adalah diploid (2n=2x=20).
Pada perlakuan biji dengan kolkisin, hanya 17
dari 90 biji dapat berkecambah dan hanya satu
tanaman yang hidup sampai penanaman di
rumah kaca, yakni perlakuan K0,03%; 24 jam
(Tabel 1). Tanaman hasil perlakuan biji
K0,03%; 24 jam yang bertahan hidup ternyata
mempunyai jumlah kromosom yang sama
dengan kontrol, yaitu diploid.
Pada perlakuan akar kecambah, 14 dari 20
kecambah hidup setelah perlakuan, tetapi
hanya 2 kecambah sampai aklimatisasi dan
tidak diperoleh tanaman yang hidup sampai
penanaman di rumah kaca (Tabel 1 dan
Gambar 1). Dari hanya empat kecambah yang
dapat dianalisis, diperoleh satu tanaman hasil
perlakuan K0,06%; 6 jam menunjukkan
terjadinya penggandaan kromosom menjadi
tetraploid (Gambar 2), tetapi kecambah tidak
dapat bertahan hidup sampai penanaman di
rumah kaca sehingga tidak dapat dianalisis
morfologi maupun anatomi.
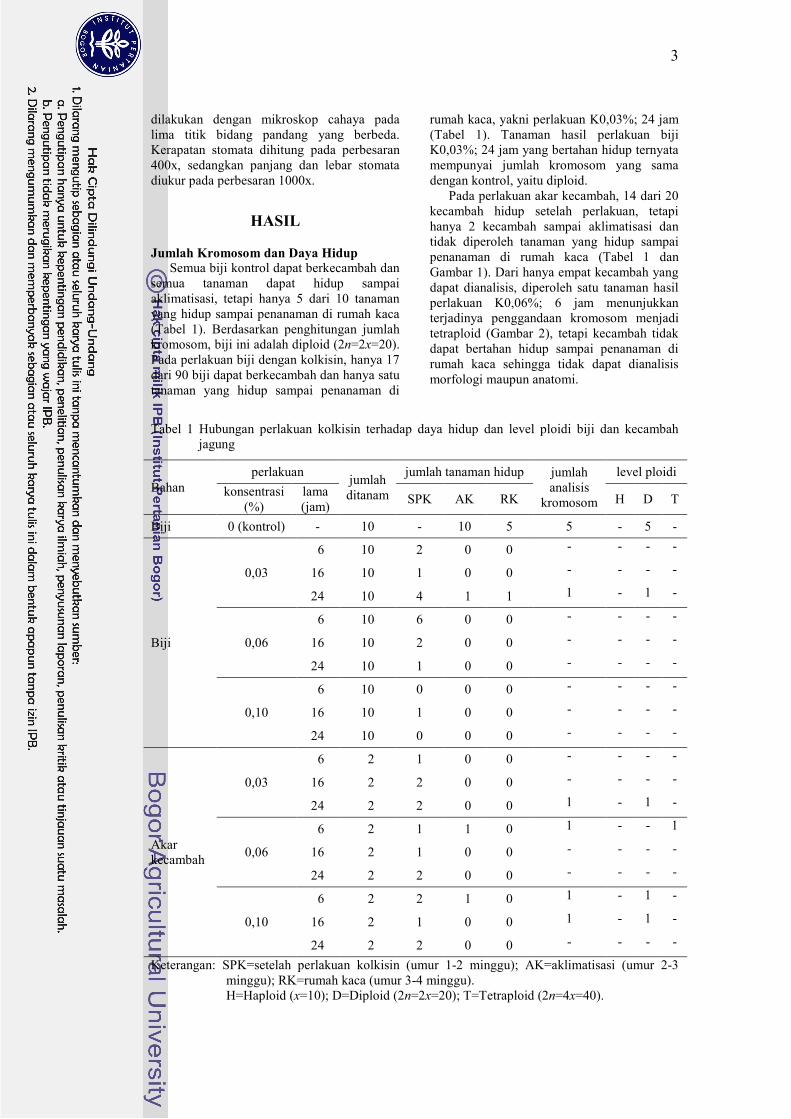
Tabel 1 Hubungan perlakuan kolkisin terhadap daya hidup dan level ploidi biji dan kecambah
jagung
Bahan
perlakuan jumlah
ditanam
jumlah tanaman hidup jumlah
analisis
kromosom
level ploidi
konsentrasi
(%)
lama
(jam) SPK AK RK H D T
Biji 0 (kontrol) - 10 - 10 5 5 - 5 -
Biji
0,03
06 10 2 00 0 - - - -
16 10 1 00 0 - - - -
24 10 4 01 1 1 - 1 -
0,06
06 10 6 00 0 - - - -
16 10 2 00 0 - - - -
24 10 1 00 0 - - - -
0,10
06 10 0 00 0 - - - -
16 10 1 00 0 - - - -
24 10 0 00 0 - - - -
Akar
kecambah
0,03
06 02 1 00 0 - - - -
16 02 2 00 0 - - - -
24 02 2 00 0 1 - 1 -
0,06
06 02 1 01 0 1 - - 1
16 02 1 00 0 - - - -
24 02 2 00 0 - - - -
0,10
06 02 2 01 0 1 - 1 -
16 02 1 00 0 1 - 1 -
24 02 2 00 0 - - - -
Keterangan: SPK=setelah perlakuan kolkisin (umur 1-2 minggu); AK=aklimatisasi (umur 2-3
minggu); RK=rumah kaca (umur 3-4 minggu).
