
2. TINJAUAN PUSTAKA - repository.ipb.ac.id · L. fermentum LBP usus ayam dewasa ... karakterisasi...
31
7 2. TINJAUAN PUSTAKA Bakteri Asam Laktat BAL ditemukan pertama kali oleh Pasteur, seorang profesor kimia di University of Lille pada tahun 1878. Pada tahun 1889, Tissier, peneliti Prancis menemukan bakteri yang mendominasi saluran usus bayi yang minum air susu ibu yaitu Bifidobacterium. BAL berbentuk bulat maupun batang, Gram positif dan (dengan sedikit perkecualian) tidak motil, katalase negatif, tidak mempunyai sitokrom, aerotoleran, anaerobik hingga mikroaerofolik, serta membutuhkan nutrisi yang kompleks seperti asam amino, vitamin (B 1 , B 6 , B 12 dan biotin), purin dan pirimidin (Surono 2004). Walaupun BAL dapat hidup dengan dan tanpa oksigen, sumber energi terbesarnya untuk tumbuh adalah fermentasi gula. Bakteri ini mempunyai kapasitas respirasi yang sangat terbatas dan tidak dapat memperoleh ATP dari proses respirasi (Salminen & Wright 2004). BAL dibagi menjadi tiga grup berdasarkan pola fermentasinya, yaitu : a. Grup I : BAL homofermentatif obligatif, yang mengubah heksosa menjadi asam laktat melalui jalur Embden-Meyerhof, namun tidak bisa memfermentasikan pentosa ataupun glukonat. BAL grup ini termasuk dalam termobakterium, yang kekurangan glukosa-6 fosfat dehidrogenase dan 6-fosfoglukonat. Sebagian besar BAL grup ini tumbuh pada suhu 45 ○ C namun tidak tumbuh pada suhu 15 °C (Hopzapfel 1998). b. Grup II : BAL heterofermentatif fakultatif, yang memfermentasikan heksosa secara homofermentatif namun sebagian galur pada beberapa kondisi mempunyai metabolisme heterofermentatif dari heksosa menjadi asam laktat, karbondioksida dan ethanol atau asam asetat. Produksi asam asetat terjadi jika NAD + dapat diregenerasi tanpa pembentukan ethanol, misalnya melalui reduksi fruktosa atau molekul oksigen. Pentosa difermentasi melalui fosfoketolase menjadi asam laktat dan asam asetat. BAL grup ini termasuk dalam streptobakterium, yang mempunyai dua enzim dehidrogenase tetapi menggunakan jalur Embden-Meyerhof untuk fermentasi glukosa (Hopzapfel 1998).
-
Upload
vuonghuong -
Category
Documents
-
view
229 -
download
0
Transcript of 2. TINJAUAN PUSTAKA - repository.ipb.ac.id · L. fermentum LBP usus ayam dewasa ... karakterisasi...

7
2. TINJAUAN PUSTAKA
Bakteri Asam Laktat
BAL ditemukan pertama kali oleh Pasteur, seorang profesor kimia di
University of Lille pada tahun 1878. Pada tahun 1889, Tissier, peneliti Prancis
menemukan bakteri yang mendominasi saluran usus bayi yang minum air susu ibu
yaitu Bifidobacterium. BAL berbentuk bulat maupun batang, Gram positif dan
(dengan sedikit perkecualian) tidak motil, katalase negatif, tidak mempunyai
sitokrom, aerotoleran, anaerobik hingga mikroaerofolik, serta membutuhkan
nutrisi yang kompleks seperti asam amino, vitamin (B1, B6, B12 dan biotin), purin
dan pirimidin (Surono 2004). Walaupun BAL dapat hidup dengan dan tanpa
oksigen, sumber energi terbesarnya untuk tumbuh adalah fermentasi gula. Bakteri
ini mempunyai kapasitas respirasi yang sangat terbatas dan tidak dapat
memperoleh ATP dari proses respirasi (Salminen & Wright 2004).
BAL dibagi menjadi tiga grup berdasarkan pola fermentasinya, yaitu :
a. Grup I : BAL homofermentatif obligatif, yang mengubah heksosa menjadi
asam laktat melalui jalur Embden-Meyerhof, namun tidak bisa
memfermentasikan pentosa ataupun glukonat. BAL grup ini termasuk
dalam termobakterium, yang kekurangan glukosa-6 fosfat dehidrogenase
dan 6-fosfoglukonat. Sebagian besar BAL grup ini tumbuh pada suhu
45 ○C namun tidak tumbuh pada suhu 15 °C (Hopzapfel 1998).
b. Grup II : BAL heterofermentatif fakultatif, yang memfermentasikan
heksosa secara homofermentatif namun sebagian galur pada beberapa
kondisi mempunyai metabolisme heterofermentatif dari heksosa menjadi
asam laktat, karbondioksida dan ethanol atau asam asetat. Produksi asam
asetat terjadi jika NAD+ dapat diregenerasi tanpa pembentukan ethanol,
misalnya melalui reduksi fruktosa atau molekul oksigen. Pentosa
difermentasi melalui fosfoketolase menjadi asam laktat dan asam asetat.
BAL grup ini termasuk dalam streptobakterium, yang mempunyai dua
enzim dehidrogenase tetapi menggunakan jalur Embden-Meyerhof untuk
fermentasi glukosa (Hopzapfel 1998).

8
c. Grup III : BAL heterofermentatif obligatif, yang memfermantasikan
heksosa menjadi asam laktat, karbondioksida dan etanol atau asam asetat,
jika terdapat akseptor elektron alternatif. Pentosa diubah menjadi asam
laktat dan asam asetat. BAL grup ini termasuk dalam betabakterium, yang
kekurangan fruktosa 1.6 difosfat aldolase (Hopzapfel 1998).
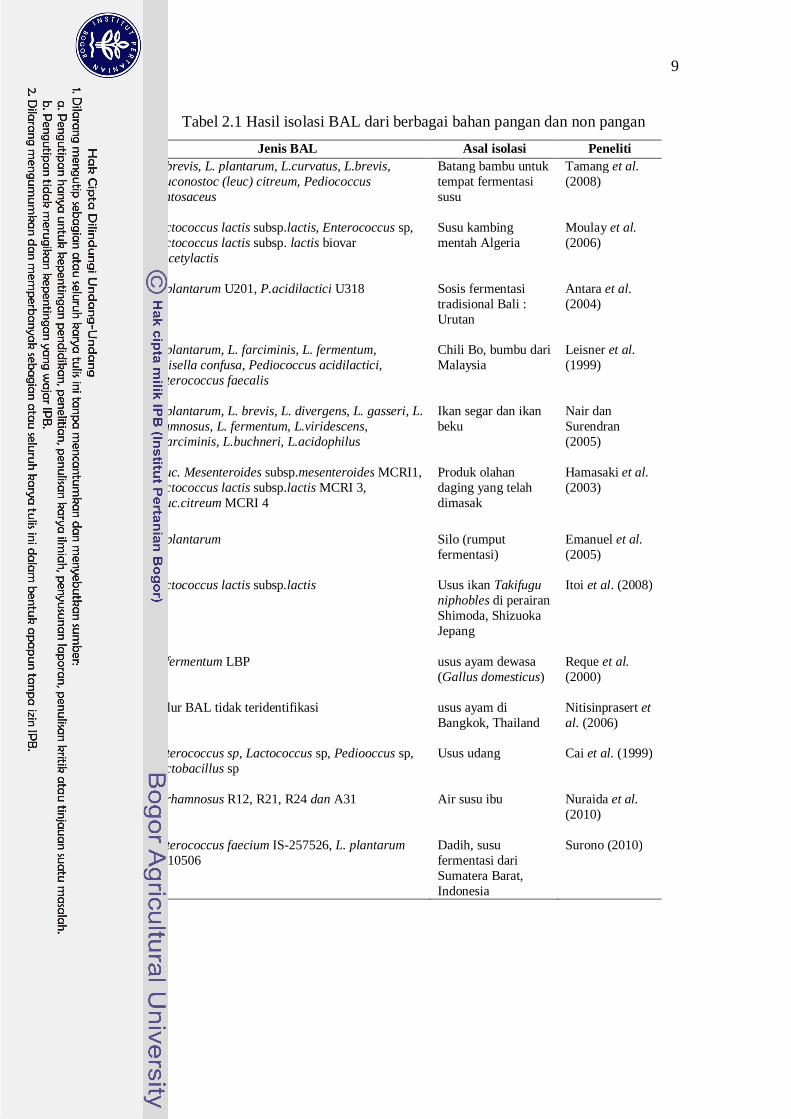
BAL sering ditemukan secara alamiah dalam bahan pangan. Bakteri ini
hidup pada susu, daging segar, dan sayur-sayuran. Pada proses fermentasi daging
spontan, BAL yang berasal dari bahan mentah atau lingkungan menyebabkan
terbentuknya asam laktat dari penggunaan karbohidrat, maupun rendahnya nilai
pH (5.9 sampai 4.6) (Surono 2004). Beberapa peneliti berhasil mengisolasi BAL
dari berbagai bahan pangan dan non-pangan termasuk di antaranya dari saluran
pencernaan (Tabel 2.1).
Lactobacillus spp. merupakan genus terbesar dari kelompok BAL
(Axelsson 1993). Genus Lactobacillus bersifat Gram positif dan tidak membentuk
spora, bersifat anaerob fakultatif, tumbuh optimum pada kisaran suhu 30-40 °C
tapi dapat tumbuh pada kisaran 5-35 °C. Lactobacillus tumbuh optimum pada pH
5.5-5.8, namun secara umum dapat tumbuh pada pH kurang dari 5. Lactobacillus
spp. banyak terdapat pada produk makanan fermentasi seperti produk-produk susu
fermentasi (yoghurt, keju, yakult) produk fermentasi daging seperti sosis
fermentasi, serta produk fermentasi sayuran seperti pikel dan sauerkraut.
Lactobacillus spp. berkontribusi untuk pengawetan, ketersediaan nutrisi, dan
flavor pada produk fermentasi tersebut (Salminen & Wright 2004).
9
Tabel 2.1 Hasil isolasi BAL dari berbagai bahan pangan dan non pangan Jenis BAL Asal isolasi Peneliti
L. brevis, L. plantarum, L.curvatus, L.brevis, Leuconostoc (leuc) citreum, Pediococcus pentosaceus
Batang bambu untuk tempat fermentasi susu
Tamang et al. (2008)
Lactococcus lactis subsp.lactis, Enterococcus sp, Lactococcus lactis subsp. lactis biovar diacetylactis
Susu kambing mentah Algeria
Moulay et al. (2006)
L. plantarum U201, P.acidilactici U318 Sosis fermentasi tradisional Bali : Urutan
Antara et al. (2004)
L. plantarum, L. farciminis, L. fermentum, Weisella confusa, Pediococcus acidilactici, Enterococcus faecalis
Chili Bo, bumbu dari Malaysia
Leisner et al. (1999)
L. plantarum, L. brevis, L. divergens, L. gasseri, L. rhamnosus, L. fermentum, L.viridescens, L.farciminis, L.buchneri, L.acidophilus
Ikan segar dan ikan beku
Nair dan Surendran (2005)
Leuc. Mesenteroides subsp.mesenteroides MCRI1, Lactococcus lactis subsp.lactis MCRI 3, Leuc.citreum MCRI 4
Produk olahan daging yang telah dimasak
Hamasaki et al. (2003)
L. plantarum Silo (rumput fermentasi)
Emanuel et al. (2005)
Lactococcus lactis subsp.lactis Usus ikan Takifugu niphobles di perairan Shimoda, Shizuoka Jepang
Itoi et al. (2008)
L. fermentum LBP usus ayam dewasa (Gallus domesticus)
Reque et al. (2000)
Galur BAL tidak teridentifikasi usus ayam di Bangkok, Thailand
Nitisinprasert et al. (2006)
Enterococcus sp, Lactococcus sp, Pediooccus sp, Lactobacillus sp
Usus udang Cai et al. (1999)
L. rhamnosus R12, R21, R24 dan A31 Air susu ibu Nuraida et al. (2010)
Enterococcus faecium IS-257526, L. plantarum IS-10506
Dadih, susu fermentasi dari Sumatera Barat, Indonesia
Surono (2010)