H=Haploid (x=10); D=Diploid (2n=2x=20); T=Tetraploid (2n=4x=40).

Gambar 1 Kematian biji dan kecambah
perlakuan biji saat kultur; (B) kematian eksplan asal perlakuan akar kecambah saat
kultur; (C) kematian tanaman asal perlakuan biji saat aklimatisasi; (D) kematian
tanaman asal perlakuan akar kecambah saa
Gambar 2 Jumlah kromosom tanaman jagung biji ungu hasil perlakuan akar kecambah K0
jam: (A) sebelum perendaman (2
(2n=4x=40). Bar = 20µm.
Analisis Morfologi
Satu-satunya tanaman yang hidup hasil
perlakuan biji K0,03%; 24 jam dan
tanaman kontrol menunjukkan panjang tajuk
dan jumlah daun yang sama pada umur 7
MST (Tabel 2). Pada 73 hari setelah tanam
(HST) aklimatisasi, baik pada tanaman
kontrol maupun tanaman perlakuan biji
K0,03%; 24 jam, dilakukan penyerbukan
sendiri. Jagung hasil penyerbukan sendiri ini
mempunyai jumlah total biji yang berbeda
pada tanaman kontrol maupun hasil
Tabel 2 Hubungan perlakuan kolkisin
setelah tanam (MST)
Bahan/kode
Biji K0,03%; 24
Biji kontrol
*) hanya dari satu tanaman (kontrol dari
A
A
Gambar 1 Kematian biji dan kecambah pasca perlakuan kolkisin: (A) kematian eksplan asal
perlakuan biji saat kultur; (B) kematian eksplan asal perlakuan akar kecambah saat
kultur; (C) kematian tanaman asal perlakuan biji saat aklimatisasi; (D) kematian
tanaman asal perlakuan akar kecambah saat aklimatisasi.
Gambar 2 Jumlah kromosom tanaman jagung biji ungu hasil perlakuan akar kecambah K0
jam: (A) sebelum perendaman (2n=2x=20); (B) setelah perendaman dengan kolkisin
Bar = 20µm.
tanaman yang hidup hasil
03%; 24 jam dan lima
menunjukkan panjang tajuk
dan jumlah daun yang sama pada umur 7
Pada 73 hari setelah tanam
(HST) aklimatisasi, baik pada tanaman
kontrol maupun tanaman perlakuan biji
03%; 24 jam, dilakukan penyerbukan
hasil penyerbukan sendiri ini
mempunyai jumlah total biji yang berbeda
pada tanaman kontrol maupun hasil
perlakuan, tetapi memiliki rasio jumlah biji
ungu dan putih yang sama untuk setiap
tongkol, yaitu 1:1 (Tabel 3
Analisis Anatomi
Berdasarkan analisis stomata daun,
diperoleh kerapatan stomata tanaman
perlakuan biji K0,03%; 24 jam tidak berbeda
dengan kisaran tanaman kontrol (Tabel
Gambar 4). Begitu pula dengan panjang dan
lebar stomata yang tidak berbeda
dibandingkan dengan tanaman kontrol.