10
Probiotik
FAO/WHO (2002) telah mengeluarkan panduan untuk mengevaluasi
probiotik dalam makanan. Working Group yang dibentuk oleh FAO/WHO
menetapkan secara rinci panduan dan kriteria rekomendasi serta metodologi yang
digunakan untuk evaluasi probiotik, mengidentifikasi serta menentukan data-data
yang dibutuhkan untuk mengklaim kesehatan probiotik. Kriteria pertama yang
harus dipenuhi adalah bahwa galur yang didapatkan harus diketahui
identifikasinya, baik secara fenotipik maupun genotipik, mulai dari genus sampai
spesies bahkan sampai tingkatan sub spesies. Kriteria selanjutnya adalah
karakterisasi fungsional, baik secara in vitro maupun in vivo, kemudian
dilanjutkan dengan pengujian keamanan secara in vitro dan in vivo, serta studi
fase satu di manusia. Untuk evaluasi pangan probiotik dilakukan studi fase dua di
manusia yaitu secara double blind, acak, kontrol placebo (DBPC) atau desain lain
yang sesuai dengan ukuran sampel dan dampak primer untuk menentukan jika
galur/produk berpotensi efikasi dan dikonfirmasi kembali hasil yang didapatkan.
Fase tiga dilakukan untuk menguji efektivitas probiotik pada kondisi spesifik.
Kemudian dilakukan aplikasi probiotik ke dalam pangan probiotik yang harus
memenuhi syarat pelabelan antara lain pencantuman isi : genus, spesies, galur;
jumlah minimum bakteri hidup pada akhir masa simpan, kondisi penyimpanan
yang sesuai, serta kontak detil perusahaan untuk informasi konsumen (FAO/WHO
2002).
Di samping itu, beberapa peneliti lain telah mengemukakan jaminan
kriteria untuk bakteri probiotik. Beberapa di antaranya adalah sebagai berikut :
a. Probiotik harus dapat bertahan melewati lambung dan usus halus, sehingga
probiotik harus toleran terhadap suasana asam dan adanya asam empedu
(Tuomola et al. 2001, Bourlioux et al. 2003, Roberfroid 2000; Sunny-
Roberts & Knoor 2008).
b. Probiotik harus mempunyai kemampuan dalam melakukan penempelan ke
usus (Nitisinprasert et al. 2006; Tuomola et al. 2001; Bourlioux et al.
2003), karena sangat berkaitan dengan beberapa efek kesehatan antara lain
pemendekan lama diare, efek imunologik dan eksklusi kompetitif dengan

11
mikroba patogen (Tuomola et al. 2001; Herick & Levkut 2002; Bourlioux
et al. 2003).
c. Probiotik harus mampu bertahan selama proses pengolahan dan
penyimpanan (FAO/WHO 2002; Sunny-Roberts & Knoor 2008), sehingga
saat dikonsumsi masih mempunyai viabilitas yang cukup untuk mengatasi
berbagai masalah di saluran pencernaan antara lain dengan meningkatkan
pencernaan laktosa, mengontrol infeksi di usus, serta menjaga
keseimbangan barier mukosa usus (Tuomola et al. 2001; Agostoni et al.
2004).
Probiotik mempunyai efek kesehatan bagi manusia diantaranya (1)
menurunkan risiko lactose intolerance, (2) mengurangi kejadian diare (Reid 1999;
Talwalkar & Kailasapathy 2004), (3) menurunkan jumlah enzim mikrobial fekal
seperti β-glukoronidase, β-nitroreduktase, nitroreduktase dan urease yang
berperan dalam aktivasi mutagenesis dan karsinogenesis di kolon, serta berperan
dalam efek hipokolesterolemik (Roberfroid 2000), (4) meningkatkan respon
sistem imun (Perdigon et al. 2001; Bhatia & Rani 2008), (5) menurunkan risiko
penyakit kardiovaskuler pada perokok (Naruszewics et al. 2002), dan (6) bersifat
hipokolesterolemik (Kusumawati 2002; Kimoto-Nira et al. 2007). Kusumawati
(2002) melaporkan bahwa pemberian susu yang difermentasi oleh BAL
Lactobacillus acidophilus FNCC 116, Lactobacillus plantarum sa28k dan
Lactobacillus casei FNCC262 mampu menurunkan kadar kolesterol darah tikus
percobaan.
Beberapa BAL mampu mencegah terjadinya kanker kolon. BAL mampu
mereduksi level enzim di kolon yang mengubah prokarsinogen menjadi
karsinogen. Secara spesifik, BAL dapat mereduksi level enzim -glukuronidase,
nitroreduktase dan azoreduktase. BAL juga berperan secara langsung dalam
mereduksi prokarsinogen, contohnya dengan mengikat nitrit dan mereduksi level
asam empedu sekunder (BC Dairy Foundation 1997; Burns & Rowlands 2000;
Brady et al. 2000; Wollowski et al. 1999).
Mekanisme yang terjadi sehingga BAL sangat bermanfaat untuk kesehatan
adalah sebagai berikut :

12
a. Mengikat karsinogen
Beberapa penelitian menunjukkan bahwa BAL mampu mengikat secara in
vitro senyawa karsinogen, di antaranya adalah heterosiklik amin selama
pemasakan daging, toksin fungi aflatoksin B1, benzo(a)pirene dan
makanan yang terkontaminasi aflatoksin AF2. Adsorpsi itu dikaitkan
dengan mekanisme pertukaran kation (Burns & Rowlands 2000).
b. Efek pada enzim bakteri dan produksi metabolit
Peningkatan konsentrasi BAL dalam setiap konsumsi mampu menurunkan
enzim bakteri yang berperanan dalam aktivasi atau sintesis karsinogen,
genotoksin dan promotor tumor (Burns & Rowlands 2000).
c. Stimulasi enzim protektif
Beberapa karsinogen seperti heterosiklik amin dan PAH (Polycyclic
Aromatic Hydrocarbon) dikenal dapat mengkonjugasikan glutation
sehingga menjadi tidak aktif. Enzim glutation transferase (GSH) berada
di hati dan jaringan lain termasuk saluran pencernaan. BAL mampu
meningkatkan enzim GSH (Wollowksi et al. 2001; Burns & Rowlands
2000).
d. Menurunkan pH kolon
Fermentasi yang dilakukan oleh probiotik mampu menurunkan pH kolon
dengan terbentuknya asam-asam organik seperti asam laktat dan SCFA
(Short Chain Fatty Acid) diantaranya asam butirat, propionat dan asam
asetat sehingga mampu menjaga kondisi kolon dari resiko kanker
(Augenlicht et al. 1999; Wollowski et al. 1999).
e. Meningkatkan respon imun
Probiotik mampu menurunkan respon inflamasi. Hal ini ditunjukkan
dengan studi mencit yang disuntikkan dengan sel tumor dan diberikan
yoghurt. Hal tersebut mampu menekan terjadinya inflamasi dengan
meningkatkan imunoglobin A (IgA) dan sel limfosit T CD 4+. Selain itu
juga, studi pada manusia menunjukkan bahwa konsumsi probiotik mampu
meningkatkan aktivitas fagositik dari sel imun monosit dan granulosit
serta meningkatkan level antibodi yang disekresikan oleh sel imun limfosit
B (Burns & Rowlands 2000; Perdigon et al. 2001). Solis et al. (2002)