Hubungan perlakuan kolkisin terhadap morfologi tanaman jagung pada umur
setelah tanam (MST)
Bahan/kode perlakuan panjang tajuk (cm) jumlah daun
03%; 24 jam* 220 9
210-240 8-
hanya dari satu tanaman (kontrol dari kisaran lima tanaman)
B
B C D
4
pasca perlakuan kolkisin: (A) kematian eksplan asal
perlakuan biji saat kultur; (B) kematian eksplan asal perlakuan akar kecambah saat
kultur; (C) kematian tanaman asal perlakuan biji saat aklimatisasi; (D) kematian
Gambar 2 Jumlah kromosom tanaman jagung biji ungu hasil perlakuan akar kecambah K0,06%; 6
=20); (B) setelah perendaman dengan kolkisin
perlakuan, tetapi memiliki rasio jumlah biji
h yang sama untuk setiap
dan Gambar 3).
Berdasarkan analisis stomata daun,
diperoleh kerapatan stomata tanaman
03%; 24 jam tidak berbeda
tanaman kontrol (Tabel 4 dan
Begitu pula dengan panjang dan
lebar stomata yang tidak berbeda
dibandingkan dengan tanaman kontrol.
pada umur 7 minggu
jumlah daun
9
-9
D

Tabel 3 Jumlah dan rasio jumlah biji hasil
Bahan/kode perlakuan
Biji K0,03%; 24
Biji kontrol
*) hanya dari satu tanaman (kontrol dari
**) berdasarkan uji χ
Gambar 3 Jagung hasil penyerbukan sendiri
Tabel 4 Ukuran dan kerapatan stomata jagung kontrol dan perlakuan biji K0
Asal tanaman
Biji K0,03%; 24 jam*
Biji kontrol
*) hanya dari satu tanaman (
Gambar 4 Kerapatan stomata tanaman jagung biji ungu
K0,03%; 24 jam
PEMBAHASAN
Berdasarkan analisis sitologi pada tanaman
kontrol, ternyata biji ungu hasil persilangan
varietas Pulut dengan tanaman dari biji kuning
BC2 yang diduga haploid
(2n=2x=20). Hasil ini membuktikan bahwa
tanaman dari biji kuning BC2 tidak bersifat
penginduksi haploid atau induksi haploid
tidak terjadi pada jagung jenis Pulut. Zhang
al. (2008) melaporkan persentase terjadinya
eliminasi kromosom sebagai mekanisme
A
A
Jumlah dan rasio jumlah biji hasil selfing tanaman jagung
Bahan/kode perlakuan jumlah biji rasio**
ungu putih ungu
03%; 24 jam* 137 114 1
352 307 1
*) hanya dari satu tanaman (kontrol dari total lima tanaman)
χ2 (Lampiran 3 dan 4)
Jagung hasil penyerbukan sendiri tanaman diploid: (A) biji K0,03%; 24 jam; (B)
Ukuran dan kerapatan stomata jagung kontrol dan perlakuan biji K0,03%; 24 jam
ukuran stomata (µm) kerapatan stomata
(jumlah stomata/mmpanjang lebar
jam* 58,2 26,3 97
48,7-55,3 23,3-32,6 92-117
*) hanya dari satu tanaman (kontrol dari kisaran lima tanaman)
Kerapatan stomata tanaman jagung biji ungu diploid: (A) tanaman berasal dari
03%; 24 jam; (B) tanaman kontrol. Bar = 50µm.
PEMBAHASAN
Berdasarkan analisis sitologi pada tanaman
biji ungu hasil persilangan
varietas Pulut dengan tanaman dari biji kuning
yang diduga haploid adalah diploid
=20). Hasil ini membuktikan bahwa
tanaman dari biji kuning BC2 tidak bersifat
penginduksi haploid atau induksi haploid
tidak terjadi pada jagung jenis Pulut. Zhang et
. (2008) melaporkan persentase terjadinya
eliminasi kromosom sebagai mekanisme
induksi haploid berbeda pada setiap genotipe,
bahkan tidak terjadi pada genotipe tertentu.
Kematian biji dan kecambah yang tinggi
pada penelitian ini diduga karena vigor biji
yang digunakan rendah. Biji jagung kontrol
dapat berkecambah 100% atau viabilitas
100% dan tumbuh dengan baik sampai
aklimatisasi, tetapi ketika penanaman di
rumah kaca yang hidup hanya tersisa 50%
(Tabel 1). Vigor biji yang rendah dapat
terekspresi lebih awal pada kondisi cekaman
(Ditjen BPTP 2004). Perlakuan kolkisin pada
penelitian ini merupakan cekaman bagi
B
B
stomata
5
rasio**
putih
1
1
03%; 24 jam; (B) kontrol
03%; 24 jam
kerapatan stomata
(jumlah stomata/mm2)
97,0
117
: (A) tanaman berasal dari biji
haploid berbeda pada setiap genotipe,
bahkan tidak terjadi pada genotipe tertentu.
Kematian biji dan kecambah yang tinggi
pada penelitian ini diduga karena vigor biji
yang digunakan rendah. Biji jagung kontrol
dapat berkecambah 100% atau viabilitas
n tumbuh dengan baik sampai
aklimatisasi, tetapi ketika penanaman di
rumah kaca yang hidup hanya tersisa 50%
(Tabel 1). Vigor biji yang rendah dapat
terekspresi lebih awal pada kondisi cekaman
(Ditjen BPTP 2004). Perlakuan kolkisin pada
pakan cekaman bagi

6
tanaman karena kolkisin merupakan senyawa
toksik. Hal ini mengakibatkan biji dan
kecambah dengan vigor rendah pada
penelitian ini menjadi mati ketika atau segera
setelah mendapatkan perlakuan. Viabilitas dan
vigor biji berbeda pada setiap genotipe atau
kultivar tanaman (Justice & Bass 2002).
Analisis sitologi pada satu-satunya
tanaman yang bertahan hidup, yaitu hasil
perlakuan biji K0,03%; 24 jam, menunjukkan
tidak terjadi penggandaan kromosom (Tabel
1). Meskipun ada perlakuan yang berhasil
menggandakan kromosom dari diploid
(2n=2x=20) menjadi tetraploid (2n=4x=40),
yaitu pada perlakuan akar kecambah K0,06%;
6 jam, tanaman tidak dapat bertahan hidup
sampai penanaman di rumah kaca (Tabel 1).
Berdasarkan keberhasilan penggandaan
kromosom, perlakuan akar kecambah diduga
lebih efektif dibandingkan dengan perlakuan
biji. Menurut Chang & Coe (2009) dan Geiger
(2009), perlakuan kolkisin dengan cara
perendaman kecambah sangat efektif pada
jagung, karena penggandaan kromosom
terjadi pada seluruh bagian tanaman. Namun
pada penelitian ini, jumlah tanaman yang
hidup setelah perlakuan kolkisin tidak
representatif untuk menduga dengan akurat
efektifitas dan efisiensi pengaruh perlakuan
pada biji dan akar kecambah.
Analisis morfologi dan anatomi pada
tanaman hasil perlakuan biji K0,03%; 24 jam
dan tanaman kontrol yang sama-sama diploid
diperoleh tidak menunjukkan perbedaan
(Tabel 2 dan 4 dan Gambar 4). Kesamaan
morfologi ditunjukkan dari panjang tajuk dan
jumlah daun pada umur 7 MST (Tabel 2).
Sedangkan kesamaan anatomi ditunjukkan
dari ukuran dan kerapatan stomata yang tidak
berbeda (Tabel 3). Persamaan lain antara
tanaman kontrol dan tanaman yang berasal
dari biji perlakuan K0,03%; 24 jam tersebut
adalah dari nisbah biji ungu : biji putih hasil
penyerbukan sendiri, yaitu 1:1 (Lampiran 3
dan 4). Hal ini mengindikasikan selain tidak
terjadi perubahan ploidi, juga tidak terjadi
mutasi pada tingkat gen yang ditandai dengan
kesamaan dalam segregasi alel.