13
melaporkan bahwa pemberian susu fermentasi yoghurt dapat
meningkatkan produksi interferon IFN-γ pada anak-anak yang kekurangan
gizi. Beberapa peneliti melaporkan hasil penelitiannya yang membuktikan
bahwa konsumsi BAL mampu meningkatkan sistem imun seluler dan
humoral di antaranya peningkatkan populasi dan proliferasi sel limfosit,
produksi sitokin interferon-γ (IFN- γ), interleukin-12 (IL-12), IL-10, sel
imun Th, serta IgA, IgE, IgG, serta IgM (Kimura et al. 2006; Segawa et al.
2008; Gackowska et al. 2006; Aattouri et al. 2002).
Ketahanan Bakteri Asam Laktat pada pH Rendah
Roberfroid (2000) menyatakan bahwa probiotik adalah sel mikroba hidup
yang dikonsumsi oleh manusia, merupakan mikroflora yang dapat hidup di
saluran pencernaan dan mempunyai efek yang menguntungkan bagi kesehatan
manusia. Setelah dapat melewati lambung dan usus halus, bakteri yang termasuk
dalam probiotik dapat bertahan hidup di usus besar. Adanya kapasitas fermentasi
pada kolon memungkinkan bakteri probiotik berkembang biak, dan hasilnya,
sering ditemukan sejumlah probiotik yang terikut dalam feses. Oleh karenanya
salah satu syarat bakteri termasuk dalam probiotik adalah mampu bertahan hidup
pada kondisi sesuai saluran pencernaan yang meliputi keasaman yang tinggi dan
adanya sekresi garam empedu.
Cotter dan Hill (2003) melaporkan mekanisme homeostatik instrinsik yang
menyebabkan BAL mampu bertahan pada kondisi pH rendah atau keasaman yang
tinggi. Mekanisme yang terjadi pada BAL di antaranya adalah sistem Glutamat-
dekarboksilase (GAD), sistem arginin deiminasi (ADI) dan pompa proton H+ -
ATP ase.
Sistem glutamat dekarboksilase merupakan suatu mekanisme pertahanan
sel sebagian BAL terhadap kondisi pH yang rendah. Beberapa spesies
Lactobacillus sp dilaporkan memiliki mekanisme sistem GAD. Adapun
mekanisme sistem GAD adalah sebagai berikut : setelah mengkonsumsi glutamat
melalui suatu transporter spesifik, terjadi dekarboksilasi glutamat di dalam
intraseluler, menjadi produk γ – aminobutyrat (GABA) yang dikeluarkan dari
dalam sel oleh suatu antiporter, sehingga terjadi peningkatan pH intraseluler

14
(Cotter & Hill 2003). Untuk lebih jelas, mekanisme sistem GAD untuk
mempertahankan pH intraseleluler bakteri dapat dilihat pada Gambar 2.1.
a. b
Gambar 2.1 Mekanisme Sistem Glutamat Dekarboksilasi (Cotter & Hill 2003)
Gambar 2.1.a menjelaskan mekanisme homeostatis sel BAL terhadap pH
rendah. Mekanisme ini meliputi adanya perpindahan ion glutamat ke dalam sel
dan produk GABA keluar sel yang diiringi oleh malat dekarboksilasi, ion malat
masuk ke dalam sel dan ion laktat keluar sel serta oksaloasetat dekarboksilasi, ion
laktat keluar sel dan ion sitrat masuk ke dalam sel mengakibatkan terjadinya
perbedaan potensial elektrogenik dan meningkatkan kondisi alkali sitoplasma.
Gambar 2.1.b menjelaskan bahwa sistem GAD berhubungan dengan pompa
proton serta jalur F1F0-ATPase sebagai transporter ion-ion glutamat, malat dan
sitrat, serta produk katabolismenya.
Keberadaan glutamat sebagai zat yang dapat mempertahankan kondisi
homeostatis pH internal sel BAL menjadi faktor yang penting untuk perlindungan
BAL terhadap kondisi pH rendah. Glutamat dapat diperoleh dari berbagai sumber
terutama dari makanan pembawa BAL misalnya makanan kaya protein (susu dan
daging), beberapa buah-buahan, ataupun bahan kriogenik yang sengaja
Pompa proton

15
ditambahkan untuk pengawetan BAL selama pengolahan; misalnya monosodium
L-glutamat monohidrat (MSG),
Arginin deiminasi (ADI) sistem merupakan suatu mekanisme homeostatis
terhadap kondisi pH rendah yang dimiliki beberapa BAL seperti Lactobacillus
casei dan Lactobacillus sanfranciscensis. Kedua bakteri tersebut dapat
mengkatabolisme arginin menjadi ornithin, amonia, dan CO2. Amonia (NH4)
akan meningkatkan pH internal sitoplasma. Hal ini menjadikan kedua bakteri
tersebut dapat menyesuaikan hidupnya pada kondisi pH yang rendah. Sistem ADI
ini dikendalikan oleh gen arcA, arcB, arc, dan arcT, sehingga BAL yang tidak
mempunyai gen tersebut, tidak memiliki mekanisme homeostatis sistem arginin
deiminasi untuk bertahan pada pH rendah (Cotter & Hill 2003).
Marteau et al. (1997) melakukan penelitian untuk mengetahui ketahanan
hidup BAL pada kondisi saluran pencernaan melalui model dinamis in vitro
(gerakan peristaltik, perubahan pH, perubahan konsentrasi enzim dan garam
empedu di saluran pencernaan). BAL yang dipakai adalah galur tunggal
Lactobacillus acidophilus, Lactobacillus bulgaricus, Bifidobacterium bifidum dan
Streptococcus thermophilus. Model saluran pencernaan yang dirancang terdiri
dari dua kondisi yang berbeda yaitu simulasi sekresi fisiologis empedu dan sekresi
empedu yang rendah.
Streptococcus thermophilus dan Lactobacillus bulgaricus dalam produk
yogurt hanya mampu bertahan selama 20 menit di kompartemen lambung,
sedangkan Lactobacillus acidophilus dan Bifidobacteria bifidum dalam produk
Ofilus® lebih tahan dibandingkan produk yogurt. Bakteri dalam yogurt akan
mengalami kematian (populasi nol) setelah 110 menit berada dalam kompartemen
lambung, sedangkan produk Ofilus® mengalami kematian setelah 180 menit.
Setelah memasuki kompartemen usus, BAL dan bifidobakteria mampu hidup dan
melakukan pertumbuhan dan proliferasi, sehingga populasinya meningkat setelah
2 jam berada di kompartemen usus. Peningkatan L. acidophilus dan B. bifidum
pada produk Ofilus® lebih tinggi yaitu 60-70% dari populasi saat konsumsi
dibandingkan dengan peningkatan populasi L. bulgaricus dan S. thermophilus
pada produk yogurt yang mengalami peningkatan populasi maksimal sebesar 10-
20%. Hal ini membuktikan bahwa secara alami, bakteri L. acidophilus dan B.

16
bifidum lebih tahan terhadap lingkungan saluran pencernaan yang meliputi
kondisi pH rendah, sekresi pankreatik, garam empedu, serta adanya bikarbonat,
dibandingkan dengan L. bulgaricus dan S. thermophilus (Martaeu et al. 1997).
Lin et al. (2006) juga melakukan penelitian untuk menguji sifat-sifat
probiotik beberapa produk komersial yogurt, produk granul dan bubuk yang
beredar di pasaran yang telah diklaim mengandung BAL probiotik. Sifat-sifat
probiotik yang diuji adalah sifat ketahanan hidup pada kondisi pH rendah dan
garam empedu serta uji penempelan pada sel epitel usus. Hasil penelitiannya
menunjukkan bahwa pada produk komersial yang mengandung BAL terdapat
penurunan jumlah populasi bakteri yang mampu bertahan hidup pada pH 2.0.
Populasi BAL pada produk cair sekitar 105 cfu/ml pada pH 2.0, yang lebih tinggi
dibandingkan dengan produk bubuk yaitu 104 cfu/g. Galur BAL yang mampu
bertahan hidup pada pH 2.0 juga mampu bertahan hidup pada kondisi garam
empedu 0.3%.
Zoumpopoulou et al. (2008) juga melakukan penelitian untuk menguji
ketahanan BAL pada kondisi yang disimulasikan seperti kondisi pada saluran
pencernaan manusia. Ketahanan hidup galur probiotik pada larutan PBS
(Phosphat Buffer Saline) pH 2.5 ditentukan setelah inkubasi pada suhu 370C
selama 0.5, 1, 2 dan 4 jam seperti masa transit makanan di dalam lambung.
Toleransi dalam garam empedu ditentukan dengan ketahanan hidup probiotik
pada larutan PBS pH 8 yang mengandung 1% dan 2% (w/v) oxgall (Ox-Bile,
LP0055, Oxoid) setelah inkubasi selama 1, 2 dan 4 jam seperti masa transit
makanan dalam usus halus.
Ketahanan BAL terhadap Garam Empedu
Empedu merupakan cairan kuning kehijauan yang terdiri dari asam
empedu, kolesterol, phospolipid dan pigmen biliverdin. Asam empedu primer
(asam kholat dan asam khenodeoksikholat) disintesis di hati dari kolesterol, yang
dapat dimodifikasi oleh enzim bakteri pada usus menjadi bentuk sekunder (asam
deoksikholat dan asam lithokholat). Semua asam empedu berkonjugasi dengan
glisin atau taurin sebelum disekresikan. Grup karboksil dari asam empedu dan
grup amino dari asam amino terikat dengan ikatan amida (ikatan peptida)