SIMPULAN DAN SARAN
Simpulan
Jumlah kromosom biji jagung ungu hasil
persilangan varietas Pulut dengan galur putatif
penginduksi haploid BC2 adalah diploid
(2n=2x=20). Pengaruh kolkisin dengan
konsentrasi dan lama perlakuan yang berbeda
terhadap level ploidi tidak dapat diamati pada
penelitian ini karena kematian terjadi pada
hampir seluruh biji dan kecambah jagung
yang digunakan. Tingkat kematian yang tinggi
pada biji dan kecambah disebabkan oleh vigor
biji yang rendah sehingga tidak tahan diberi
perlakuan kolkisin. Penggandaan jumlah
kromosom terjadi pada perlakuan akar
kecambah K0,06%; 6 jam, yakni dari diploid
menjadi tetraploid (2n=4x=40), tetapi tanaman
tidak dapat bertahan hidup hingga penanaman
di rumah kaca. Satu-satunya tanaman yang
bertahan hidup setelah perlakuan kolkisin
sampai penanaman di rumah kaca adalah tetap
diploid dengan sifat morfologis dan anatomis
yang tidak berbeda dari tanaman kontrol.
Saran
Sebelum diploidisasi in vitro dengan
perlakuan kolkisin, perlu dipastikan dahulu
biji jagung yang digunakan merupakan
haploid. Untuk mengetahui apakah BC2
memiliki sifat penginduksi haploid, perlu
dilakukan analisis sitologi terhadap biji ungu
hasil persilangan biji putih BC2 dengan
varietas Pulut. Selain itu, dapat dilakukan
persilangan BC2 terhadap genotipe yang
sudah diketahui dapat diinduksi dengan galur
penginduksi (3490), seperti Bisma. Viabilitas
dan vigor biji yang digunakan perlu dipastikan
sebelum percobaan karena bergantung pada
genotipe atau varietas jagungnya.
DAFTAR PUSTAKA
[BPS]. Biro Pusat Statistik. 2009. Harvested
Area, Yield Rate, and Production of
Maize by Province, 2008. [terhubung
berkala] http://www.bps.go.id [3 Des
2009].
Chahal GS, Gosal SS. 2006. Principles and
Procedures of Plant Breeding,
Biotechnological and Conventional
Approaches. Harrow: Alpha Sci Int’l.
Chang MT, Coe EH. 2009. Doubled Haploids.
Di dalam: Kriz AL, Larkins BA, editor.
Molecular Genetic Approaches to
Maize Improvement Vol 63. Berlin:
Springer-Verlag. hlm 127-142.
Darnaedi D. 1991. Informasi Kromosom.
Makalah dalam Pelatihan Sitogenetika
Tumbuhan. Bogor: Puslitbang Biologi-
LIPI.
Delorit RJ, Greub LJ, Ahlgren HL. 1974.
Crop Production. New Jersey:
Prentice-Hall, Inc.

7
[Depkominfo]. Departemen Komunikasi dan
Informasi. 2008. Deptan Hentikan
Impor Jagung Tahun 2009. [terhubung
berkala] http://www.depkominfo.go.id
[24 Jan 2009].
[Ditjen BPTP]. Direktorat Jenderal Bina
Produksi Tanaman Pangan. 2004.
Pengujian Mutu Benih Tanaman
Pangan dan Hortikultura. Depok:
Balai Pengembangan Mutu Benih
Tanaman Pangan dan Hortikultura.
[Ditjentan]. Direktorat Jenderal Tanaman
Pangan. 2008. Pedoman Pelaksanaan
Kegiatan Pembangunan Tanaman
Pangan TA 2009 [terhubung berkala].
http://ditjentan.deptan.go.id [31 Jan
2009].
Dhillon BS. 1998. Maize. Di dalam: Banga
SS, Banga SK, editors. Hybrid Cultivar
Development. New Delhi: Narosa Pub
House. hlm 282-315.
Eder J, Chalyk S. 2002. In vitro haploid
induction in maize. Theor Appl Genet.
104:703–708.
Gayen P, Madan JK, Kumar R, Sarkar KR.
1994. Chromosome doubling in
haploids through colchicines. Maize
Genet Coop Newslett. 68:65.
Geiger HH. 2009. Doubled Haploids. Di
dalam: Bennetzen JL, Hake S, editor.
Maize Handbook Vol. II: Genetics and
Genomics. New York: Springer
Secience+Business Media. hlm 641-
657.
Iriany RN, Takdir A. 2007. Jagung hibrida
unggul baru. Warta Litbangtan 29:1-3.
Justice OL, Bass LN. 2002. Prinsip dan
Praktek Penyimpanan Benih. Roesli R,
penerjemah; Jakarta: Raja Grafindo
Persada. Terjemahan dari: Principles
and Practices of Seed Storage.