17
(Gambar 2.2). Garam empedu disintesis dalam hepatosit perisentral dalam hati,
disimpan dan dikonsentrasikan pada kantong empedu dan dilepaskan ke
duodenum setelah ada makanan masuk ke usus halus (Begley et al. 2005)
Gambar 2.2 Struktur kimia asam empedu a) kolesterol, b) ikatan peptida pada garam empedu, c) misel (Begley et al. 2005)
Fungsi biologis garam empedu adalah sebagai detergen yang
mengemulsifikasi dan melarutkan lipid. Namun selain itu, garam empedu juga
dapat berfungsi sebagai antimikroba yaitu melalui perusakan membran sel bakteri
(Begley et al. 2005). Hal inilah yang menyebabkan isolat BAL mengalami
penurunan populasi pada kondisi media yang dipapar oleh 0.5% garam empedu.
Bron et al. (2004) menggambarkan kerusakan secara morfologis L.plantarum
pada kondisi dipapar garam empedu 0.05% sampai 0.15% selama 4 jam.
Kerusakan dinding sel diamati di bawah Scanning Electron Microscope (SEM),
semakin tinggi persentase garam empedu, maka kerusakan dinding sel bakteri
semakin besar dan akhirnya sel lisis.
Terdapat dua hipotesis yang menjelaskan BAL mampu bertahan pada
kondisi garam empedu. Hipotesis pertama adalah beberapa spesies BAL mampu
mendekonjugasi garam empedu dengan menggunakan asam amino taurin sebagai
kolesterol
Ikatan peptida
misel

18
akseptor elektron. Hipotesis kedua menyatakan bahwa BAL mampu bertahan
pada kondisi garam empedu karena sebagian besar galur BAL mempunyai enzim
Bile Salt Hydrolase (BSH) yang diatur oleh gen bsh. Hipotesis kedua ini lebih
banyak dibuktikan oleh beberapa peneliti daripada hipotesis pertama (Moser &
Savage 2001). L. plantarum WCFS1, L. johnsonii NCC533, B. longum
NCC2705, L. acidophilus NCFM ATCC 700396, L. brevis ATCC 367 dan L.
gasseri ATCC 33323 mempunyai gen bsh yang terdiri atas 255-338 asam amino.
L. plantarum mempunyai aktivitas enzim BSH berdasarkan penelitian yang
dilakukan oleh Begley et al. (2006), yang membuktikan bahwa pada media MRS
yang disuplementasi garam empedu dan diinokulasikan L. plantarum terjadi
presipitasi asam empedu tidak terkonjugasi. Enzim BSH menguraikan asam
empedu terkonjugasi menjadi asam empedu tidak terkonjugasi dan melepaskan
asam amino glisin atau taurin (Gambar 2.3).
Gambar 2.3 Peranan enzim BSH dalam menguraikan asam empedu: a) asam empedu, b) peranan enzim BSH, c) visualisasi presipitasi asam empedu di media MRS (Begley et al. 2006)
Aktivitas Antimikroba BAL
Kelompok BAL merupakan bakteri yang menghasilkan metabolit primer
berupa asam laktat. Peran lain dari BAL adalah mampu meningkatkan keamanan
pangan dengan cara menghambat pertumbuhan bakteri pembusuk makanan dan
bakteri patogen, baik bakteri Gram positif maupun Gram negatif (Robredo &
Torres 2000; El-Naggar 2004). Penghambatan yang dilakukan oleh BAL terhadap
mikroorganisme yang lainnya dimungkinkan karena BAL menghasilkan produk

19
metabolit yang bersifat antimikroba antara lain diasetil, hidrogen peroksida, asam-
asam organik dan bakteriosin (Jenie & Rini 1995; Surono 2004; Helander et al.
1997; Naidu & Clemens 2000).
Kemampuan BAL dalam menghasilkan senyawa antimikroba dilaporkan
oleh beberapa peneliti. Nowroozi et al. (2004) menyatakan bahwa L. plantarum
mempunyai aktivitas antimikroba lebih besar terhadap S. aureus dan E. coli
dibandingkan dengan beberapa BAL lainnya, seperti Lactobacillus brevis,
Lactobacillus casei, Lactobacillus delbruekii dan Lactobacillus acidophilus.
Toksoy et al. (1999) menyatakan bahwa L. plantarum AX5L yang diisolasi dari
sosis dapat menghambat E. coli, S. aureus dan B. subtilis karena L. plantarum
AX5L mampu menghasilkan H2O2, asam laktat sebesar 0.88% dan bakteriosin
plantarisin. Streptococcus lactis memiliki aktivitas bakterisidal terhadap bakteri
Gram positif maupun Gram negative, antara lain Enterococcus faecalis, Bacillus
subtilis, Salmonella typhimurium dan Eschericia coli (Suarsana et al. 2001).
Bakteriosin
Sejumlah galur BAL secara alami mampu menghasilkan substansi protein,
biasanya memiliki bobot molekul yang kecil yang mampu menghambat bakteri
lain, secara umum substansi ini dikenal dengan nama bakteriosin. Bakteriosin
mempunyai aktivitas antimikroba terhadap patogen pencemar makanan
(foodborne) dan organisme berspora lainnya (Tannock 1999; Karaoglu et al.
2003; Bromberg et al. 2004).
Bakteriosin diproduksi oleh BAL yang menguntungkan bagi kesehatan
manusia yang termasuk dalam GRAS yang merupakan pendekatan baru untuk
mengontrol mikroba patogen dalam bahan pangan. Bakteriosin merupakan
molekul protein atau peptida ekstraseluler yang mempunyai aksi bakterisidal atau
bakteriostatik terhadap bakteri yang mempunyai kekerabatan dekat. Bakteriosin
tersebut dapat didegradasi oleh enzim protease dalam saluran pencernaan.
Bakteriosin bersifat irrevesible, mudah dicerna, berpengaruh positif terhadap
kesehatan dan aktif pada konsentrasi rendah (Savadogo et al. 2006; Meghrous et
al. 1997).

20
Berdasarkan karakteristiknya, bakteriosin dapat dikelompokkan menjadi
empat kelas yaitu kelas I adalah grup lantibiotik (modified bacteriocins) di
antaranya nisin, lactococin, lacticin, carnocin dan cytolysin; kelas II adalah
bakteriosin yang mempunyai berat molekul rendah (< 10 kDa), tahan panas 100-
1210C; kelas III adalah bakteriosin yang mempunyai berat molekul tinggi (> 30
kDa) dan bersifat tidak tahan panas, serta kelas IV yaitu kompleks bakteriosin,
proteinnya berikatan dengan lipid dan atau karbohidrat (Karaoglu et al. 2003;
Savadogo et al. 2006).
Setiap bakteriosin mempunyai reseptor spesifik sel sasaran, dan memiliki
cara kerja yang berbeda-beda dalam menghambat sel sasaran, antara lain sebagai
berikut: (1) mengganggu metabolisme sel mikroba, (2) menghambat sintesis
dinding sel mikroba, (3) mengganggu keutuhan membran sel mikroba, (4)
menghambat sintesis protein sel mikroba yang berlangsung di ribosom, dan (5)
menghambat sintesis asam nukleat sel mikroba. Bakteriosin terlebih dahulu
masuk ke dalam sel sasarannya, melewati dinding atau membran sitoplasma agar
dapat masuk atau teradsorpsi ke dalam sel sasaran untuk menghambat bakteri
(Ogunbawo et al. 2003). Bakteriosin membentuk pori di membran sel yang
sensitif dan menurunkan potensial atau gradien pH yang menyebabkan rusaknya
material seluler. Efek penghambatan bakteriosin dipengaruhi oleh komposisi
fosfolipid pada galur bakteri target dan pH lingkungan (Albano et al. 2007; Pal et
al. 2005).
Bakteriosin kelas 1 yang dikenal dengan sebutan lantibiotik membentuk
pori pada dinding sel target dengan model ’wedge-like’, sedangkan bakteriosin
kelas II membentuk pori dengan sistem ’barrel-stave’ atau melalui mekanisme
’carpet’ (Gambar 2.4). Lantibiotik mengganggu susunan lapisan lipid bilayer
saat terikat pada membran sel. Masuknya lantibiotik ke dalam membran
dipromotori oleh gradien trans membran (∆ pH) atau trans negatif (∆ ψ). C-
terminal dari lantibiotik kemudian dapat masuk ke dalam membran, dan akhirnya
seluruh lantibioik dapat translokasi menembus membran sel, yang juga
dipengaruhi oleh ’proton motive force’. Mekanisme ini disebut dengan
pembentukan pori ’wedge-like pore’. Bakteriosin kelas II membentuk pori model
’barrel stave’. Adanya residu asam amino heliks di bagian tengah struktur