Mariska I, Kosmiatin M, Hutami S,
Purnamaningsih R, Budiarti SG,
Supriati Y, Adil WH. 2007. Laporan
hasil penelitian: Pembentukan tanaman
dihaploid jagung melalui kultur anther.
Bogor: BB-Biogen.
Murashige T, Skoog F. 1962. A revised
medium for rapid growth and bio-assay
with tobacco tissue cultures. Physiol
Plant. 15:473-497.
Nanda DK, Chase SS. 1966. An embryo
marker for detecting monoploids of
maize (Zea mays L.) [abstrak]. Crop
Sci. 6:213-215.
Rober FK, Gordillo GA, Geiger HH. 2005. In
vivo haploid induction in maize–
performance of new inducers and
significance of doubled haploid lines in
hybrid breeding. Maydica. 50:275-283.
Wongpiyasatid A, Hormchan P, Chusreeaeom
K, Ratanadilok N. 2005. Stomatal size,
stomatal frequency and pollen grain
diameter as indirect method for
identification of ploidy levels in cotton.
Kasetsart J. 39:552-559.
Zhang Z, Qiu F, Liu Y, Ma K, Li Z, Xu S.
2008. Chromosome elimination and in
vivo haploid production induced by
Stock 6-derived inducer line in maize
(Zea mays L.). Plant Cell Rep.
27:1851-1860.

8
LAMPIRAN
9
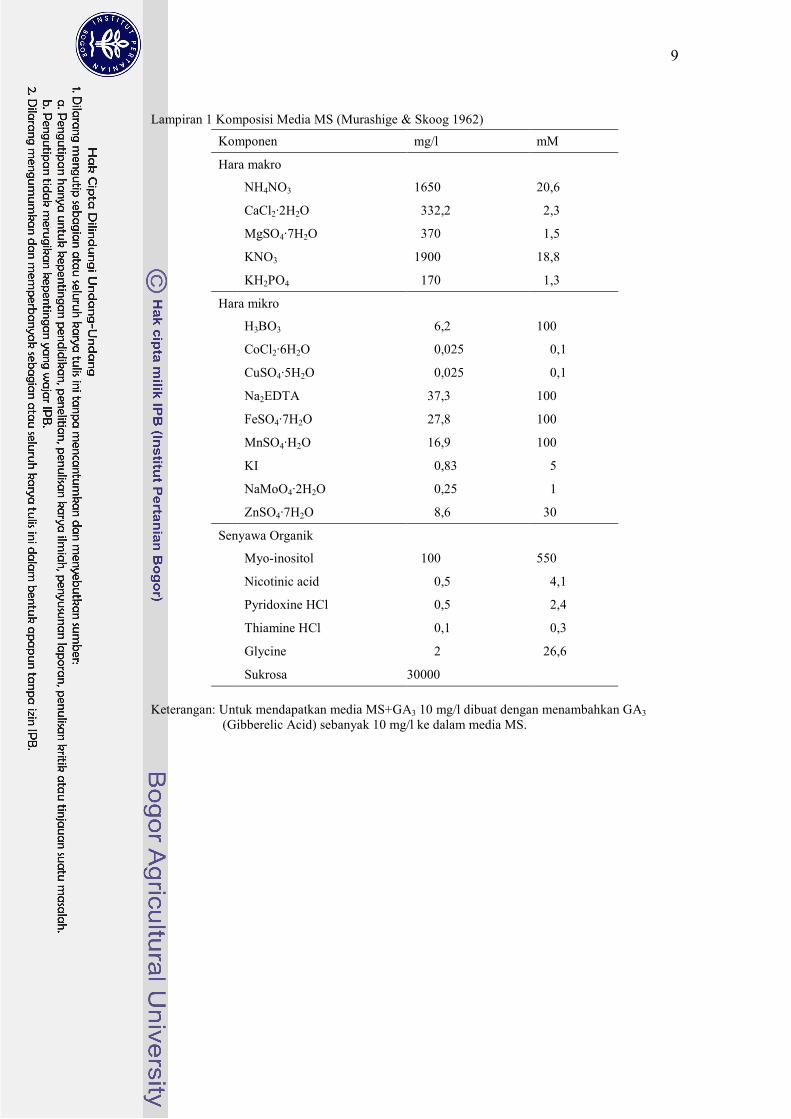
Lampiran 1 Komposisi Media MS (Murashige & Skoog 1962)
Keterangan: Untuk mendapatkan media MS+GA3 10 mg/l dibuat dengan menambahkan GA3
(Gibberelic Acid) sebanyak 10 mg/l ke dalam media MS.