21
bakteriosin kelas II memulai insersi masuknya peptida ke dalam membran sel
target. Sisi hidrofilik dari ikatan peptida ampifatik α heliks akan membuka ikatan
asam lemak pada lipida membran. Akhirnya seluruh peptida bakteriosin dapat
menembus membran sel (Moll et al. 1999).
Gambar 2.4 Mekanisme pembentukan pori oleh bakteriosin (a) model ’wedge- like pore’, (b) model ’barrel-stave pore’ (Moll et al. 1999).
Asam Organik
Terbentuknya asam laktat dan asam organik oleh BAL dapat
menyebabkan penurunan pH, akibatnya mikroba yang tidak tahan terhadap
kondisi pH yang relatif rendah akan terhambat. Akumulasi produk akhir asam
yang rendah pH-nya menghasilkan penghambatan yang luas terhadap Gram
positif maupun Gram negatif (Naidu & Clemens 2000).
Efek penghambatan dari asam organik terutama berhubungan dengan
jumlah asam yang tidak terdisosiasi. Asam yang tidak terdisosiasi dapat berdifusi
secara pasif ke dalam membran sel. Di dalam sel, asam tersebut terdisosiasi
menjadi proton dan anion lalu mempengaruhi pH di dalamnya (Branen &
Davidson 1993; Jenie 1996). Pada kondisi asam, konstanta disosiasi (nilai pKa)
dan konsentrasi molar merupakan faktor penentu aktivitas penghambatan asam
laktat dan asam asetat. Asam asetat mempunyai aktivitas antimikroba yang lebih
luar sel
dalam sel
Asam
Basa

22
tinggi dibandingkan dengan asam laktat. Asam asetat mempunyai nilai pKa 4.756
sedangkan asam laktat mempunyai nilai pKa 3.860. Pada pH usus sekitar 5.8,
sebanyak 8.4% asam asetat dan 1.1% asam laktat berada pada kondisi tidak
terdisosiasi (Naidu & Clemens 2000).
Asam asetat dan asam laktat yang tidak terdisosiasi merupakan asam
lipofilik yang pada kondisi tidak terdisosiasi dapat melakukan penetrasi ke
membran sel dan pada pH intraseluler yang lebih tinggi akan terdisosiasi
memproduksi ion hidrogen dan dapat mengganggu fungsi metabolik seperti
translokasi substrat dan fosforilasi oksidatif. Beberapa penelitian menunjukkan
bahwa asam laktat dan asam asetat dapat menghambat Staphylococcus aureus
(Ratanapibulsawat et al. 2005). Salmonellae dihambat oleh asam laktat pada pH
lebih rendah dari 4.4 dan asam asetat pada pH 5.4 (Naidu & Clemens 2000).
Hidrogen Peroksida
Hidrogen peroksida (H2O2) merupakan oksidator, bleaching agent dan
anti bakteri. Hidrogen peroksida murni tidak berwarna, berbentuk cairan seperti
sirup, dan memiliki bau yang menusuk. Senyawa ini dapat terdekomposisi
menjadi air dan oksigen. Pada suhu ruang dekomposisi H2O2 berjalan lambat.
Perubahan kondisi lingkungan seperti pH dan suhu mempengaruhi kecepatan
H2O2 terdekomposisi. Dengan kenaikan suhu, keefisienan dalam menghancurkan
bakteri meningkat tetapi kecepatan terdekomposisinya juga semakin cepat
(Branen & Davidson 1993).
BAL memproduksi H2O2 (hidrogen peroksida) melalui transport elektron
via enzim flavin. Dengan adanya H2O2, bentuk anion superoksida merusak
radikal hidroksi. Proses antimikrobanya melibatkan peroksidase lipid membran
dan meningkatkan permeabilitas membran. Hasilnya adalah efek bakterisidal dari
metabolit oksigen yang mengakibatkan terjadinya oksidasi sel bakteri dan
akhirnya merusak asam nukleat dan protein sel (Naidu & Clemens 2000).
Fungsi H2O2 sebagai antimikroba tergantung pada kemampuan
oksidatifnya. Kemampuan untuk mengoksidasi menyebabkan perubahan tetap
pada sistem enzim sel mikroba. Kemampuan bakterisidal dari H2O2 beragam
tergantung pH, konsentrasi, suhu, waktu, dan tipe serta jumlah mikroorganisme.

23
Pada kondisi tertentu spora bakteri ditemukan paling resistan terhadap H2O2,
diikuti dengan bakteri Gram positif. Bakteri yang paling sensitif terhadap H2O2
adalah bakteri Gram negatif, terutama koliform (Branen & Davidson 1993).
Sifat Penempelan BAL pada Sel Epitel Usus
Selain sifat ketahanannya terhadap pH rendah dan garam empedu, sifat
ketahanan BAL sebagai probiotik di saluran pencernaan juga ditentukan dengan
uji penempelan. Berbagai model penempelan secara in vitro dilakukan antara lain
penempelan ke epitel usus babi oleh Kos et al. (2003), Mishra dan Prasad (2005)
yang melakukan uji penempelan Lactobacillus casei secara in vitro ke sel usus
tikus dan pemodelan penempelan ke stainless steel serta Blum et al. (1999) yang
menggunakan model penempelan BAL sebagai probiotik secara in vitro ke sel
Caco-2.
Penempelan bakteri pada permukaan epitel usus atau yang sering disebut
adhesi terjadi melalui beberapa mekanisme. Bakteri dapat berinteraksi dengan sel
epitel usus, matriks ekstraseluler dan lapisan mukus. Lapisan mukus yang
menutupi sel epitel merupakan kontak pertama di usus bagi mikroorganisme
untuk melakukan penempelan dan kolonisasi di usus. Jika mukosa rusak, maka sel
epitel usus merupakan tempat penempelan bakteri (Adlerberth et al. 2000). Mukus
terbentuk dari musin yang merupakan glikoprotein yang menyusun gel tersebut.
Karbohidrat penyusun musin adalah galaktosa, fruktosa, N-asetilglukosamin, N-
asetilgalaktosamin dan asam sialat. Karbohidrat tersebut tersusun pada beberapa
jenis struktur yang berbeda. Polisakarida yang terbentuk merupakan bagian yang
berinteraksi langsung dengan bakteri dan berfungsi pada proses pengenalan sel
(cell-cell recognition). Pada struktur glikoprotein yang melekat pada permukaan
membran sel, karbohidrat membentuk glikonjugat dengan protein melalui ikatan
nitrogen dengan gula selain manosa. Glikoprotein yang dapat menyebabkan
bakteri melekat, baik secara spesifik maupun secara alami, melalui suatu adhesin
yang dimiliki oleh bakteri (Bourlioux et al. 2003). Struktur sederhana
glikoprotein ditunjukkan pada Gambar 2.5.

24
Gambar 2.5 Struktur sederhana glikoprotein (Bourlioux et al. 2003)
Permukaan sel epitel usus diselimuti oleh lapisan yang bersifat visko-
elastik yang terdiri dari glikprotein. Gel mukosa dibentuk dari musin yang
disekresikan oleh sel goblet. Musin dan glikoprotein memegang peranan penting
pada proses penempelan bakteri. Glikoprotein merupakan sisi tempat penempelan
bakteri seperti ditunjukkan pada Gambar 2.6. Selain itu, musin dan glikoprotein
juga merupakan senyawa nutrien yang digunakan oleh bakteri untuk
pertumbuhannya. Setelah bakteri berhasil menempel dengan adanya interaksi
antara adhesin dengan reseptor yang berupa glikoprotein di sel epitel usus, maka
bakteri tersebut akan berkolonisasi dan memanfaatkan musin serta glikoprotein
lainnya untuk pertumbuhannya.
Gambar 2.6 Penempelan bakteri ke glikoprotein di sel epitel usus (Danone research team 2002)
Bakteri terikat pada reseptor
Reseptor glikokonjugat
Epitel usus usus
Bakteri

25
Adhesin merupakan suatu protein yang mengenali karbohidrat pada
glikoprotein membran sel usus (Ouwehand et al. 2001). Pada bakteri Gram
positif, adhesin berada di dinding sel. L. plantarum mengekspresikan adhesin
yang mengikat manosa yang lebih baik daripada spesies Lactobacillus lainnya.
Oleh karenanya L. plantarum cenderung memiliki sifat penempelan ke permukaan
usus yang baik (Adlerberth et al. 2000).
Kompetisi Lactobacillus dengan Bakteri Enteropatogen di Saluran
Pencernaan
Galur Lactobacillus sebagai probiotik mampu mencegah diare yang
disebabkan oleh bakteri enteropatogen khususnya EPEC melalui sejumlah
mekanisme. Lu & Walker (2001) menyatakan bahwa pencegahan diare yang
disebabkan oleh EPEC diawali dengan pencegahan translokasi EPEC ke sel epitel
usus oleh probiotik. Probiotik mampu berkompetisi dengan EPEC dalam
memanfaatkan nutrien penting dalam usus lalu mampu tumbuh dan mendapatkan
sisi penempelan pada sel epitel usus. Setelah berhasil menempel dan berkolonisasi
pada sel epitel usus, probiotik memproduksi dan mengeluarkan senyawa metabolit
antimikroba yang mampu menghambat pertumbuhan bakteri patogen ke usus.
Selain itu, sisi penempelan sel epitel usus yang telah digunakan oleh probiotik
menyebabkan EPEC tidak mampu menempel ke sel epitel usus. Kondisi inilah
yang menyebabkan bakteri probiotik mampu mencegah pertumbuhan EPEC
sehingga diare tidak terjadi. Untuk lebih jelasnya, kompetisi antara probiotik
dengan bakteri patogen dalam usus dapat dilihat pada Gambar 2.7.