Komponen mg/l mM
Hara makro
NH4NO3 1650 20,6
CaCl2·2H2O 0332,2 02,3
MgSO4·7H2O 0370 01,5
KNO3 1900 18,8
KH2PO4 0170 01,3
Hara mikro
H3BO3 0006,2 100
CoCl2·6H2O 0000,025 000,1
CuSO4·5H2O 0000,025 000,1
Na2EDTA 0037,3 100
FeSO4·7H2O 0027,8 100
MnSO4·H2O 0016,9 100
KI 0000,83 005
NaMoO4·2H2O 0000,25 001
ZnSO4·7H2O 0008,6 030
Senyawa Organik
Myo-inositol 0100 550
Nicotinic acid 0000,5 004,1
Pyridoxine HCl 0000,5 002,4
Thiamine HCl 0000,1 000,3
Glycine 0002 026,6
Sukrosa 30000
10
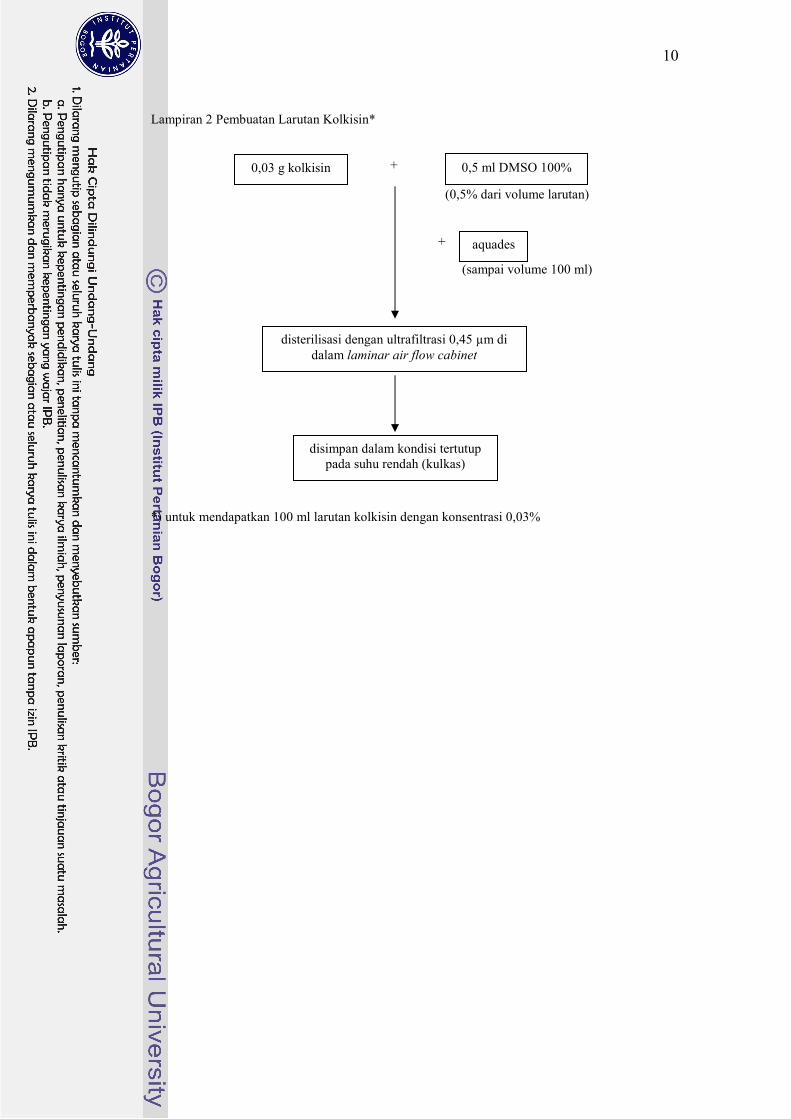
Lampiran 2 Pembuatan Larutan Kolkisin*
+
+
*) untuk mendapatkan 100 ml larutan kolkisin dengan konsentrasi 0,03%
(sampai volume 100 ml)
0,03 g kolkisin
(0,5% dari volume larutan)
0,5 ml DMSO 100%
aquades
disterilisasi dengan ultrafiltrasi 0,45 µm di
dalam laminar air flow cabinet
disimpan dalam kondisi tertutup
pada suhu rendah (kulkas)
11
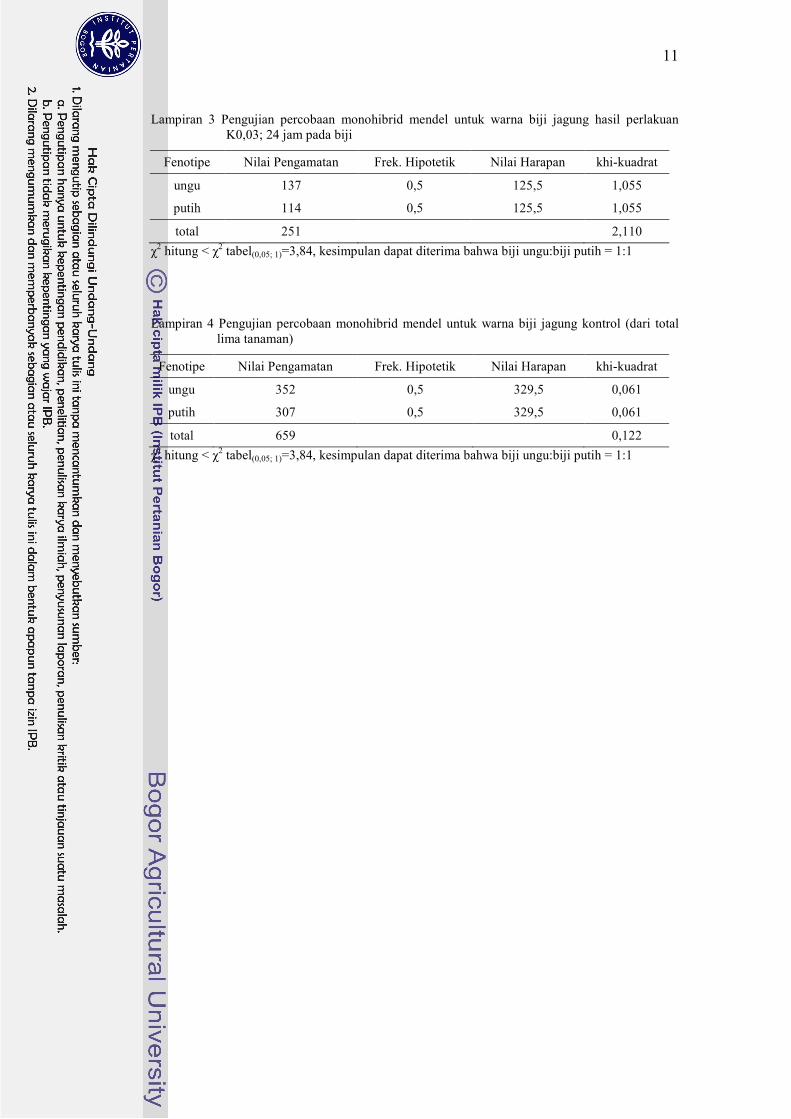
Lampiran 3 Pengujian percobaan monohibrid mendel untuk warna biji jagung hasil perlakuan
K0,03; 24 jam pada biji
Fenotipe Nilai Pengamatan Frek. Hipotetik Nilai Harapan khi-kuadrat
ungu 137 0,5 125,5 1,055
putih 114 0,5 125,5 1,055
total 251
2,110
χ2 hitung < χ
2 tabel(0,05; 1)=3,84, kesimpulan dapat diterima bahwa biji ungu:biji putih = 1:1
Lampiran 4 Pengujian percobaan monohibrid mendel untuk warna biji jagung kontrol (dari total
lima tanaman)
Fenotipe Nilai Pengamatan Frek. Hipotetik Nilai Harapan khi-kuadrat
ungu 352 0,5 329,5 0,061
putih 307 0,5 329,5 0,061
total 659
0,122
χ2 hitung < χ
2 tabel(0,05; 1)=3,84, kesimpulan dapat diterima bahwa biji ungu:biji putih = 1:1




