26
Gambar 2.7 Kompetisi antara bakteri probiotik (flora normal) dan bakteri
patogen di usus (Lu & Walker 2001)
EPEC merupakan E. coli penyebab diare yang tidak disebabkan oleh toksin
namun karena kemampuannya melakukan penempelan yang sangat kuat dan
menyebabkan terjadinya lesi/luka pada sel epitel usus (Adlberberth et al. 2000, Lu
& Walker 2001). Penempelan EPEC diinisiasi oleh bundle-forming pilus (BFP)
fimbrae. BFP merupakan plasmid berukuran 50-70 M Da, sehingga juga dikenal
dengan EAF plasmid (EPEC adherence factor). E. coli bergerak dengan flagella
peritrik. E. coli memiliki macam–macam fimbria atau pili sesuai struktur dan
speksitivitas antigen, antara lain membentuk filamen dan berupa protein. Fimbria
merupakan rangkaian hidrofobik dan merupakan organ spesifik yang juga
berfungsi sebagai adhesin/ reseptor penempelan (MacFarlane et al. 2000). Inisiasi
penempelan oleh BFP merupakan model interaksi antara bakteri dengan sel epitel
usus (Adlberberth et al. 2000). Setelah menempel, mikrovili usus dapat rusak
dan selanjutnya EPEC mensekresikan faktor virulen melalui reseptor Tir dan sisi
sekresi tipe III pada dinding sel EPEC ke dalam sel inang yaitu sel epitel usus.
EPEC lalu terikat ke sel epitel melalui outer membrane protein-intimin. Sinyal
transduksi terjadi dalam sel epitel usus, termasuk di antaranya aktivasi protein
kinase C (PKC), inositol triphosphat (IP3) dan pelepasan Ca2+. Beberapa protein
Perlindungan terhadap penyakit gastroenteritidis
Penyakit oleh bakteri enteropatogen
Flora normal
Bakteri patogen

27
sitoskeletal di antaranya aktin juga berperan dalam penempelan EPEC. Akhirnya
terjadi perubahan struktur sitoskeletal setelah penempelan Tir-intimin,
menghasilkan bentuk pedestal (Lu & Walker 2001). Hal inilah yang menyebabkan
EPEC mampu menempel lebih kuat daripada bakteri patogen lainnya (Gambar
2.8).
Gambar 2.8 Mekanisme penempelan EPEC di sel epitel usus (Lu & Walker 2001)
Identifikasi BAL
Identifikasi BAL sangat diperlukan untuk mengetahui genus, spesies
bahkan galurnya. Menurut FAO/WHO (2002), untuk dapat diketahui dan didekati
sifat fungsionalnya sebagai probiotik, maka BAL harus diketahui spesies bahkan
sampai galur untuk klaim sifat fungsional tertentu. Dalam perkembangannya,
metode identifikasi BAL dapat dibagi menjadi dua yaitu :
1. Metode klasik
Metode klasik meliputi metode untuk menguji morfologi dan sifat
biokimiawi BAL. Uji morfologi yang dimaksud adalah reaksi Gram, produksi
katalase dan morfologi sel. Uji biokimiawi meliputi produksi CO2 dari glukosa,
produksi NH3 dari arginin, produksi dekstran dari sukrosa, pertumbuhan pada
suhu 15 dan 45°C, pertumbuhan pada NaCl 6.5% dan reaksi pada litmus milk.
Semuanya akan dapat menentukan genus dari BAL. Untuk mengetahui sampai
tingkat spesies BAL, maka untuk metode klasik digunakan pengujian pola

28
fermentasi pada berbagai gula (49 jenis gula), yang umumnya digunakan adalah
API CHL dan API CHS (Counter et al. 2005; Reque et al. 2000; Vassu et al.
2001). Namun demikian, metode klasik ini mempunyai beberapa kelemahan
antara lain : (a) identifikasi hanya terbatas sampai tingkat spesies, (b) identifikasi
hanya tergantung pada pola hasil fermentasi beberapa gula sehingga terkadang
bias untuk beberapa spesies tertentu atau bahkan tidak dapat teridentifikasi.
Pengujian dengan sistem identifikasi API 50 CH yang dilakukan oleh Counter et
al. (2005) membuktikan bahwa dari 90 isolat BAL yang diisolasi dari sosis
fermentasi, terdapat 10% dari keseluruhan isolat tidak dapat diidentifikasi
(unidentified) dan ada kesalahan software dimana Lactobacillus sakei subsp. sakei
diidentifikasi sebagai Lactococcus lactis subsp. lactis.
Namun demikian, identifikasi dengan metode klasik tetap diperlukan
untuk mengetahui sifat fisiologis dan biokimiawi dari BAL. Pada seleksi BAL,
metode untuk mengetahui sampai tingkat genus umumnya menggunakan metode
klasik, penentuan dugaan awal sampai tingkat spesies juga digunakan metode
klasik. Namun, untuk mengkonfirmasi spesies BAL tersebut perlu dilakukan
identifikasi secara molekuler (Vamanu et al. 2006).
2. Metode modern/ molekuler
Metode molekuler dikembangkan untuk mengatasi kelemahan metode
identifikasi klasik. Di samping itu, perkembangan beberapa metode molekuler
ditujukan untuk mempertimbangkan kedekatan beberapa galur bakteri BAL.
Teknik PFGE (Pulse Field Gel Electrophoresis) dan RFLP (Restricted
Fragment Length Polymorphism)
Dasar teknik PFGE adalah penggunaan sistem getaran elektrik untuk
migrasi fragmen DNA yang besar melewati gel agarosa. Fragmen DNA diperoleh
dari digesting genom dengan enzim restriksi yang mempunyai recognition site 8
bp dan 6 bp. Fragmen DNA yang direstriksi di gel agarosa dimasukkan ke setiap
sumur agarosa dan dipisahkan dengan PFGE berdasarkan ukuran fragmennya
(Tannock 1999). Teknik RFLP mirip dengan teknik PFGE. Perbedaannya adalah
jika sebelum dielektroforesis restriksi DNA dilakukan pada gel untuk PFGE,
namun tidak untuk RFLP. Teknik RFLP dan PFGE ini dapat mendeteksi BAL
sampai karakteristik spesifik bakteri pada tingkat galur (Beasley 2004).

29
Ribotyping
Tahapan utama teknik ini adalah mengisolasi DNA dari isolat kultur,
dilakukan restriksi dan pemisahan ukuran di dalam gel agarosa. Selanjutnya gel
dihibidrisasi dengan menggunakan penanda rRNA yang dilabel, yang mengikat
fragmen berisi salinan dari operon rRNA. Untuk deteksi penanda, fragmen yang
terikat pada penanda selanjutnya divisualisasikan dan membentuk karakteristik
RFLP (Tannock 1999).
Analisis urutan basa (sequencing) gen 16S rRNA
Teknik ini merupakan teknik cepat yang mempunyai tahapan amplifikasi
gen 16S rRNA, menggunakan PCR dengan primer target yang berlaku secara
umum dan disepakati secara internasional untuk gen tersebut. Analisis urutan
basa filogenetik 16S rRNA merupakan metode molekuler yang akurat untuk
identifikasi mikroba (Walter et al. 2000; Anukam et al. 2005; Ennahar et al.
2003). Sekuen nukleotida-nukleotida pada gen 16S rRNA dapat diidentifikasi
lebih lanjut dengan menggunakan PCR-DGGE (Denaturating Gradient Gel
Electrophoresis) (Ennahar et al. 2003) dan TGGE (Temperature Gradient Gel
Electrophoresis) (Cocolin et al. 2000).
Tahapan yang dilakukan pada teknik analisis PCR-TGGE didahului
dengan ekstraksi DNA dan reaksi PCR dengan prinsip yang sama seperti
dijelaskan pada Anukam et al. (2005) kemudian dilanjutkan dengan analisis
TGGE. Tahapan prosedur analisis TGGE seperti yang dilakukan oleh Walter et
al. (2000) adalah sebagai berikut : elektroforesis dilakukan dengan menggunakan
gel poliakrilamid dan bufer TAE (tris asetat) pada tegangan 110 V selama 5 jam.
Gradien dari temperatur 65.5 – 68°C digunakan secara paralel pada saat
elektroforesis berjalan (running). Setelah elektroforesis selesai (komplet) maka
dilakukan pewarnaan pada TAE, selanjutnya gel divisualisasikan dengan
menggunakan sistem kamera polaroid.
Hibridisasi DNA
Hibridisasi DNA dilakukan untuk mengetahui kandungan G+C (mol%)
dari BAL. Metode yang digunakan adalah isolasi DNA lalu penentuan
kandungan G+C dilakukan dengan menggunakan thermal melting temperature

30
DNA menggunakan spektrofotometer (contohnya : Gilford Response
Spectrophotometer) (Krockel et al 2003).
DAFTAR PUSTAKA
Aattouri N, Bouras M, Tome D, Marcos A, Lemonnier D. 2002. Oral ingestion
of lactic acid bacteria by rats increases lymphocyte proliferation and interferon-γ production. Braz J Nutr 87: 367-373.
Aberle ED, Forrest JC, Hendrick HB, Judge MD, Merkel RA. 2000. Principles of Meat Science. San Fransisco:W.H. Freeman and Co.
Adlerberth I, Cerquetti M, Poilane I, Wold A, Collignon A. 2000. Mechanisms of colonization and colonization resistance of the digestive tract. Part 1: bacteria/host interactions. Microbial Ecol Health Disease. 2 : 223-239.
Agostoni CI et al. 2004. Probiotic bacteria in dietetic products for infants : a commentary by the ESPHGHAN Committee on Nutrition. J Pediatric Gastroenterol Nutr 28 : 365-374.
Albano H et al. 2007. Characterization of two bacteriocins produced by Pediococcus acidilactici isolated from ‘Alheira’ a fermented sausage traditionally produced in Portugal. Int J of Food Microbiol 116 : 239-247.
Antara NS, Sujaya IN, Yokota A, Asano K, Tomita F. 2004. Effects of indigenous starter cultures on the microbial and physicochemical uharacteristics of Urutan, a Balinese fermented sausage. J Biosci and Bioeng 98 : 92-98.
Anukam KC, Osazuwa EO, Ahonkai I, Rheid G. 2005. 16S rRNA gene sequence and philogenetic tree of Lactobacillus spesies from the vagina of healthy Nigerian women. Afr J of Biotechnol 4 : 1222-1227.
Arief II, Maheswari RRA, Suryati T. 2007. Karakteristik dan Nilai Gizi Protein Daging Sapi Dark Firm Dry (DFD) yang Difermentasi oleh Lactobacillus plantarum yang Diisolasi dari Daging Sapi. Laporan Penelitian Hibah Bersaing XIII. LPPM-IPB.
Augenlicht LH et al. 1999. Short chain fatty acid metabolism, apoptosis and Apc-initiated tumorigenesis in the mouse gastrointestinal mucosa. Cancer Research 59:6005-6009.
Axelsson, L. 1993. Lactid Acid Bacteria : classification and physiology. Di dalam: Lactid Acid Bacteria: Microbiology and Functional Aspects. 2nd Edition, Revised and Expanded. Salminen, S., and von Wright, A. (Editors.). New York : Marcell Dekker Inc.

31
Aymeric T, Martin B, Garriga M, Hugas M. 2003. Microbial quality and direct PCR identification of lactic acid bacteria and nonpatogenic Staphylococci from artisanal low-acid sausages. Appl Environ Microbiol 69 : 4583-4594.
Badan POM. 2005. Keputusan Kepala Badan Pengawas Obat dan Makanan Republik Indonesia Nomor : 005.05.1.0421 tentang pembentukan tim mitra bestari pangan fungsional.
Bhatia A, Rani U. 2008. Immunomodulatory effect of S. thermophilus : an experimental study. J Calicut Med 6 :2-10.
BC Dairy Foundation. 1997. The probiotic effects of lactic acid bacteria, an interpretive review of recent nutrition research.
Beasley S. 2004. Isolation, identification and exploitation of lactic acid bacteria from human and animal microbiota. Academic Dissertation in Microbiology. Faculty of Agricultue and Forestry Science. University of Helsinki.
Begley M, Gahan CGM, Hill C. 2005. The interaction between bacteria and bile. FEMS Microbiol Rev 29 : 625-651.
Begley M, Hill C, Grahan CGM. 2006. Bile salt hydrolase activity in probiotics. Appl Environ Microbiol 72: 1729-1738.
Blum S et al. 1999. Adhesion studies for probiotics : need for validation and refinement. Trends in Food Sci Technol 10 : 405-410.
Branen AL, Davidson PM. 1993. Antimicrobial in Food. New York : Marcel Dekker.
Bourlioux P, Koletzko B, Guarner F, Braesco V. 2003. The intestine and its microflora are partners for the protection of the host : report on the Danone symposium ‘ The Intelligent Intestine’, held in Paris, June 14, 2002. Am J Clin Nutr 78: 675-683.
Brady LJ, Gallaher DD, Busta FF. 2000. The role of probiotic cultures in the prevention of colon cancer. Symposium : Probiotic bacteria : Implications for Human Health. J Nutr 410S-414S.
Bromberg R, Moreno I, Zagagini CL, Delboni RR, Oliveira J. 2004. Isolation of bacteriocin-producing lactic acid bacteria from meat and meat products and its spectrum of inhibitory activity. Braz J Microbiol 35:137-144.
Bron PA et al. 2004. Genetic characterization of the bile salt response in Lactobacillus plantarum and analysis of responsive promoters in vitro and in situ in the gastrointestinal tract. J Bacteriol 186 : 7829-7835.
Burns AJ, Rowland IR. 2000. Anti-carcinogenicity of probiotics and prebiotics. Curr Issues Intest Microbiol 1: 13-24.

32
Cai Y, Suyanandana P, Saman P, Benno Y. 1999. Classification and characterization of lactic acid bacteria isolated from the intestines of common carp and freshwater prawns. J Gen Appl Microbiol 45 : 177-184.
Cocolin L, Manzano M, Antoni C, Corni G. 2000. Development of a rapid method for the identification of Lactobacillus spp. Isolated from naturally fermented Italian sausages using a Polymerase Chain Reaction-Temperature Gradient Gel Electrophoresis (PCR-TGGE). L Appl Microbiol 30 : 126-129.
Cotter PD, Hill C. 2003. Surviving the acid test : responses of Gram-positive bacteria to low pH. Microbiol Molecul Biol Rev 67: 429-453.
Counter M et al. 2005. Characterization of lactic acid bacteria isolated from Italian dry fermented sausage. Ann Fac Medic Vet 27 : 167-174.
Danone research team. 2002. Danone Nutritopics: The three lines of defense in human intestine; Effect of probiotics on body’s natural defenses. 10th International Congress of Bacteriology and Applied Microbiology “The Word of microbes’. Paris.
El-Naggar MYM. 2004. Comparative study of probiotic cultures to control the growth of Escherichia coli O157:H7 and Salmonella typhimurium. Biotechnol 3: 173-180.
Emanuel V, Adrian V, Ovidiu P, Gheorghe C. 2005. Isolation of Lactobacillus plantarum strain used for obtaining a product for the preservation of fodders. Afr J Biotechnol 4 : 403-408.
Ennahar S, Cai Y, Fujita Y. 2003. Phylogenetic Diversity of lactic Acid bacteria associated with paddy rice silage as determined by 16S Ribosomal DNA Analysis. Appl Environ Microbiol 69 : 444-451.
Erkilla S. 2001. Bioprotective and probiotic meat starter cultures for the fermentation of dry sausages. [Academic Dissertation]. Department of Food Technology. University of Helsinki.
Erkilla S, Petaja E. 2000. Screening of commercial meat starter cultures at low pH and in the presence of bile salts for potential probiotic use. Meat Sci 55 : 297-300.
FAO/ WHO. 2002. Guidelines for the evaluation of probiotics in food. Report of Joint FAO/WHO Working Group on drafting Guidelines for the evaluation of probiotics in food. London Ontario, Canada.
Gackowska L et al. 2006. Combined effect of different lactic acid bacteria galur on the mode of cytokines pattern expression in human peripheral blood mononuclear cells. J Physiol Pharmacol 57 : 13-21.

33
Hamasaki Y, Ayaki M, Fuchu H, Sugiyama M, Morita H. 2003. Behaviour of pschotrophic lactic acid bacteria isolated from spoiling cooked meat products. Appl Environ Microbiol 69 : 3668-3671.
Helander IM, von Wright A, Mattila-Sandholm TM. 1997. Potential of lactic acid bacteria and novel antimicrobials against Gram-negative bacteria (review). Trends in Food Sci Technol 8: 146-150.
Herich R, Levkut M. 2002. Lactic acid bacteria, probiotics and immune system. Vet Med Czech 47 :169-180.
Hopzapfel WH. 1998. The Gram-poisitive bacteria associated with meat and meat products. Dalam : The Microbiology of Meat and Poultry. Davies A and R. Board (editors). London : Blackie Academic & Proffesional.
Ishibashi N, Yamakazi S. 2001. Probiotics and safety. Am J Clin Nutr 73 (suppl) : 4658-4708.
Itoi S et al. 2008. Isolation of halotolerant Lactococcus lactis subsp. lactis from intestinal tract of coastal fish. Int J Food Microbiol 121 : 116-121.
Jenie BSL, Rini SE. 1995. Aktivitas antimikroba dari beberapa spesies Lactobacillus terhadap mikroba patogen dan perusak makanan. Bul Teknologi dan Industri Pangan. 6: 46 – 51.
Jenie BSL. 1996. Peranan BAL sebagai pengawet hayati makanan. J Ilmu dan Tek Pangan 1: 60-73.
Karaoglu SA, Aydin F, Killic SS, Killic AO. 2003. Antimicrobial activity and characteristics of bacteriocins produced by vaginal lactobacilli. Turk J Med Sci 32 : 7-13.
Kimoto-Nira H et al. 2007. Lactococcus sp. as potential probiotic lactic acid bacteria. JARQ 41: 181-189.
Kimura M, Danno K, Yasui H. 2006. Immunomodulatory function and probiotic properties of lactic acid bacteria isolated from Mongolian fermented milk. Biosci Microflora 25 : 147-155.
Kos B et al. 2003. Adhesion and agggregration ability of probiotic galur Lactobacillus acidophilus M92. J Appl Microbiol 94: 981-987.
Krockel L, Schilinger U, Franz CAMP, Bantleon C, Ludwig W. 2003. Lactobacillus versmoldensis sp nov., isolated from raw fermented sausage. Int J Sys Evol Microbiol 53 : 513-517.
Kusumawati N. 2002. Seleksi BAL Indigenus sebagai Galur Probiotik dengan Kemampuan Mempertahankan Keseimbangan Mikroflora Feses dan Mereduksi Kolesterol Serum Darah Tikus. [Thesis]. Program Pascasarjana Institut pertanian Bogor.

34
Lee YK. 2009. Probiotic Microorganisms. Di dalam : Handbook of Probiotic and Prebiotics. 2nd edition. Yuan Kun Lee and Seppo Salminen (editor). John wiley & Sons, Inc.
Leisner JJ et al. 1999. Identification of lactic acid bacteria from Chili Bo, a Malaysian food ingredient. Appl Environ Microbiol 65 : 599 – 605.
Lin WH, Hwang CF, Chen LW, Tsen HY. 2006. Viable counts, characteristic evaluation for commercial lactic acid bacteria products. Food Microbiol 23: 78-81.
Lu L, Walker WA. 2001. Pathologic and physiologic interactions of bacteria with the gastrointestinal epithelium. Am J Clin Nutr 73 (suppl) ; 1124S-1130S.
Mac. Farlane S, Hopkins MJ, Mc.Farlane GT. 2000. Bacterial growth and metabolism on surfaces in the large intestine. Microbial Ecology in health and Disease (Suppl) 2 : 64-72.
Makarova KS, Koonin EV. 2007. Evolutionary Genomic of Lactic Acid Bacteria. J Bact 189 : 1199-1208.
Marteau P, Minekus M, Havenar R, Huis In’t Veld JHJ. 1997. Survival of lactic acid bacteria in a dynamic model of the stomach and small intestine : validation and the effects of bile. J Dairy Sci 80: 1031-1037.
Medellin-Pena MJ, Griffiths MW. 2009. Effects of molecules secreted by Lactobacillus acidophilus galur La-5 on Escherichia coli O157:H7 Colonization. Appl Environ Microbiol 75 : 1165-1172
Meghrous J, Lacroix C, Bouksaim M, La-Pointe G, Simard RE. 1997. Genetic and biochemical characterization of nisin Z produced by Lactococcus lactis ssp. lactis biovar. diacetylactis UL 719. J Appl Microbiol 83 : 133-138.
Mishra V, Prasad DN. 2005. Application of in vitro methods for selection of Lactobacillus casei galurs as potential probiotics. Int J Food Microbiol 103 : 109-115.
Molin G. 2003. The role of Lactobacillus plantarum in foods and in human health. Dalam: Handbook of Fermented Functional Foods. E.R. Farnworth (Editor). Boca Raton: CRC Press.
Moll GN, Konnings WN, Driessen AJM. 1999. Bacteriocins: mechanism of membrane insertion and pore formation. Antonie van Leeuwenhoek 76: 185-198.
Morelli L. 2007. In vitro assessment of probiotic bacteria : from survival to functionally. Int Dairy J 17: 1278-1283.

35
Moser SA, Savage DC. 2001. Bile salt hydrolase activity and resistance to toxicity of conjugated bile salts are unrelated properties in Lactobacilli. Appl Environ Microbiol 67: 3476-3480.
Moulay M et al. 2006. Cultivable lactic acid bacteria isolated from Algerian raw goat’s milk and their proteolytic activity. World J Dairy Food Sci 1: 12-18
Naidu AS, Clemens RA. 2000. Probiotics. Dalam Natural Food Antimicrobial Systems. A.S. Naidu (editor). Florida : CRC Press.
Nair PS, Surendran PK. 2005. Biochemical Characterization of lactic acid bacteria isolated from fish and prawn. J Cult Collect 4 : 48-52.
Naruszewicz ML, Johansson D. Downar Z, Bukowska H. 2002. Effect of Lactobacillus plantarum 299v on cardiovascular disease risk factors in smokers. Am J Clin Nutr 76 : 1249-1255.
Nitisinprasert S, Pungsungworn N, Wanchaitanawong P, Loiseau G, Montet D. 2006. In vitro adhesion assay of lactic acid bacteria, Escherichia coli and Salmonella sp. by microbiological and PCR methods. Songklanakarin J Sci Technol 28 (suppl.1) : 99-106.
Nowroozi J, Mirzaii M, Norouzi M. 2004. Studi of Lactobacillus as Probiotic Bacteria. Iranian J Publ Health 33 : 1-7.
Nuraida L, Prisilia D, Nurjanah S. 2010. Use of Lactobacillus rhamnosus isolated from breast milk as starter culture of yoghurt [abstrak]. Di dalam : 2nd International Symposium on Probiotic and Prebiotic, as Functional food for Human Health Promotion; Jakarta 4-5 Agustus 2010. hlm 55.
Ogunbanwo ST, Sanni AI, Onilude AA. 2003. Characterization of bacteriocin produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1. Afr J Biotechnol 2:219-227
Ouwehand AC, Tuomola EM, Tolkko S, Salminen S. 2001. Assessment of adhesion properties of novel probiotics galurs to human intestinal mucus. Int J of Food Microbiol 64 : 119-126
Oyetayo VO. 2004. Performance of rats orogastrically dosed with faecal galurs of Lactobacillus acidophilus and challenged with Escherichia coli. Afr J Biotechnol 3: 409-411.
Pal V, Jamuna M, Jeevaratnam K. 2005. Isolation and Characterization of Bacteriocin Producing Lactic Acid Bacteria from a South Indian Special Dosa (APPAM) Batter. J Culture Collection 4: 53-60
Perdigon, Fuller GR, Raya R. 2001. Lactic acid bacteria and their effect on immune system. Curr Issues Intest Microbiol. 2 : 27-42.

36
Plengvidhya V, Breidt FJ, Lu Z, Fleming HP. 2007. DNA fingerprinting of lactic acid bacteria in sauerkraut fermentations. Appl Environ Microbiol 73 : 7697-7702.
Rantsiou et al. 2005. Culture-dependent and independent methods to investigate the microbial ecology of Italian fermented sausages. Appl Environ Microbiol 71 : 1977-1986.
Ratanapibulsawat, C., P. Kroujkaew, O. Sadahiro, and S. Nitisinprasert. 2005. Screening and characterisation of lactic acid bacteria producing antimicrobial substance againts Staphylococcus aureus. Kasetsart Journal (Natural Science) 39 : 284-293.
Reid G. 1999. The scientific basis for probiotic galurs of Lactobacillus. Appl Environ Microbiol 65 : 3763-3766.
Reque EF, Pandey A, Franco SG, Soccol CR. 2000. Isolation, identification and physiological study of Lactobacillus fermentum LPB for use as probiotic in chickens. Braz J Microbiol 31 : 303-307.
Roberfroid MB. 2000. Prebiotics and probiotics: are they functional foods?. Am Clin Nutr 71(suppl) : 1682S-1687S.
Robredo B, Torres C. 2000. Bacteriocin Production by Lactobacillus salivarius of Animal Origin. J Clin Microbiol 38 : 3908-3909.
Salminen S, Wright AV. 2004. Lactic Acid Bacteria. Microbiology and Functional Aspects. 2nd Edition, Revised and Expanded. New York : Marcell Dekker, Inc.,
Savadogo A, Outtara CAT, Bassole IHN, Traore AS. 2006. Bacteriocins and lactic acid bacteria – a minireview. Afr J Biotechnol 5: 678-683.
Segawa S et al. 2008. Effect of oral administratition of heat-killed Lactobacillus brevis SBC8803 on total and ovalbumin-spesific immunoglobulin E production through the improvement of Th1/Th2 balance. Int J Food Microbiol 121 : 1-10.
Solis B et al. 2002. The effect of fermented milk on interferon production in malnourished children and in anorexia nervosa patient undergoing nutritional care. Eur J Clin Nutr 56 (suppl 4) : 527-533.
Suarsana IN, Utama IH, Suartini NGAA. 2001. Aktivitas invitro senyawa antimikroba dari Streptococcus lactis. J Vet 2 : 25-31.
Sunny-Roberts EO, Knoor D. 2008. Evaluation of the response of Lactobacillus rhamnosus VTT E-97800 to sucrose-induced osmotic stress. Food Microbiol 25 : 183-189.

37
Surono IS. 2010. Potency of local wisdom : probiotics and prebiotics today and tomorrow. 2nd International Symposium on Probiotic and Prebiotic, as Functional Foods for Human Health Promotion : health benefits, local knowledge, technical and regulatory issues. Jakarta, August 4-5 2010.
Surono IS. 2004. Probiotik, Susu Fermentasi dan Kesehatan. YAPPMI. Jakarta.
Tamang B et al. 2008. Phenotytpic and genotypic identification of lactic acid bacteria isolated from ethnic bamboo tender shoots of North East India. Int J Food Microbiol 121 : 35-40.
Talwalkar A, Kailasapathy K. 2004. The role of oxygen in the viability of probiotic bacteria wirh reference to L.acidophilus and Bifidobacterium spp. Curr Issues Intest Microbiol 5 : 1-8.
Tannock GW. 1999. Probiotic :A Critical Review. Horizon Press. Norfolk.
Toksoy A, Beyatli Y, Aslim B. 1999. Study on metabolic and antimicrobial activities of some L. plantarum galurs isolated from sausages. Tr J Vet An Sci 533-540.
Tuomola E, Crittenden R, Playne M, Isolauri I, Salminen S. 2001. Quality assurance criteria for probiotic bacteria. Am J Clin Nutr 73 (suppl): 393S-398S.
Vamanu A, Vamanu E, Drugulescu M, Popa O, Campeanu G. 2006. Identification of lactic bacterium galur used for obtaining a pollen-based probiotic product. Turk J Biol 30 : 75-80.
Vasiljevic T, Shah NP. 2008. Probiotic-from Metchnikoff to bioactives. Int Dairy J 18 : 714-728.
Vassu T et al. 2001. Biochemical and genetic characterization of Lactobacillus plantarum cmgb-1 galur used as probiotic. Roum Biotechnol Lett 7 : 585-598.
Walter J et al. 2000. Detection and identification of gastrointestinal species by using Denaturating Gradient gel Electrophoresis (DGGE) and spesific-spesific PCR primer. Appl Environ Microbiol 66 : 297:303.
Wollowski I et al. 1999. Bacteria used for the production of yogurt inactive carcinogens and prevent DNA Damage in the Colon of Rats. J Nutr 77-82.
Wollowski I, Rechkemmer G, Pool-Zobel BL. 2001. Protective role of probiotics and prebiotics in colon cancer. Am J Clin Nutr 73 (suppl) : 451S-455S.
Zoumpopoulou G et al. 2008. Lactobacillus fermentum ACA-DC 179 displays probiotic potential in vitro and protects againts trinitrobenzene sulfonic acid (TNBS)-induced colitis and Salmonella infection in murine models. Int J Food Microbiol 121 : 18-19.










